










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.17 no.3 Bogotá Sept./Dec. 2012
MORFOLOGÍA Y MORFOMETRÍA DE DOS ESPECIES DE BANDERITAS (ORCHIDACEAE: MASDEVALLIA) EN COLOMBIA
Morphology and Morphometry of Two Banderitas Species (Orchidaceae: Masdevallia) in Colombia
MÓNICA ADRIANA CUERVO MARTÍNEZ1, M.Sc.; MARÍA ARGENIS BONILLA GÓMEZ2, Ph. D.; RODRIGO BUSTOS SINGER3, Ph. D.
1Grupo Biología de Organismos Tropicales. Universidad Nacional de Colombia. Bogotá, Colombia. macuervom@unal.edu.co
2Profesora Asociada, Departamento de Biología, Universidad Nacional de Colombia, Grupo Biología de Organismos Tropicales. Bogotá, Colombia. mabonillag@unal.edu.co
3Profesor Universidade Federal de Rio Grande do Sul. Rio Grande do Sul, Brasil. rbsinger1@yahoo.com.Correspondencia: Mónica Adriana Cuervo Martínez, diagonal 82C # 73A-59, interior 22. macuervom@unal.edu.co
Presentado el 8 de febrero de 2012, aceptado el 13 de agosto de 2012, correcciones el 15 de agosto de 2012.
RESUMEN
Masdevallia coccinea y Masdevallia ignea (Orchidaceae) conocidas como "banderitas", son orquídeas ornamentales muy apreciadas por cultivadores aficionados y coleccionistas. En Colombia, la presión de colecta sobre estas especies ha sido enorme y pocas poblaciones naturales sobreviven en los departamentos de Boyacá (Arcabuco, Páramo de Berlín, Duitama) y Santander (entre Málaga y Bucaramanga), estas poblaciones son reducidas y de difícil acceso; razón por la cual se encuentran en el apéndice II de CITES. Poco se sabe sobre su biología reproductiva, sistema reproductivo y polinización y parte de lo que consta en la literatura es incompleto. En este marco el objetivo general del trabajo fue estudiar la morfología y morfometría floral de M. coccinea y M. ignea en condiciones de semicultivo al aire libre en la finca Villa Rosa ubicada en el municipio de Guasca, Cundinamarca, para lo cual se realizó fotografía digital, pruebas histoquímicas, morfometría y Microscopía Electrónica de Barrido. Los resultados principales de la morfometría mostraron que los sépalos dorsal y lateral fueron más largos en M. coccinea (X= 53,0 mm σ = 7,4 mm y X = 44,4 mm y σ = 8,3 mm) en comparación a M. ignea (X = 34 mm σ = 7,7 mm y X = 31,5 mm y σ = 6,1 mm), pero las coloraciones de las partes florales de esta última fueron más intensas y su labelo fue más largo (X = 7,1 mm y σ = 0,6 mm). En las dos especies el labelo estuvo articulado a la columna y no se encontró presencia de glándulas con estructuras secretoras como nectarios ni osmóforos.
Palabras clave: labelo, Masdevallia coccinea, Masdevallia ignea, morfología, morfometría floral.
ABSTRACT
Masdevallia coccinea and the Masdevallia ignea "banderitas" are ornamental orchids which are very prized by amateur farmers and collectors. In Colombia, the harvest pressure on these species has been enormous and few natural populations survive in the departments of Boyacá (Arcabuco of Berlin Páramo, Duitama) and Santander (between Málaga and Bucaramanga), in which these populations are reduced and of difficult access. For this reason these species are in the II appendix of CITES. However, little is known on their reproductive biology, floral biology and pollination and the literature about this is incomplete. Under this framework, the goal of the project was to study the morphology and morphometry of M. coccinea and M. ignea (Pleurothallidinae) under semicultivation conditions in the Villa Rosa farm located in the Municipality of Guasca, Cundinamarca (Colombia). The floral morphology was analyzed by digital photography, morphometry and Scanning Electron Microscope. The main results were differences in color and length of dorsal and lateral sepals between M. coccinea (X = 53.0 mm σ = 7.4 mm and X = 44.4 mm and σ = 8.3 mm) and M. ignea (X = 34 mm σ = 7.7 mm and X = 31.5 mm and σ = 6.1 mm). These parts were longest in M. coccinea in contrast to M. ignea. However the lip was longest in M. ignea (X = 7.1 mm y σ = 0.6 mm). On the other hand both species had lip articulated to the column but without rewards as nectar and osmophores.
Keywords: Lip, Masdevallia coccinea, Masdevallia ignea, morphology, floral morphometry.
INTRODUCCIÓN
El género Masdevallia Ruiz & Pav. pertenece a la subtribu Pleurothallidinae Lindl ex G. Don y se distribuye desde Brasil hasta México. Comprende cerca de 330 especies, siendo Colombia uno de los centros de mayor diversidad (Fernández, 1971; Koniger y Koniger, 1995; Luer, 2000; Ortiz, 1995; Ortiz, 2000, Abele, 2007). En Colombia se reporta en las cordilleras Oriental y Central y en el Nudo de Los Pastos en el sur del país (Calderón y Farfán, 2003).
La mayoría de las especies de este género son epifitas, que crecen en bosques montanos lluviosos de altitudes entre los 1500 msnm y los 2500 msnm, en ambientes sujetos a lluvia o neblina y una humedad relativa del aire entre 80 y 90 %. Algunas especies son también terrestres o litofíticas (Abele, 2007).
Las especies del género se caracterizan por ser plantas perennes, con hojas con gran variación en la forma. Con flores solitarias o en inflorescencias en racimo; flores zigomórficas, con una o varias brácteas florales y ovario trilobado. Los sépalos conspicuos y variadamente connados. Los pétalos callosos, pequeños e inconspícuos que usualmente forman una carina. El labelo, no más largo que los pétalos laterales y más o menos ligulado. La columna semiterete con la antera ventral y dos polinios (Abele, 2007). Las banderitas se caracterizan por presentar dos polinios (400-1500 µm), lateralmente aplanados y en forma de triángulo de elongado a comprimido, caudículas comúnmente elaboradas, superficie comúnmente cubierta con elastovicina, escultura ciliada a puntada, o granulada a gemada (Stenzel, 2000).
Existen pocos datos publicados sobre la polinización de especies de Masdevallia, van der Pijl y Dodson (1966) documentaron algunos casos de polinización por moscas pequeñas similares a Drosophila en M. erythochaete Rchb.f. y moscas no identificadas en M. fractiflexa F. Lehm. & Kraenzl. Asimismo, estos autores sugirieron que las especies del género con perianto rojizo a rosado (como es el caso de especies estudiadas aquí) podrían ser polinizadas por colibríes (Trochilidae). Observaciones inéditas (Singer) indican que especies de Masdevallia con flores de aroma afrutado (como M. floribunda Lindl.), son polinizadas por moscas Drosophilidae, igual que M. infracta Lindl. en Brasil. Además, existen otros trabajos sobre aspectos reproductivos de M. ignea en el Jardín Botánico de Bogotá (Chavarro, 2005; Chavarro et al., 2006). La polinización por moscas parece ser la regla en las orquídeas de la subtribu Pleurothallidinae (Dressler,1993). Hasta ahora, no se han detectado néctar u otras secreciones en las flores frescas de M. ignea (Chavarro et al., 2006) y M. coccinea. Masdevallia coccinease distribuye en Colombia en los departamentos de Cundinamarca, Santander y Norte de Santander, en un rango altitudinal entre 2600 msnm y 3500 msnm (Calderón y Farfán, 2003). Es una orquídea semiterrestre, con reproducción sexual y asexual. M. ignea se distribuye en Colombia en Norte de Santander entre los 2950 msnm y 3130 msnm, se encuentra en la cordillera Oriental en cercanías de la Sierra Nevada del Cocuy (Calderón y Farfán, 2003; Chavarro, 2005; Chavarro et al., 2006), es una orquídea semiterrestre.
En Colombia, la presión de colecta sobre estas especies ha sido enorme y pocas poblaciones naturales sobreviven en los departamentos de Boyacá (Arcabuco, Páramo de Berlín, Duitama) y Santander (entre Málaga y Bucaramanga); estas poblaciones son reducidas y de difícil acceso; razón por la cual actualmente se encuentran en el apéndice II de Cites (UNEP WCMC, 2003). El nombre común de estas especies es banderitas, representan un buen modelo ecológico para comprender la biología reproductiva en este género y son carismáticas por su importancia como ornamentales. Poco se sabe sobre su biología reproductiva y polinización (van der Pijl y Dodson, 1966). Por lo tanto, es fundamental conocer aspectos de la biología y la morfología floral funcional, la articulación del labelo a la columna, la disposición del labelo respecto a la columna, la presencia de recompensas como glándulas con estructuras secretoras como nectarios y osmóforos, entre otras. Todas estas características están relacionados con el despliegue floral, el éxito reproductivo, la eficiencia de la polinización y el acople morfológico de la flor y el polinizador. En conjunto, son aspectos fundamentales para futuros planes de conservación y manejo de relictos naturales de estas especies.
En este contexto se estudió la morfología floral de las banderitas (Masdevallia coccinea Linden ex Lindl. y Masdevallia ignea Rchb. f.) las cuales son orquídeas ornamentales, muy apreciadas por cultivadores aficionados y coleccionistas (Leathers, 2007) y cuyas poblaciones naturales son escasas y se ven reducidas por la destrucción y transformación de los hábitats.
MÉTODOS
Stitio de estudio
La investigación se realizó entre febrero y agosto de 2009 en un semicultivo en la finca Villa Rosa ubicada en el municipio de Guasca, Cundinamarca (04°52'00"N y 73°52'59"W) a 2600 msnm. El sitio se caracteriza por una temperatura media de 13,2 °C, una precipitación media mensual de 65,7 mm, con precipitaciones máximas en el mes de mayo (98,3 mm) y una humedad relativa de 85,3 % (Instituto de Hidrología Meteorología y Estudios Ambientales de Colombia, 2011). En el semicultivo los individuos de Masdevallia se encuentran en remanentes de bosque que ha conservado el propietario, sembradas en el suelo y mezcladas con otras orquídeas.
MORFOLOGÍA FLORAL
La morfología floral se caracterizó en diez flores frescas de cada una de las especies. Las partes florales (sépalos, pétalos, labelo, columna, capuchón de la antera y polinios) se observaron en un estereoscopio Nikon SMZ800, y se tomaron fotografías de microscopía digital y MEB. Las fotografías digitales se realizaron con una cámara digital Nikon Coolpix 950 en el Laboratorio de Investigaciones en Biología Vegetal del Departamento de Biología de la Universidad Nacional de Colombia.
MORFOMETRÍA FLORAL
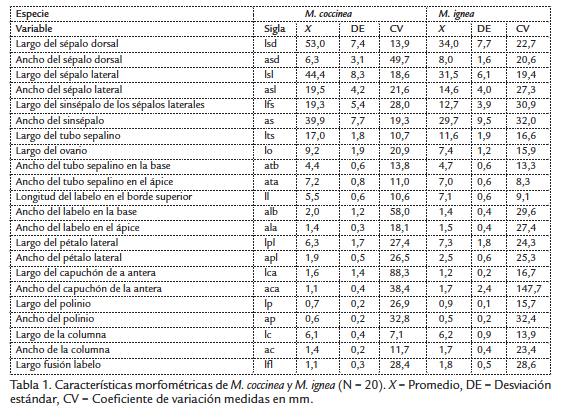
Para comparar las flores de M. coccinea y M. ignea se midieron 21 variables morfológicas (N = 10-20 dependiendo de la disponibilidad y estado de las flores) en flores frescas en antesis con un calibrador análogo (Spi 2000, precisión 0,1 mm). Las variables morfológicas medidas estuvieron relacionadas con: i) el despliegue floral: largo del sépalo dorsal (lsd), ancho del sépalo dorsal (asd), largo del sépalo lateral (lsl), ancho del sépalo lateral (asl), largo del sinsépalo de los sépalos laterales (lfs), ancho del sinsépalo (as); ii) el acceso de los visitantes y/o polinizadores a la flor: largo del tubo sepalino (lts), largo del ovario (lo), ancho del tubo sepalino en la base (atb), ancho del tubo sepalino en el ápice (ata). Y finalmente, iii) el mecanismo de polinización y el acople morfológico entre la flor y el polinizador: longitud del labelo (ll), ancho del labelo en el ápice (ala), ancho del labelo en la base (alb), largo del pétalo lateral (lpl), ancho del pétalo lateral (apl), largo del capuchón de la antera (lca), ancho del capuchón de la antera (aca), largo del polinio (lp), ancho del polinio (ap), largo de la columna (lc), ancho de la columna (ac), largo de la fusión del labelo a la columna (lfl). Se siguió la metodología propuesta por Knyasev et al., 2000.
La morfometría floral se analizó con estadística descriptiva (promedio, desviación estándar, valores máximo y mínimo), coeficiente de variación, correlación entre las variables (Spearman). Se verificó la normalidad de los datos y luego se realizó un análisis no-paramétrico de U de Mann Whitney para probar si existían diferencias significativas de los valores promedio de las variables para las dos especies.
HISTOQUÍMICA
Para establecer la existencia de recompensas florales y la presencia de glándulas secretoras (nectarios, osmóforos y glándulas secretoras), se sumergieron cinco flores de diferentes plantas por cada especie de Masdevallia por 20 minutos en una solución 1/1000 de rojo neutro en agua destilada y se observó al estereoscopio la presencia de puntos rojos que denotaran la presencia de nectarios en la superficie de sépalos, pétalos y puntos de inserción de los mismos al ovario. Adicionalmente, se sumergieron dos flores de cada especie en agua destilada como control (Davies et al., 2003; Dafni et al., 2005).
MICROSCOPÍA ELECTRÓNICA DE BARRIDO (MEB)
Para evidenciar la presencia de tricomas y glándulas en las superficies de sépalos, pétalos, columna, labelo y polinios que no fueron visibles con la fotografía digital, se realizaron fotografías de MEB. Estas estructuras (tricomas y glándulas) son fundamentales en el reconocimiento táctil de los insectos. Las muestras de sépalo lateral, sépalo dorsal, columna, pétalo, labelo y polinios fueron separadas de la flor, fijadas en glutaraldehído por 24 horas, luego deshidratadas en una serie de alcoholes de graduación creciente (50 %, 70 %, 100 %), dejando actuar el alcohol por períodos de 15 min siguiendo el protocolo de Davies et al. (2003, 2008). Posteriormente, se utilizó la técnica de secado en punto crítico (SAMDRI®-795), y los preparados se montaron sobre portaláminas, se realizó la metalización con oro (Metalizador Dentom Vacuum Desk IV) y finalmente las fotografías respectivas (Microscopio Electrónico de Barrido JEOL JSM- 6490LV) con una resolución de 3,0 nm. Las fotografías se realizaron en el Laboratorio de Microscopia Electrónica de Barrido de la Universidad de los Andes. Posteriormente, las fotografías se observaron detalladamente para detectar la presencia de tricomas y glándulas (Davies et al., 2003).
RESULTADOS
MORFOLOGÍA FLORAL Masdevallia coccinea
Las ramas de M. coccinea presentaron una flor solitaria de color rosado oscuro; sépalos dorsal y lateral conspicuos y pubescentes de color rosado oscuro; parte interna del sépalo dorsal y base de los sépalos laterales de color amarillo; sépalos connados formando un tubo sepalino arqueado más largo que ancho; sépalo dorsal triangular y reflexo; sépalos laterales triangulares; pétalos blancos ovados, con una vena media prominente, cristales y tricomas abundantes; labelo blanco, inconspícuo, oblongo, ápice amarillo, superficie con tres venas longitudinales blancas, articulado a la columna; columna blanca, paralela al labelo; antera ventral, blanca, granulosa y con cristales; estigma ventral, dos polinios amarillos, lateralmente aplanados (Fig. 1).
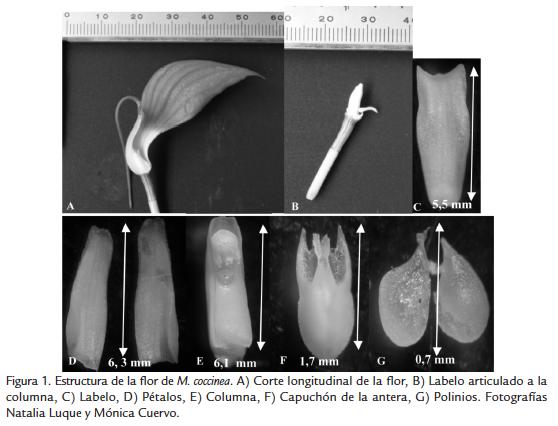
Las partes de las flores con medidas más variables se encontraron en los polinios, el labelo y el sépalo dorsal. Las variables que presentaron mayor coeficiente de variación en M. coccinea fueron: el largo del capuchón de la antera (88), el ancho del labelo en el ápice (58), el ancho del sépalo dorsal (50), el ancho del capuchón de la antera (38) y el ancho del polinio (33) (Tabla 1).
Las variables más altamente correlacionadas fueron: el largo del sépalo dorsal (lsd) y el ancho del sépalo lateral (asl) (r = 0,608 p < 0,0001), el largo del sépalo dorsal (lsd) y el ancho del sinsepalo (as) (R 0,535 p < 0,0001), el ancho del sépalo lateral (asl) y el ancho del sinsepalo (as) (R 0,955 p < 0,0001), el ancho del tubo sepalino en la base (atb) y el ancho del labelo en el ápice (ala) (R 0,623 p < 0,0001).
En M. coccinea las fotografías de MEB evidenciaron la presencia de tricomas en las superficies del sépalo dorsal y de los pétalos (Fig. 2).

MORFOLOGÍA FLORAL Masdevallia ignea
Las ramas de M. ignea presentaron una flor solitaria de color rojo oscuro; sépalos dorsal y lateral conspicuos y pubescentes de color rojo oscuro; parte interna del sépalo dorsal y base de los sépalos laterales de color amarillo; sépalos connados formando un tubo sepalino más largo que ancho; sépalo dorsal subtriangular, retrorso; sépalos laterales ovado triangulares, connados en la base; pétalos blancos, ovados, ápice redondeado, con una vena media prominente morada, cristales y tricomas abundantes; labelo blanco, ovado con una mancha de color morado en el centro, articulado a la columna con tres venas longitudinales blancas; columna blanca, con bordes morados, paralela al labelo; antera ventral, blanca, granulosa y con cristales; estigma ventral, dos polinios amarillo claro, lateralmente aplanados (Fig. 3).
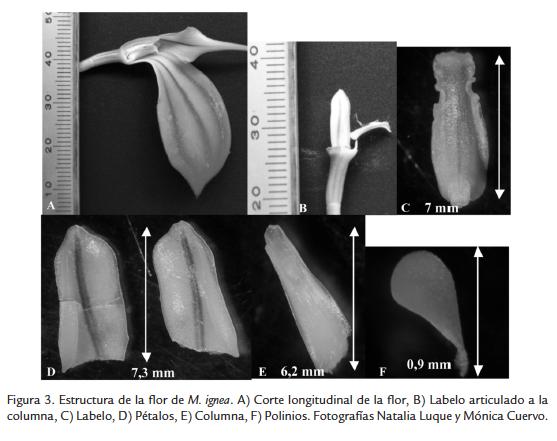
Las partes de las flores con medidas más variables se encontraron en el ancho de la columna, el ancho del pétalo, el ancho del sinsépalo y el largo de la fusión del sinsépalo. Las variables que presentaron mayor coeficiente de variación en M. ignea fueron: el ancho de la columna (148), el ancho del pétalo (32), el ancho del sinsépalo (32), el largo de la fusión del sinsépalo (31) (Tabla 1).
Las variables más altamente correlacionadas fueron: el largo del sépalo dorsal (lsd) y el largo del sépalo lateral (lsl) (R 0,762 p < 0,0001), el ancho del sépalo dorsal (asd) y el largo del sépalo lateral (lsl) (R 0,776 p < 0,0001), el ancho del sépalo lateral (asl) y el largo del sépalo lateral (lsl) (R 0,749 p < 0,0001).
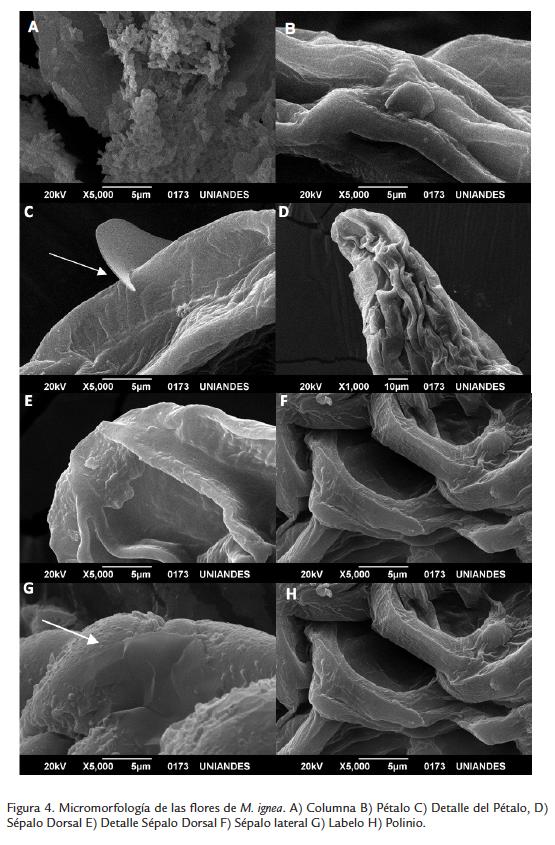
En M. ignea las fotografías de MEB evidenciaron la presencia de tricomas en la superficie de pétalos y sépalos laterales y dorsal (Fig. 4).
Las flores de M. cocccinea y M. ignea no presentaron estructuras secretoras como néctar, aceites, osmoforos u otro tipo de glándulas y los dos controles fueron negativos.
COMPARACIÓN MORFOMÉTRICA DE LAS FLORES DE M. cocccinea Y M. ignea.
En todas las variables analizadas se encontraron diferencias significativas entre las dos especies de orquídeas; a excepción de las variables ancho del sinsépalo (as), ancho del orificio del tubo sepa-lino en la base (aob), ancho del orificio del tubo sepalino en el ápice (aoa) y ancho del labelo en el ápice (ala).
DISCUSIÓN
Las flores de M. coccinea y M. ignea presentaron diferencias en la morfología y morfometría de sus partes, el color de la columna y la longitud del labelo. En todas las variables morfométricas analizadas M. coccinea presentó valores superiores a M. ignea a excepción del ancho del sépalo dorsal, la longitud y el ancho del labelo, la longitud y el ancho del pétalo lateral. Estas diferencias en la morfología y morfometría floral podrían estar afectando la eficiencia de sus polinizadores, posiblemente moscas (Drosophilidae); como vectores de polinios y en últimas la dinámica de remoción y deposición de polinios (Cresswell, 2000; Fetscher, 2001; Fukuda et al., 2001; Cesaro et al., 2004; Fenster et al., 2005).
Por otra parte, estos resultados mostraron una alta variabilidad en los rasgos florales de las especies, los cuales son de las características reproductivas que presentan mayor variación (Cronquist, 1968), así como ausencia de recompensas, lo cual ha sido encontrado frecuentemente en plantas con sistemas de polinización por engaño en comparación con orquídeas que ofrecen recompensas (Juillet y Scopece, 2010). Adicionalmente, se ha planteado que como consecuencia de esta variabilidad floral y de la ausencia de recompensas estas plantas podrían aumentar su éxito reproductivo (Heinrich,1975; Nilsson,1992). Sin embargo, aunque la variabilidad fenotípica en la morfología (Ackerman y Galarza,1991) ha sido ampliamente reportada en orquídeas con polinización por engaño se requieren trabajos adicionales para probar las habilidades cognitivas de los polinizadores y la relación entre la variabilidad fenotípica floral y el éxito reproductivo como lo sugieren Juillet y Scopece, 2010.
El tamaño y la morfología floral en especies simpátricas podrían ser importantes en el aislamiento reproductivo prepolinización (aislamiento morfológico) (Shiestl y Schlüter, 2009), mientras que la variación morfométrica de ciertas piezas florales podría facilitar un mejor acople morfológico entre cada flor y su polinizador. Por esta razón, sería importante analizar a futuro el acople morfológico entre estas dos especies de orquídeas y su polinizadores, como también la importancia de claves visuales referidas a la reflectancia espectral de las diferentes partes florales como estímulos para sus polinizadores (Chittka y Raine, 2006).
Los resultados de la biología floral también mostraron que el labelo y la columna de M. coccinea y M. ignea se encuentran en posición ventral y el labelo es articulado con la columna, estas características son determinantes en el mecanismo de polinización, la forma de remoción y deposición de polinios y afectan el éxito reproductivo de estas especies de orquídeas. Este mecanismo de polinización consiste en que el labelo de las flores de Masdevallia está en un ángulo de 45 ° y cuando el polinizador (Drosophilidae) entra de frente al tubo sepalino, por su peso baja el labelo el cual está articulado a la columna y por esta razón el ángulo aumenta, pero a medida que el insecto entra más en la flor, el labelo recupera su posición original y el polinio se desprende porque al intentar salir el polinizador toca el polinio y este se adhiere a su escutelo y luego es depositado al entrar en otra flor de Masdevallia en la cavidad estigmática que es cóncava (Cuervo, 2012). Este mecanismo de polinización es característico de orquídeas que tienen las anteras incumbentes y el labelo y la columna paralelos (Singer et al., 2008) como es el caso de M. coccinea y M. ignea.
En síntesis, los resultados de los componentes de morfología floral, pruebas histoquímicas y MEB en las flores de M. coccinea y M. ignea mostraron la ausencia de estructuras secretoras como: nectarios, osmóforos u otro tipo de glándulas, indicando que estas especies no ofrecen recompensas alimentarias (néctar, aceites o tejidos) o principios de precursores de feromonas sexuales (fragancias como las que colectan los machos de las abejas euglosinas), lo cual puede sugerir que el sistema de polinización de estas especies sea por engaño como ha sido reportado para otras especies de orquídeas (Van der Pijl y Dodson, 1966; Dressler, 1993; Johnson et al., 1999; Singer, 2002; Singer, 2004; Blanco y Barbosa, 2005; Pellegrino et al., 2005; Schiestl, 2005; Jersakova et al.,2006). Aunque falta aún dilucidar cuál de los sistemas por engaño es el que predomina en estas especies: engaño por alimentación y/o engaño por sitio de refugio. Por otra parte, los polinizadores de estas dos especies de orquídeas (Drosophilidae) las cuales se encuentran en las flores en comportamientos gregarios (Cuervo, 2012), ya han sido reportadas como polinizadores de orquídeas relacionadas como M. floribunda y M. infracta en Brasil (Rodrigo Singer, com pers).
Adicionalmente, las dos especies de orquídeas presentaron flores con colores vistosos (rosado oscuro y rojo oscuro) y produjeron un abundante número de flores. Es decir, la inversión energética en el despliegue floral fue alta, lo cual es congruente con la teoría según la cual las plantas que no ofrecen recompensa usualmente presentan un mayor despliegue floral (Ackerman et al., 2000) y además los colores brillantes amarillo de los polinios contrastan con las coloraciones rojas y muchas orquídeas polinizadas por engaño tienen colores rojos, verde o blancos (Gaskett y Herberstein, 2010).
De acuerdo con los resultados de morfología, morfometría floral, histoquímica y MEB es necesario explorar otros componentes de la biología reproductiva como la fenología, el sistema reproductivo, los polinizadores, las claves visuales y químicas, así como comprobar si se trata de sistemas de polinización por engaño. Esto permitirá una mejor comprensión de las especies y aportará herramientas conceptuales y metodológicas importantes para la conservación de las mismas.
AGRADECIMIENTOS
Al Laboratorio de Investigaciones en Biología Vegetal del Departamento de Biología de la Universidad Nacional de Colombia por el acceso a los equipos. Al Laboratorio de Microscopía Electrónica de Barrido Universidad de los Andes por las fotografías de MEB. A Luis Eduardo Álvarez por el acceso y todo el apoyo brindado para realizar la fase de campo en su finca Villa Rosa en Guasca, Cundinamarca. A la División de Investigación, sede Bogotá de la Universidad Nacional de Colombia por la financiación del proyecto "Biología reproductiva de Masdevallia coccineay Masdevallia ignea en condiciones de semicultivo en Guasca, Cundinamarca" (CÓDIGO: 2010100), convocatoria "Apoyo de la DIB a tesis de investigación en posgrados 2010".
BIBLIOGRAFÍA
ABELE AD. Phylogeny of the Genus Masdevallia Ruiz y Pav. (Orchidaceae) based on Morphological and Molecular Data. [tesis de Doctorado]. Departamento de Biología. Universidad de Hamburg, Hamburg; 2007. [ Links ]
ACKERMAN EJ, ACKERMAN JD, RODRIGUEZ JA. Reproduction in Orchid can be resource-limited over its lifetime. Biotropica. 2000;32(2):282-290. [ Links ]
ACKERMAN JD, GALARZA-PÉREZ M. Patterns and maintenance of extraordinary variation in the Caribbean orchid, Tolumnia (Oncidium) variegata. Syst Bot. 1991;16(1):182-194. [ Links ]
BLANCO MA, BARBOZA G. Pseudopulatory pollination in Lepanthes (Orchidaceae: Pleurothallidinae) by Fungus Gnats. Ann Bot. 2005;95(5):763-772. [ Links ]
CALDERÓN E, FARFÁN J. Especies de los géneros Dracula y Masdevallia en Colombia. B.C. 2003;4(2):187-201. [ Links ]
CESARO AC, BARRETT SCH, VAISSIERE M,THOMPSON JD. An experimental evaluation of self-interference in Narcissus assoanus: Functional and evolutionary implications. J . Evol. Biol. 2004;17(6):1367-1376. [ Links ]
CHAVARRO N. Aspectos reproductivos de la orquídea Masdevallia ignea Rchb.f en condiciones de cultivo en el Jardín Botánico de Bogotá José Celestino Mutis. [tesis de pregrado]. Bogotá: Universidad Militar Nueva Granada; 2005. [ Links ]
CHAVARRO N, BONILLA MA, CURE JR, PACHECO, R. Aspectos reproductivos de la orquídea Masdevallia ignea Rchb.f en condiciones de cultivo en el Jardín Botánico José Celestino Mutis. Pérez Arbelaezia. 2006;16:89-99. [ Links ]
CHITTKA L, RAINE NE. Recognition of flowers by pollinators. Curr Opin Plant Biol. 2006;9:428-435. [ Links ]
CRESSWELL JE. Manipulation of female architecture in flowers reveals a narrow optimum for pollen deposition. Ecology. 2000;81(11):3244-3249. [ Links ]
CRONQUIST A. The evolution and classification of flowering plants. Riverside Studies in Biology. Allen Press, New York; 1968. p. 396. [ Links ]
CUERVO MA. Biología reproductiva de Masdevallia coccineay Masdevallia ignea en condiciones de semicultivo en Guasca, Cundinamarca. [tesis de maestría].Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia. 2012. [ Links ]
DAFNI A, KEVAN P, HUSBAND BC. Practical Pollination Biology, Enviroquest Ltda, Ontario; 2005. p. 231-312. [ Links ]
DAVIES KL, STPICZYNSKA M. Labellar micromorphology of two euglossine- pollinated orchid genera Scutilaria Lindl. and Dichaea Lindl. Ann Bot. 2008;102: 805-824. [ Links ]
DAVIES KL, TURNER MP, GREGG A. Lipoidal labellar secretions in Maxillaria Ruiz y Pav. (Orchidaceae). Ann Bot. 2003;91(4):439-446. [ Links ]
DRESSLER R. Phylogeny and Classification of the Orchid Family. Cambridge University Press; 1993. p. 195-196. [ Links ]
FERNÁNDEZ A, SARMIENTO F. Sinopsis sobre el género Masdevallia en Colombia. Rev Fac Nal Agr Medellín. 1971;27(1):31-42. [ Links ]
FENSTER CB, ARMBRUSTER WS, WILSON P, DUDASH MR, THOMPSON JD. Pollination syndromes and floral specialization. Annu Rev Ecol Syst. 2005;35:375-403. [ Links ]
FETSCHER AE. Resolution of male - female conflict in a hermaphrodite flower. P Roy Soc Lond B Bio. 2001;268:525-529. [ Links ]
FUKUDA Y, SUZUKI K, MURATA J. The function of each sepal in pollinator behavior and effective pollination in Aconitum japonicum var. montanum. Plant Spec Biol. 2001;16(2):151-157. [ Links ]
GASKETT AC, HERBESTEIN ME. Color mimicry and sexual deception by tongue orchids. (Cryptostylis). Naturwissenschaften. 2010;97(1):97-102. [ Links ]
HEINRICH B. Bee flowers: a hypothesis on flower variety and blooming times. Evolution. 1975;29(2):325-334. [ Links ]
INSTITUTO DE HIDROLOGÍA METEOROLOGÍA Y ESTUDIOS AMBIENTALES DE COLOMBIA. Datos multianuales 1990-2010 de temperatura, precipitación y humedad estación climatológica de Guasca; URL: http://institucional.ideam.gov.co/jsp/loader.jsf?lServicio=Usuarios&lTipo=usuarios&lFuncion=login& 2011. [ Links ]
JERSAKOVA J, JOHNSON S, KINDLMANN P. Mechanisms and evolution of deceptive pollination in orchids. Biol Rev. 2006;81:219-235. [ Links ]
JUILLET N, SCOPECE G. Does floral trait variability enhance reproductive success in deceptive orchids? Perspect Plant Ecol. 2010;12(4):317-322. [ Links ]
JOHNSON SD, NILSSON LA. Pollen carryover, geitonogamy, and the evolution of deceptive pollination systems in orchids. Ecology. 1999;80(8):2607-2619. [ Links ]
KNYASEV MS, KULIKOV PV, KNYASEVA OI, SEMERIVOK VL. Interespecific hybridization in northern eurasian Cypripedium: morphometric and genetic evidence of the hybrid origin of C. ventricosum. Lindleyana. 2000;15(1):10-20. [ Links ]
KONIGER W, KONIGER H. Masdevallia. En: Escobar. R (ed.). Orquídeas nativas de Colombia. Sociedad Colombiana de Orquideologia, Editorial Colina, Medellín.1995; 2:268. [ Links ]
LEATHERS JJ. Masdevallia coccinea. Orchids. 2007;76(10):752-755. [ Links ]
LUER CA. Icones Pleurothallidinarum XIX Systematics of Masdevallia, Part one. Monog Syst Botan. 2000;77:1-264. [ Links ]
NILSSON LA. Orchid pollination biology. Trends Ecol Evol. 1992;7(8):255-259. [ Links ]
ORTIZ VP. Orquídeas del género Masdevallia en Colombia. Corporación Capitalina de Orquideologia, Bogotá, D. C.; 2000. p. 172. [ Links ]
ORTIZ VP. Orquídeas colombianas. Segunda edición. Corporación Capitalina de Orquideología, Bogotá D. C.; 1995. p. 320. [ Links ]
PELLEGRINO G, GARGABO D, NOCE ME, MUSACCHIO A. Reproductive biology and pollinator limitation in a deceptive orchid, Serapias vomeracea (Orchidaceae). Plant Spec Biol. 2005;20(1):33-39. [ Links ]
SHIESTL FP. On the success of a swindle: pollination by deception in orchids. Naturwissenschaften. 2005;92(6):255-264. [ Links ]
SHIESTL FP, SCHLÜLER PM. Floral isolation, specialized pollination, and Pollinator Behavior in Orchids. Annu Rev Entomol. 2009;54:425-446. [ Links ]
SINGER RB. The pollination mechanism in Trigonidium obtusum Lindl. (Orchidaceae: Maxillariinae): Sexual mimicry and trap-flowers. Ann Bot London. 2002;89(2):157-163. [ Links ]
SINGER RB, FLACH A, KOEHLER S, MARSAIOLI A, AMARAL MC. Sexual mimicry in Mormolyca ringens (Lindl.) Schltr. (Orchidaceae:Maxillariinae. Ann Bot London. 2004;93(6):755-762. [ Links ]
SINGER R, GRAVEENDEEL B, CRUZ H, RAMIREZ S. The use of orchid pollinia or pollinaria for taxonomic identification. Selbyana. 2008;29(1):16-19. [ Links ]
STENZEL H. Pollen morphology of the subtribu Pleurothalidinae Lindl Orquidaceae. Grana. 2000;39:108-125. [ Links ]
UNEP WCMC. 1-339. UNEP World Conservation Monitoring Centre, Cambridge; 2003. [ Links ]
VAN DER PIJL L, DODSON CH. Orchid flowers and their pollination and evolution. Coral Gables: University of Miami Press; 1966. p. 404. [ Links ]
