










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.18 no.1 Bogotá Jan./Apr. 2013
INTERACCIÓN ENTRE DOS ÁCAROS DEPREDADORES DE Tetranychus urticae KOCH (ACARIFORMES: TETRANYCHIDAE) EN LABORATORIO
Interaction Between Two Predator Mites of Tetranychus urticae Koch (Acariformes: Tetranychidae) in Laboratory
ANGÉLICA ARGÜELLES R1, Bióloga; NATALI PLAZAS1, Bióloga; ALEXANDER BUSTOS R1, Biólogo; FERNANDO CANTOR R.1, Ph. D.; DANIEL RODRÍGUEZ1, Ph. D.; ALEJANDRA HILARION1, M.Sc. 1
1Laboratorio de Control Biológico. Facultad de Ciencias, Universidad Militar Nueva Granada, Colombia. Autor de correspondencia: Fernando Cantor. fernando.cantor@unimilitar.edu.co / ecologia@unimilitar.edu.co. CajicáCundinamarca, Colombia. Kilómetro 2, vía Cajicá – Zipaquirá, Tel.: 57 1 650 00 00, ext: 3269.
Presentado el 26 de julio de 2012, aceptado el 31 de septiembre de 2012, correcciones el 22 de noviembre de 2012.
RESUMEN
Tetranychus urticae (Acari: Tetranychidae) es una de las principales plagas de cultivos ornamentales, entre las especies más utilizadas para su control se encuentran Neoseiulus californicus y Phytoseiulus persimilis (Parasitiformes: Phytoseiidae). En el presente trabajo se propone el manejo de la plaga mediante el empleo de liberaciones simultáneas de los dos fitoseidos. Se evaluaron varias situaciones, por un lado se estudiaron las interacciones cuando un depredador se encuentra en una densidad baja mientras que el otro depredador se presenta en alta densidad (esta situación se analizó tanto en presencia como en ausencia de la presa). Por otro lado, se evaluaron las interacciones cuando los P. persimilis y N. californicus dos están presentes en igual densidad y en presencia de T. urticae. Cuando uno de los depredadores está en mayor densidad y hay presencia de la presa, se observa que al incrementar la edad del depredador que tiene menor densidad, aumenta también la interferencia en el consumo de presas por parte de los depredadores que están en mayor densidad. Además cuando disminuye el consumo de T. urticae se incrementa el consumo intraguilda. Phytoseiulus persimilis en ausencia de T. urticae y en presencia de N. californicus adopta un comportamiento de depredación intraguilda sobre todos los estados de desarrollo de su conespecifico, mientras que N. californicus únicamente consume larvas de conespecíficos en ausencia del fitófago y en presencia de P. persimilis. Cuando se encontraban los dos depredadores en el mismo montaje y la misma densidad de población, no se observó un mayor consumo de T. urticae que cuando cada depredador es empleado por separado.
Palabras clave: control biológico, fitoseidos, intraguilda.
ABSTRACT
Tetranychus urticae (Acari: Tetranychidae) is an important pest of ornamental crops. A species of predatory mite used for its control is Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae). This research proposes the use of joint releases of the two cited predators for the control of the pest. Several situations leading to interaction were evaluated: High density of one predator and low density of the other one, being the prey present or absent. The scenario with predators in equal densities and in presence of the prey was also evaluated. When a predator is in higher density and the prey present, the predator with the lower density increases the interference with the comsumption of preys by the predator with higher density. On the other hand, when the comsumption of T. urticae reduces, intraguild predation increases. P. persimilis shows intraguild predation behaviour when T. urticae is absent and N. californicus is present, consuming all developmental stages of its conspecific. Instead, N. californicus only feed on conspecific larvae, when the fitofagous was absent and P. persimilis was present. When the two predators were present in the same assemblage and with the same population density, the quantity of T. urticae consumed by both of them was not higher than the consumed one when each predator was present in separate way.
Keywords: biological control, phytoseiid, intraguild.
INTRODUCCIÓN
Tetranychus urticae Koch es considerada una de las principales plagas en cultivos de Rosa sp. en la sabana de Bogotá (Acosta, 2010), causando pérdida de calidad, disminución en el crecimiento de las plantas y reducción en la producción de botones florales (Acosta et al., 2001). El alto potencial reproductivo de T. urticae, le permite incrementar su densidad de población en corto tiempo (Mesa et al., 1987; Mendoza et al., 2001), llevando a los productores a elevar las aplicaciones de acaricidas y directamente aumentar los costos de producción. Estos acaricidas generan consecuencias desfavorables para el ambiente y los seres humanos (Santamaría et al., 2002a, Santamaría et al., 2002b). Una alternativa efectiva como componente de un manejo integrado, ha sido la implementación del control biológico mediante el uso de ácaros depredadores pertenecientes a la familia Phytoseiidae (Zhang, 2003). Desde hace varias décadas se ha implementado el uso de Phytoseiulus persimilis como controlador específico de esta plaga, aunque existen otros ácaros depredadores como Neoseiulus californicus, ambos controladores han demostrado ser eficientes, pero actualmente en Colombia no se han realizado estudios sobre la liberación conjunta de estos depredadores, por lo cual, según Schausberger y Walzer (2001), bajo condiciones de invernadero se deben hacer liberaciones iniciales de P. persimilis, con el fin de hacer que la presa decaiga y posterior a esto hacer liberaciones con N. californicus para que este se mantenga presente en el cultivo y controle los individuos de la plaga que no haya consumido P. persimilis.
Phytoseiulus persimilis es un ácaro depredador especialista y Neoseiulus californicus es una especie generalista, esta clasificación es dada según preferencias alimenticias y características morfológicas del depredador (Croft et al., 2004). No obstante, una de las limitantes del control biológico son las diferencias entre las condiciones ambientales del sitio de la introducción al de origen del benéfico, y la residualidad de los productos químicos que pueden interferir con la actividad de los depredadores (Gough, 1991). También es de vital importancia conocer las interacciones entre depredadores, al momento de realizar las liberaciones conjuntas (por las posibles interacciones tróficas que se crean). Tales interacciones, involucran a la planta, los fitófagos y los depredadores y debido a su complejidad no son un aspecto trivial cuando se utiliza el control biológico (Janssen et al., 1998).
Trabajos previos sobre las interacciones en otros depredadores han demostrado que sistemas depredador-depredador y depredador-presa pueden verse afectados en comportamiento y dinámicas de población por competencia, interferencia y depredación intraguilda (Schausberger y Walzer et al., 2001, Onzo et al., 2004, Amarasekare, 2006). La competencia es la interacción interespecífica que se genera entre dos o más especies cuando el recurso es un factor limitante (Price, 1997, Gillott, 2005, Begon et al., 2006), la interferencia es la interacción en donde una de las especies cambia el comportamiento de forrajeo de la otra, en el caso de parasitoides y depredadores, los depredadores pueden consumir hospederos parasitados y por lo tanto detienen el desarrollo del parasitoide. También se puede dar un caso de interferencia en el cual parasitoides de diferentes especies ovipositan en huevos previamente atacados por competidores superiores o gastan mucho tiempo de búsqued a de la presa discriminando entre hospederos sanos y parasitados. Además, la interferencia se presenta entre especies que defienden su territorio o evitan la explotación del recurso en un espacio determinado. En algunos ejemplos también se pueden presentar ambos elementos como explotación e interferencia, cuando se agota el recurso y aumenta la agresión entre los ácaros depredadores con el fin de disminuir la densidad de individuos y así mismo disminuir la competencia y liberación de compuestos químicos es un caso de interferencia (Begon et al., 2006) y la depredación intraguilda hace referencia a especies del mismo nivel trófico que utilizan el mismo recurso y son competidores potenciales que pueden matar, comerse o parasitar (en el caso de parasitoides) a su competidor (Janssen et al., 1998; Rosenheim, 1998).
Los fitófagos también han desarrollado estrategias para evitar la depredación haciendo uso de refugios y/o escapes conocido como comportamientos antidepredadores, inducidos por depredadores (Janssen et al., 1998, Venzon et al., 2000,). En algunas ocasiones las liberaciones conjuntas tanto de depredadores como de parasitoides han generado efectos positivos en el control de la población de la plaga (Onzo et al., 2004), pero en otros casos estas pueden conducir a depredación intraguilda afectando el control de la plaga (Walzer y Schausberger, 1999,Venzon et al., 2001, Schausberger y Walzer et al., 2001). También puede presentarse interferencia, por la liberación de compuestos químicos emitidos por la presencia de alguno de los depredadores (Guillott, 2005), esta interferencia se puede dar por comportamientos territoriales. Por lo tanto, es importante conocer todas las interacciones generadas en un sistema trófico para mejorar la efectividad del control biológico, conociendo por completo la biología y ecología de cada uno de los individuos que en este sistema interactúan. El objetivo del presente trabajo fue evaluar las interacciones entre los ácaros depredadores N. californicus y P. persimilis que puedan afectar la efectividad del control de Tetranychus urticae cuando los dos depredadores actúan simultáneamente sobre la misma población del fitófago y determinar si se presenta depredación intraguilda en ausencia de T. urticae.
MATERIALES Y MÉTODOS
El estudio se realizó en el laboratorio de Control Biológico de la Universidad Militar Nueva Granada, ubicado en Cajicá, Cundinamarca, bajo condiciones controladas a una temperatura promedio de 25 ± 1 ºC y 55 ± 5 % de humedad relativa. Para la obtención de los ácaros depredadores se llevó a cabo una cría bajo condiciones controladas siguiendo una modificación de la metodología propuesta por Mesa y Bellotti (1987), en cajas transparentes de 30 x 20 x 25 cm con tapa semihermética, provistas de un orificio central para ventilación de la caja. En el interior se dejaron foliolos de fríjol (Phaseolus vulgaris L.) variedad cerinza (que contenían los ácaros presa), utilizados para alimentar a P. persimilis y N. californicus. La producción de T. urticae se llevó a cabo en un sistema de cría masiva en campo, donde se infestaron plantas de fríjol de cinco semanas de edad, luego de emergidas las hojas verdaderas y se recolectaron para los respectivos montajes (Argüelles et al., 2006).
Se utilizó un diseño de bloques al azar con 17 tratamientos, donde se realizaron combinaciones de las dos especies de depredadores, con el fitófago actuando cada uno de ellos por aparte o en conjunto y los dos depredadores sin la presa en conjunto, para cada tratamiento se realizaron seis repeticiones. Las unidades experimentales consistían en una caja de Petri de 15 cm de diámetro por 2 cm de altura. En el fondo se ubicó un trozo circular de papel absorbente humedecido del tamaño de la base de la caja, un foliolo de fríjol como sustrato y se colocó el número y edad de los individuos correspondientes a cada tratamiento con la ayuda de un pincel sobre cada foliolo. Las cajas se cubrieron con papel vinipel transparente para evitar que los ácaros se escaparan. Para todos los tratamientos, se observó el número de presas y depredadores muertos por depredación con un estereoscopio (cada cuatro horas), por un lapso total de 20 h. No se reemplazaron individuos consumidos. El número de huevos consumidos fueron contados cada 4 h, restando la densidad inicial (20 o 30 huevos según tratamiento), al número de huevos presentes en el momento de evaluación, de acuerdo con lolo recomendado por Fernando y Hassell (1980). Para las otras edades únicamente se registro el número de individuos succionados (depredados).
PREFERENCIA ALIMENTICIA
Para estimar las preferencias alimenticias de los depredadores sobre las diferentes edades de la presa, se utilizó un diseño de bloques al azar con cinco tratamientos y seis repeticiones. Para cada tratamiento se usó una proporción de un depredador y 80 presas discriminadas en 20 individuos por edad del fitófago (huevo, larva, ninfa y adulto). Los tratamientos se diferencian entre sí, según la edad de los dos depredadores a evaluar (Tabla 1). Las larvas de P. persimilis fueron excluidas de los tratamientos porque según Zhang (2003) y Gotoh et al. (2004), estas no tienen un comportamiento de depredación en este estado. Se hicieron conteos de los individuos depredados, con una regularidad de 4 h durante 20 h.

EVALUACIÓN DE LOS DEPREDADORES SOBRE T. urticae A LA MISMA DENSIDAD
Se evaluaron los efectos de P. persimilis y N. californicus sobre poblaciones de T. urticae en el mismo sistema. Para estos tratamientos se utilizaron ninfas o adultos de ambas especies y 120 presas por tratamiento. Se comparó el consumo total por parte de los fitoseidos cuando actúan únicamente sobre el fitófago y también cuando están en presencia de un depredador de la misma edad, contando en cada caso el número de presas consumidas en un periodo de 20 horas (Tabla 2).
EVALUACIÓN DE LOS DEPREDADORES EN PRESENCIA Y AUSENCIA DE T. urticae
Se evalúo el consumo de P. persimilis y N. californicus sobre los diferentes estados de T. urticae y y en presencia y ausencia del mismo. Se utilizaron cinco tratamientos y seis repeticiones para cada uno de estos. En presencia de T. urticae se colocaron 81 fitoseidos (80 individuos de un depredador + 1 individuo del otro depredador) en una proporción de 1:1 para el depredador de mayor población con respecto a T. urticae. Para evaluar cómo la presencia de los depredadores en el mismo sistema afectan el consumo sobre T. urticae, al compararse con los consumos de cada depredador actuando independiente y a la misma densidad. Para conocer la depredación intraguilda de N. californicus y P. persimilis en ausencia del fitófago, los montajes consistían en un depredador en edad ninfa o adulto para P. persimilis y 80 individuos de N. californicus, considerando todas las edades de T. urticae estaban discriminadas en 20 huevos, 20 larvas, 20 ninfas y 20 adultos para un total de 80 individuos por tratamiento. Con N. californicus se tomaron larvas, o ninfas, o adultos con 80 individuos de P. persimilis, con la misma distribución que P. persimilis. Se registró el número de depredadores de la misma especie consumidos cada 4 h, durante 20 h.
ANÁLISIS ESTADÍSTICO
Se realizaron los análisis requeridos para comparar el consumo de los dos depredadores en presencia o ausencia de T. urticae y para efectuar las comparaciones del efecto del consumo de los depredadores individualmente o en conjunto. Se realizaron análisis de varianza y pruebas de comparación múltiple de Tukey para evaluar las diferencias entre tratamientos, en aquellos casos en que los residuales presentaron distribución normal. En los casos en los cuales no fue posible obtener una aproximación razonable de los residuales a la distribución normal, se utilizaron pruebas no paramétricas de Kruskall Wallis. Los análisis se realizaron con el software estadístico R 2.6.0 de libre distribución (http://www.rproject.org/).
RESULTADOS
PREFERENCIA ALIMENTICIA
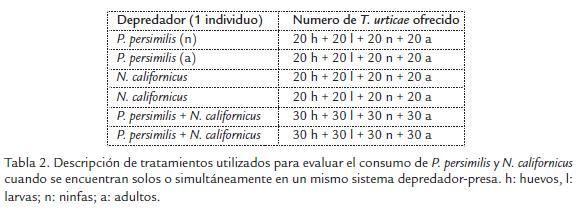
Se observó que el consumo total del fitófago fue en promedio de 14 ± 0,3 individuos por parte de ninfas y adultos de P. persimilis. Mientras que ninfas y adultos de N. californicus consumieron en promedio 17 ± 2.2 y 15 ± 1.0 individuos respectivamente. Sin embargo, no se presentaron diferencias estadísticamente significativas (p > 0.05) cuando se comparó el consumo de individuos de T. urticae entre ninfas y adultos de un mismo depredador. De la misma forma, no se presentaron diferencias en el consumo de presas cuando se comparó el número de individuos de T. urticae depredados en las edades de ninfa y adulto entre depredadores (p > 0.05). En términos de preferencias por edad de la presa, por cada una de las edades de los dos depredadores se observó que ninfas de P. persimilis consumieron más huevos (p = 0,030) y larvas (p = 0,007) que adultos de T. urticae (Fig. 1) lo cual coincide con lo reportado por Takafuji y Chant (1976). Lo anterior demuestra una preferencia de las ninfas del depredador hacia estados inmaduros a pesar que pueden consumir estados maduros. Aunque los adultos del depredador no presentaron diferencias significativas (p = 0,179) en la preferencia por alguna de las edades de la presa, se observa un mayor número de individuos consumidos en edades de huevo y larva. El consumo por parte de ninfas de N. californicus no presentó una preferencia hacia alguna de las edades de T. urticae (p = 0,09). Sin embargo, cuando se encontraba en edad adulta tuvo un mayor consumo sobre larvas y ninfas (p = 0,008), lo anterior concuerda con lo observado por Forero (2008), donde se obtuvieron consumos superiores hacia larvas (18) y ninfas (19) que hacia adultos (3,4) de T. urticae cuando N. californicus lo controlaba.
EVALUACIÓN DE LOS DEPREDADORES SOBRE T. urticae A LA MISMA DENSIDAD
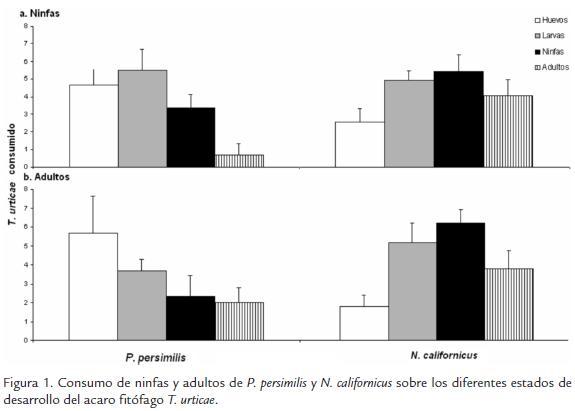
En el caso de ninfas de P. persimilis y N. californicus a la misma densidad, se presentó un consumo promedio de 10 ± 2.5 huevos, 7 ± 0.9 larvas, 7 ± 0.6 ninfas y 8 ± 0.8 adultos, siendo el consumo sobre larvas y ninfas menor que el consumo registrado cuando los dos depredadores interactúan con su presa en sistemas independientes. Para adultos se registró un menor consumo de 4 ± 0.8 huevos, 6 ± 1.0 larvas, 6 ± 1.2 ninfas y 5 ± 0.9 adultos, en este caso el consumo sobre huevos es menor en sistemas donde P. persimilis se encuentra con T. urticae que cuando actúa independiente (Fig. 2).
EVALUACIÓN DEL CONSUMO DE N. californicus Y P. persimilis EN PRESENCIA DE T. urticae
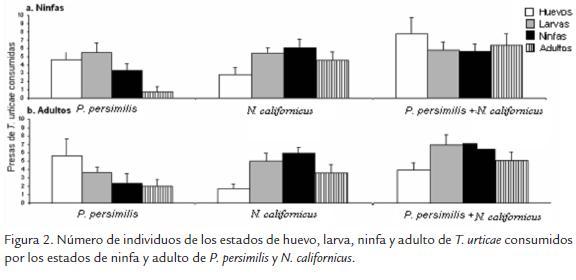
La presencia de un segundo depredador generó una reducción del consumo de T. urticae por parte del depredador que se encontraba en mayor proporción. En el caso de N. californicus en proporción 1:1 con T. urticae y un solo individuo de su P. persimilis (una ninfa o un adulto), se observó una disminución en el consumo total de individuos. N. californicus en presencia de un adulto de P. persimilis consume 30 ± 4,4 individuos de T. urticae y en presencia de una ninfa de P. persimilis fue de 50 ± 2,4 individuos (p = 0,003). En la figura 3 se observa que al incrementar la edad de P. persimilis se presenta mayor interferencia y menor consumo de N. californicus sobre T. urticae. El comportamiento anterior fue observado con P. persimilis cuando se encontraba en proporción 1:1 con T. urticae y en presencia de larvas, ninfas o adultos de N. californicus. El adulto de N. californicus generó una mayor interferencia sobre el consumo de P. persimilis comparado a la interferencia que generan larvas (p = 0,0002) o ninfas (p = 0,003) de N. californicus sobre el consumo de P. persimilis.
DISCUSIÓN
PREFERENCIA ALIMENTICIA
El consumo por parte de larvas fue menor en relación al consumo por ninfas y adultos en el caso de N. californicus. Este comportamiento posiblemente pueda atribuirse a las diferentes demandas nutricionales y energéticas de los depredadores dependiendo de la edad, como lo reportan Sabelis (1985) y Fernando y Hassell (1980), puesto que los depredadores en edades adultas tienen mayores exigencias metabólicas y menores tiempos de manipulación, lo que tiene como resultado un mayor número de presas consumidas.
EVALUACIÓN DE LOS DEPREDADORES SOBRE T. urticae A LA MISMA DENSIDAD
Debido a las preferencias encontradas por las diferentes edades de la presa, se puede pensar que en el momento de una liberación en conjunto los depredadores tendrían un efecto sinérgico, presentándose un consumo superior en la edad donde las preferencias por parte de ambos depredadores se superponen (larvas), también se esperaría que al estar los dos depredadores en el mismo sistema los valores de consumo sobre huevos, larvas, ninfas y adultos fueran el resultado de la suma del consumo de las dos especies cuando actúan por aparte, pero esto no sucedió en el sistema con ambos depredadores. De acuerdo a los resultados obtenidos se observó que estos resultados son similares a los obtenidos por Schausberger y Walzer (2001) cuando realizaban liberaciones en conjunto o individuales de P. persimilis y Neoseiulus californicus en gérbera.
Por lo anterior, se puede afirmar que a pesar de que se presentan diferencias por la preferencia de la edad de T. urticae, por parte de P. persimilis y N. californicus cuando atacan independientemente, el efecto de la liberación conjunta de los dos depredadores no permite un control más alto que el obtenido con liberaciones de cada uno por separado. El único estado del fitófago en que se observa un consumo aproximadamente aditivo para los dos depredadores al hacer la liberación conjunta, es el estado adulto de la plaga. La interferencia es una interacción que afecta el consumo entre depredadores disminuyendo la cantidad de presas atacadas y consumidas (Price, 1997; Gillott, 2005), siendo posiblemente el motivo por el cual no se presentó un efecto acumulativo de consumo cuando los depredadores se encontraban atacando a T. urticae en conjunto. Resultados similares fueron obtenidos por Onzo et al. (2004), quienes realizaron experimentos donde los depredadores Typhlodromalus aripo De Leon y Typhlodromalus manihoti (Moraes) fueron usadas para controlar independientemente y en conjunto poblaciones de Mononychellus tanajoa (Bondar) en yuca. En este caso tampoco se observo un efecto aditivo del consumo del fitófago al compararse con los consumos de cada depredador cuando se encontraba actuando sobre su presa en ausencia del otro depredador.
EVALUACIÓN DEL CONSUMO DE N. californicus Y P. persimilis EN PRESENCIA DE T. urticae
Se generó un consumo intraguilda entre los depredadores por falta de suficientes recursos, similar a lo reportado por Schausberguer y Croft (2000). Esto posiblemente se debe a que estados móviles del fitófago buscan refugios para evitar ser depredados, pues T. urticae tiene la capacidad de reconocer conespecíficos muertos y evitar estas áreas de alimentación, dejando vulnerables al ataque y consumo estados inmóviles como huevos o quiescentes (Venzon et al., 2000).
Walzer et al. (2001) reportan depredación intraguilda para P. persimilis y N. californicus. Esto es usual en artrópodos generalistas, los cuales no solo atacan las poblaciones de la plaga, sino a otros enemigos naturales (cuando las presas tradicionales no se encuentran dentro del sistema). Se ha observado que la densidad de la plaga incrementa cuando dos especies de depredadores coexisten por causa de la disminución de la población de los depredadores o parasitoides, debido al consumo intraguilda de estados inmaduros (Janssen et al., 1998). En el caso de P. persimilis y N. californicus los dos prefieren consumir larvas y ninfas de T. urticae que huevos, por la dificultad de penetrar el corión de los huevos (Walzer y Schausberger, 1999). Phytoseiulus persimilis en ausencia de T. urticae y en presencia de las tres edades de N. californicus consume todos los estados de coespecificos como se observa en la figura 4. Walzer y Schausberguer (1999) reportan comportamiento de canibalismo por parte de ninfas y protoninfas de P. persimilis sobre estados inmaduros en ausencia de T. urticae. Los mismos autores argumentan que P. persimilis es un depredador especialista, que en ausencia del fitófago depreda a conespecíficos, en lugar de consumir otro tipo de presa diferente como Neoseiulus californicus. Lo anterior coincide con posteriores estudios realizados por Schausberger y Croft, (2000) donde se evaluaron especies generalistas y especialistas en relación al consumo intraespecífico en ausencia de las presas acostumbradas, dando como resultado consumos intraespecíficos para los dos tipos de depredadores sobre estados de huevo y larva.
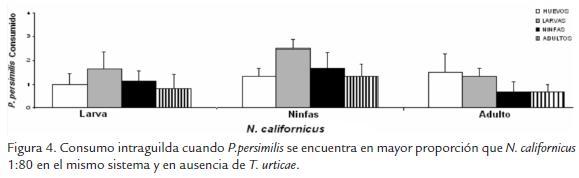
Huevos y larvas de N. californicus fueron consumidos por parte de conespecíficos en presencia de una ninfa de P. persimilis y se reportó un consumo de larvas únicamente en presencia de un adulto de P. persimilis (Fig. 5). Walzer y Schausberger (1999) sugieren que este comportamiento se encuentra asociado a la capacidad que presentan los ácaros especialistas para discriminar entre conespecíficos y heteroespecíficos, generando un mayor consumo de los conespecíficos de su misma especie, al contrario de las especies generalistas que prefieren consumir heteroespecíficos. Al ser considerado P. persimilis una especie especialista y N. californicus una especie generalista se puede observar un mayor consumo por parte de P. persimilis sobre sus conespecificos, al compararse la depredación intraguilda de N. californicus (Fig. 4), y la de P. persimilis (Fig. 5) (Croft et al., 2004). Esto puede deberse a que especies generalistas pueden permanecer por más tiempo en ayuno que especies especialistas. Por lo tanto, la tendencia al canibalismo es menor en N. californicus que en P. persimilis similar a los resultados reportados por Schausberger (2003) en N. californicus y P. persimilis.

CONCLUSIONES
Cuando P. persimilis y N. californicus se encuentran en el mismo sistema se presenta una disminución en el consumo de T. urticae, debido a la interferencia generada por cada depredador entre sí, principalmente en edad adulta. Sin embargo, no cambia la preferencia alimenticia de cada depredador por los estados de T. urticae.
AGRADECIMIENTOS
Agradecemos a las Universidad Militar Nueva Granada por la financiación del proyecto del cual se derivo esta investigación.
BIBLIOGRAFÍA
ACOSTA A. Biología, ecología y manejo de ácaros en cultivos de flores en: Memorias del Simposio de actualización en el control de ácaros y Trips. 2010. [ Links ]
ACOSTA O, YEPES M, ACOSTA A, DÍAZ L. Ciclo biológico de Tetranychus cinnabarinus Boisduval y Tetranychus urticae Koch en rosa, clavel y alstroemeria bajo siete ambientes. Asocol. 2001;60:25-39. [ Links ]
AMARASEKARE P. Productivity,dispersal and the coexistence of intraguild predators and prey. J Theor Biol. 2006;243(1):121-133. [ Links ]
ARGÜELLES A, PLAZAS N, TÁUTIVA L. Evaluación de un método a campo abierto para la producción de Tetranychus urticae- Koch (Acari: Tetranychidae), como suministro de presas para Phytoseiulus persimilis- Athias. Rev Fac Cien Basic. 2006;2(1):117-122. [ Links ]
BEGON M, TOWNSEND C, HAPER JL. Ecology from individuals to ecosystems. Blackwell Publishing.Malden. USA. 754. 2006. p. 227-297 [ Links ]
CROFT BA, BLACKWOOD JS, MCMURTRY JA. Classifying life-style types of phytoseiid mites: diagnostic traits. Exp Appl Acarol. 2004;33(4):247-260. [ Links ]
FERNANDO MH, HASSELL MP. Predatory-prey responses in an acarine system. Popul Ecol. 1980;22(2):301-322. [ Links ]
FORERO G. Evaluación de Amblyseius sp. (Acari: Phytoseiidae) enemigo natural de Tetranychus urticae (Acari: Tetranychidae) en cultivo de rosa. Agro Col. 2008;26(1):78-86. [ Links ]
GILLOTT C. Entomology. Third edition. Springer. Canada. 2005. p.705-711. [ Links ]
GOTOH T, NOZAWA YAMAGUCHI, K. Prey consuption and funtional response of three acarophagous species to eggs of the two-spotted spider mites in the laboratory. Appl Entomol Zool. 2004;39(1):97-105. [ Links ]
GOUGH N. Long term stability in the interaction between Tetranychus urticae and Phytoseiulus persimilis producing succesful integrated control on roses in southest Quesland. Exp Appl Acarol. 1991;12:83-101. [ Links ]
JANSSEN A, PALLINI A, VENZON M, SABELIS M. Behaviour and indirect interactions in food webs of plant-inhabiting arthropods. Exp Appl Acarol. 1998;22:497-591. [ Links ]
MENDOZA LA, ACOSTA A, OSPINA J, DÍAZ L. Evaluación de diferentes estados de ácaros del genero Tetranychus, en cultivos de flores de la Sabana de Bogotá Asocol. 2001;60:11-24. [ Links ]
MESA NC, BELLOTTI A, DUQUE M. Tablas de vida de Mononychellus progresivus Doreste y Tetranychus urticae Koch (Acarina: Tetranychidae) en yuca. Rev Col Entomol. 1987;13(2):11-22. [ Links ]
ONZO AR, HANNA R, JANSSEN A, SABELIS M. Interactions between two neotropical phytoseiid predators on cassava plants and consequences for biological control of a shared spider mite prey: a screenhouse evaluation. Biocontrol Sci Technol. 2004;14(1):63-76. [ Links ]
PRICE P.W. Insect ecology. 3ra Edición. John Wiley & Sons. Inc. Toronto, Canada. 1997. ROSENHEIM JA. Higherorden predators and the regulation of insect herbivore populations. A Rev Entomol. 1998;43:421-447. [ Links ] [ Links ]
SABELIS MW. Predator-Prey Interaction. En: Helle W, Sabelis MW. Eds. Spider Mites Their Biology, Natural Enemies and Control. Elsevier. Amsterdam; 1985. p. 285-297 [ Links ]
SANTAMARÍA G, SALAS C, ACOSTA A, AMADOR J. Caracterización de la ubicación de focos de ácaros fitófagos dentro de invernaderos, para cultivos de rosa, clavel y alstroemeria. Rev Asocol. 2002a;63:19-27. [ Links ]
SANTAMARÍA G, SALAS C, ACOSTA A, AMADOR J. Estudio de la disposición vertical y horizontal de Tetranychus spp. en plantas de rosa, clavel y alstroemeria. Rev Asocol. 2002b;63:44-60. [ Links ]
SCHAUSBERGER P, CROFT B.A. Cannibalism and intraguild preation among phytoseiid mites: are agressiveness and prey preference related to diel specialization? Exp Appl. Acarol. 2000;24(9):709-725. [ Links ]
SCHAUSBERGER P, WALZER A. Combined versus single species of predaceous mites: Predatorpredator interactions and pest suppression. BioControl. 2001;20(3):269-278. [ Links ]
SCHAUSBERGER P. Cannibalism among phytoseiid mites: a review. Exp Appl Acarol. 2003;29(3-4):173-191. [ Links ]
TAKAFUJI A, CHANT D. Comparative studies of two species of predacious phytoseiid mites (Acarina: Phytoseiidae), with special reference to their responses to the density of their prey. Res Popul Ecol. 1976;17:255-310. [ Links ]
VENZON M, JANSENN A, PALLINI A, SABELIS M. Diet of a poliphagous predator affects refuge seeking of its thrips prey. Anim. Behav. 2000;60(3):369-375. [ Links ]
VENZON M, PALLINI A, JANSSEN A. Interactions mediated by predators in arthropod food webs. Neotrop Entomol. 2001;30(1):1-9. [ Links ]
WALZER A, BLÜMEL S, SCHAUSBERGER P. Population dynamics of interacting predatory mites, Phytoseiulus persimilis and Neoseiulus californicus, held on detached bean leaves. Exp Appl Acarol. 2001;25(9):731-743. [ Links ]
WALZER A, SCHAUSBERGER P. Cannibalism and interspecific predation in the phytoseiid mites Phytoseiulus persimilis and Neoseiulus californicus: predation rates and effects on reproduction and juvenile development. Biocontrol. 1999;43(4):457-458. [ Links ]
ZHANG Z. Mites of greenhouses: identification, biology and control. Cambridge. USA. CABI publishing; 2003. p. 171-194. [ Links ]
