










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.18 no.2 Bogotá May/Aug. 2013
TAXOCENOSIS DE MOLUSCOS Y CRUSTÁCEOS EN RAÍCES DE Rhizophora mangle (RHIZOPHORACEAE) EN LA BAHÍA DE CISPATÁ, CÓRDOBA, COLOMBIA
Taxocenosis of mollusks and crustaceans on roots of Rhizophora mangle (Rhizophoraceae) at Cispatá Bay, Córdoba, Colombia
JORGE ALEXANDER QUIRÓS R1 , M.Sc.; JORGE ENRIQUE ARIAS R1 , M.Sc.
1 Universidad de Córdoba. Facultad de Ciencias Básicas, Departamento de Biología, Montería, Colombia. Autor de correspondencia: Jorge Quirós Rodríguez. scaride@yahoo.com;alexander_quiroz@hotmail.com
Presentado el 9 de mayo de 2013, aceptado el 30 de mayo de 2013, correcciones el 18 de junio de 2013.
RESUMEN
A pesar del conocimiento existente sobre la ecología de los manglares en la bahía de Cispatá, pocos estudios han sido enfocados en la asociación de invertebrados en las raíces del mangle rojo, por lo que entre diciembre 2010 y septiembre 2011, se caracterizó la taxocenosis de moluscos y crustáceos en raíces de Rhizophora mangle en dos sectores de muestreo de la bahía de Cispatá, Colombia. Para la recolección del material biológico se tomaron al azar tres raíces de mangle homogéneas en cuanto al diámetro. Los moluscos y crustáceos fueron obtenidos de la raíz raspando la superficie con un cuchillo, luego fueron separados y fijados en formalina al 10 % para su posterior identificación hasta especie mediante claves taxonómicas especializadas. De los 12289 individuos recolectados en los cuatro muestreos, 10470 pertenecieron al phylum Mollusca (85,2 %) y los restantes 1819 al subphylum Crustacea (14,8 %). De moluscos se identificaron 14 especies distribuidas en 11 familias y dos clases; Bivalvia y Gastropoda. De crustáceos se identificaron 24 especies distribuidas en 16 familias y cuatro órdenes; Sessillia, Decapoda, Isopoda y Amphipoda. En los dos sectores de muestreo Mytella charruana, Balanus eburneus y Crassostrea rhizophorae fueron las especies más importantes en términos de abundancia, no obstante se sabe que moluscos como M. charruana y B. eburneus presentan una gran capacidad de adaptación y ajuste a las variaciones hidroclimáticas, lo que se reflejó en la dominancia de dichas especies en el sector con mayor influencia del río Sinú. La presencia de los crustáceos Petrolisthes armatus y Aratus pisonii en el sector con más cercanía al mar Caribe, indica que son especies de bosques de manglar con gran movilidad y mecanismos de adaptación fisiológicos.
Palabras clave: crustáceos, mar Caribe, moluscos, Rhizophora mangle, río Sinú.
ABSTRACT
Despite the existing knowledge on the ecology of mangroves at Cispatá Bay, few studies have focused on the association of invertebrates on red mangrove roots, so between December 2010 and September 2011, it was characterized taxocenosis of mollusks and crustaceans on roots of Rhizophora mangle in two study sites at Cispatá Bay, Colombia. For the collections of biological material were randomly taken three homogeneous mangrove roots in diameter. Mollusks and crustaceans were obtained from the root surface with a scraping knife then were removed and fixed in 10 % formalin for later identification to species using specialized taxonomic keys. Of the 12289 individuals collected in the four samples, 10470 belonged to the phylum Mollusca (85,2 %) and the remaining 1819 to subphylum Crustacea (14,8 %). For mollusks were identified 14 species distributed in 11 families and two classes; Bivalvia and Gastropoda. For crustaceans were identified 24 species distributed in 16 families and four orders; Sessillia, Decapoda, Isopoda and Amphipoda. In both sectors sampling Mytella charruana, Balanus eburneus and Crassostrea rhizophorae were the most important species in terms of abundance, however mollusks like M. charruana and B. eburneus have a great ability to adapt and adjust to changing hydroclimatic, which was reflected in the dominance of these species in the sector with the greatest influence Sinu River. The presence of crustaceans Petrolisthes armatus and Aratus pisonii in the sector with more proximity to the Caribbean Sea indicate that are species with great mobility and physiological adaptation mechanisms.
Key words: Caribbean Sea, Crustaceans, Mollusks, Rhizophora mangle, Sinu River.
INTRODUCCIÓN
La bahía de Cispatá se caracteriza por ser un ambiente estuarino, delimitada en su mayoría por extensas coberturas de manglar, la cual recibe el aporte directo de agua dulce por varios efluentes del río Sinú y presenta la influencia directa del mar Caribe; estos aportes se encuentran influenciados por los periodos climáticos, así como por la hidrodinámica del mar (Ruíz et al., 2008; Castaño et al., 2010). Dentro de este ecosistema, Rhizophora mangle constituye un recurso importante, debido a que mantiene en sus raíces sumergidas una gran biodiversidad epibionte dominada principalmente por moluscos y crustáceos (Carballo, 2000; Dittmann, 2001). Estos invertebrados se caracterizan por desarrollar mecanismos de adaptación a las variaciones del ambiente, además de resistir la desecación e inmersión. Así mismo, presentan hábitos errantes, incrustantes, perforadores y vágiles (Reyes y Campos, 1992a), y pueden mostrar gradientes en la zonación sobre las raíces (Ellison y Farnsworth, 1990).
Los moluscos que viven asociados a las raíces del mangle rojo, aprovechan la materia orgánica en suspensión como fuente directa de alimento. Luego son capturados por otros niveles tróficos superiores, contribuyendo así con el flujo de energía del sistema (Reyes y Campos, 1992b). Esta comunidad está formada por gasterópodos de hábitos detritívoros, herbívoros y ramoneadores (Farnsworth y Ellison, 1996). También es común encontrar otros moluscos de hábitos filtradores como las ostras y mejillones (Lacerda et al., 2001). Los crustáceos se distribuyen sobre la porción sumergida, media e inferior de las raíces de mangle, estando representados la mayoría de los taxones, desde anfípodos, isópodos y decápodos donde se asocian a una gran variedad de microhábitats generados por estas raíces, hasta cirripedios que forman agregados y colonizan una porción importante de las raíces (Romero y Polanía, 2008).
En el Caribe colombiano existen algunos trabajos relacionados con los organismos asociados a los manglares, entre estos cabe mencionar a Reyes y Campos (1992a) y Reyes y Campos (1992b) quienes evaluaron los macroinvertebrados asociados a las raíces de Rhizophora mangle en la región de Santa Marta y la bahía de Chengue. Patiño y Flórez (1993) y Pérez (1998), caracterizaron el ecosistema de manglar en el golfo de Morrosquillo; Báez (2001), estudió la estructura de la biocenosis asociada a raíces sumergidas de mangle rojo en la isla Tintipán, archipiélago de San Bernardo; Romero y Polanía (2008), evaluaron la sucesión temprana de anélidos, moluscos y crustáceos asociados a las raíces de mangle rojo en la isla de San Andrés. Así mismo, Quiceno y Palacio (2008), realizaron un estudio de los macroinvertebrados asociados a las raíces sumergidas de R. mangle en la ciénaga La Boquilla, Sucre. Finalmente, García y Palacio (2008), determinaron las características taxonómicas de la fauna de macroinvertebrados del mangle rojo (R. mangle) en dos bahías de la costa oriental del golfo de Urabá.
Debido a la gran importancia bioecológica que presenta la bahía de Cispatá y las escasas investigaciones relacionadas con la asociación de invertebrados marinos a las raíces de R. mangle, se planteó como objetivo caracterizar la taxocenosis de moluscos y crustáceos en raíces del mangle rojo en dos sectores de la bahía de Cispatá, teniendo en cuenta en un sector la cercanía del mar Caribe y en el otro la influencia del río Sinú.
MATERIALES Y MÉTODOS
Área de estudio
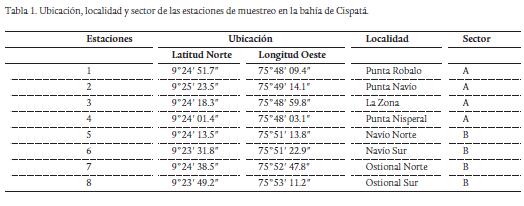
La bahía de Cispatá está ubicada en el litoral Caribe colombiano, específicamente en el departamento de Córdoba, entre los 9° 20'- 9° 25' norte y 75° 47'- 75° 55' oeste, al costado suroccidental del golfo de Morrosquillo, abarcando los municipios de San Antero y San Bernardo del Viento (Fig. 1). En el área de estudio se muestra un régimen climático unimodal, en donde se observa una temporada seca de diciembre a abril y una temporada de lluvias de abril a noviembre. Sin embargo, abril y diciembre se consideran meses de transición (Patiño y Flórez, 1993). En el área de estudio, se eligieron dos sectores de muestreo (sector A y sector B) con ocho estaciones apropiadamente geoposicionadas, utilizando un GPS marca Garmin (± 15 m) (Tabla 1). En cada sector se efectuaron muestreos trimensuales entre diciembre 2010 y septiembre 2011.
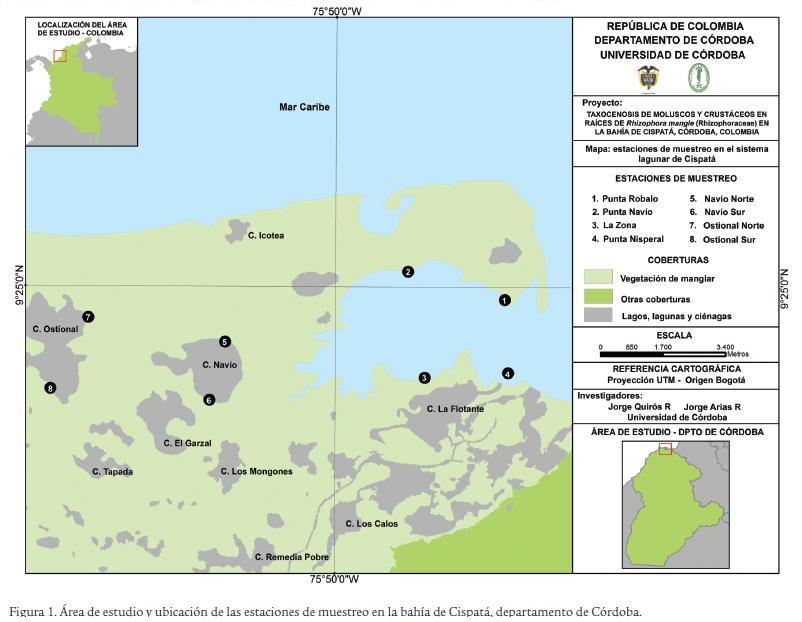
Descripción de los sectores de muestreo
El sector A, con mayor cercanía del mar Caribe, corresponde a la ciénaga de La Zona, constituido por un bosque maduro y mixto, principalmente de R. mangle, Conocarpus erectus y Laguncularia racemosa, la especie dominante en el borde de la ciénaga es R. mangle. El sector B, con mayor influencia del río Sinú, está formado por las ciénagas del Ostional y Navío, el cual se ubica en la parte interna del sistema estuarino. La ciénaga del Ostional presenta un suelo inundable y en general está constituida por un bosque joven y monoespecífico de R. mangle, mientras que la ciénaga de Navío presenta un suelo en general firme e igualmente inundado, en donde se desarrolla un bosque maduro y poco intervenido, dominado principalmente por R. mangle.
Fase de campo
Para la recolección del material biológico se tomaron al azar tres raíces sumergidas de mangle rojo con un diámetro homogéneo por estación. Una vez seleccionada la raíz, se introdujo por su parte inferior en una bolsa de lona previamente rotulada. En cada estación, se procedió a lavar las raíces con agua dulce para lograr la relajación y separación de los individuos adheridos a las raíces; paso seguido se dispuso las raíces en bandejas plásticas para evitar la pérdida de organismos (modificado de Márquez et al., 2006). Los moluscos y crustáceos se separaron del resto de los invertebrados y se colocaron en bolsas de plástico rotuladas, luego las muestras se almacenaron en frío dentro de una nevera de icopor (Márquez y Jiménez, 2002), hasta su llegada al laboratorio de Zoología de la Universidad de Córdoba.
Fase de laboratorio
En esta fase los moluscos y crustáceos fueron fijados en formalina al 10 % (Romero y Polanía, 2008). La identificación taxonómica de los crustáceos se realizó con la ayuda de las claves de Rathbun (1930), Williams (1984) para los decápodos; Barnard (1969) y Ortiz (1994) para los anfípodos; Kensley y Schotte (1989) para los isópodos; Granadillo y Urosa (1984) para los cirrípedos. Para la identificación de los moluscos se emplearon las claves de Abbott (1974), Warmke y Abbott (1975), Díaz y Puyana (1994), Lodeiros et al. (1999) para los bivalvos y gastrópodos.
Análisis de los datos
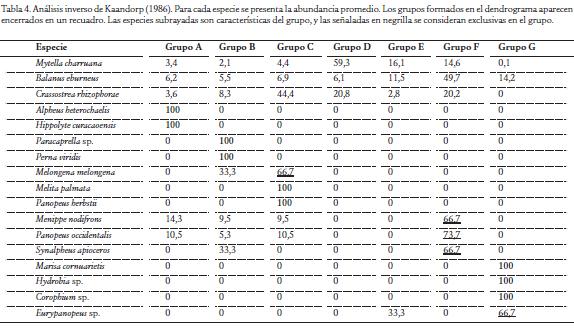
Para caracterizar la taxocenosis de moluscos y crustáceos en las estaciones de estudio y en los meses de muestreo, se elaboraron matrices de abundancia transformada con raíz cuarta, las cuales fueron sometidas a un análisis de clasificación, utilizando la distancia euclidiana y agrupadas por el método de promedios pares no ponderados (UPGMA), posteriormente fueron ordenadas en un escalamiento no métrico multidimensional (NMDS). Finalmente, para determinar el grado de contribución de las distintas especies que caracterizaron los sectores de muestreo, se utilizó el análisis de similitud porcentual (SIMPER). Estos análisis se realizaron a través del programa PRIMER-E v6 (Clarke y Gorley, 2005). Una vez identificadas las asociaciones se realizó un análisis inverso de Kaandorp (1986) para establecer la composición de las agrupaciones y determinar especies exclusivas (concentran el 100 % de su abundancia en un ensamblaje), especies características (concentran al menos el 65 % de su abundancia en un grupo y tiene una frecuencia mayor al 65 %) y especies generalistas (aparecen en todos los sectores o en la gran mayoría de ellos).
RESULTADOS
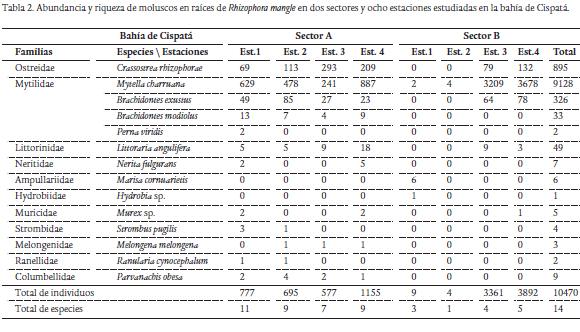
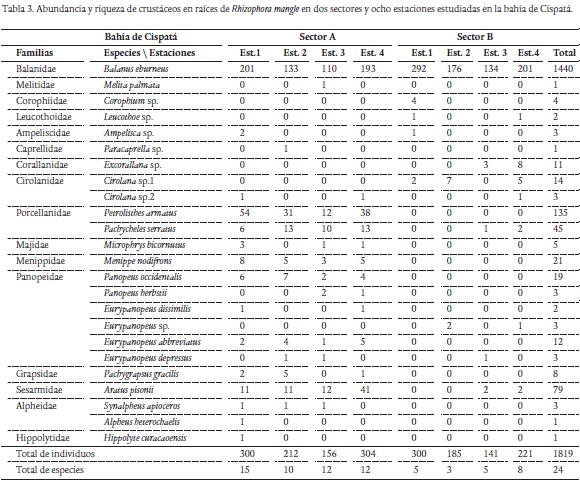
De los 12289 individuos recolectados en los cuatro muestreos, 10470 pertenecieron al phylum Mollusca (85,2 %) y los restantes 1819 al subphylum Crustacea (14,8 %) (Tablas 2 y 3). De moluscos se identificaron 14 especies distribuidas en 11 familias y dos clases; Bivalvia y Gastropoda. La clase Bivalvia representó el 99 % del total de moluscos recolectados en las raíces, siendo Mytella charruana (9128 ind.) y Crassostrea rhizophorae (895 ind.) las especies más abundantes. La clase Gastropoda reportó el 1 % del total de moluscos recolectados, siendo Laguncularia racemosa la dominante con 49 individuos (Tabla 2). De crustáceos se identificaron 24 especies distribuidas en 16 familias y cuatro órdenes; Sessillia, Decapoda, Isopoda y Amphipoda. El orden Sessillia representó el 79,2 % del total de crustáceos registrados en las raíces, siendo Balanus eburneus la especie más abundante con un total de 1440 individuos. Para el orden Decapoda se registraron 15 especies y ocho familias, equivalente al 18,7 % del total de crustáceos recolectados, siendo Petrolisthes armatus (135 ind.), Aratus pisonii (79 ind.), Pachycheles serratus (45 ind.) y Menippe nodifrons (21 ind.) las especies más abundantes (Tabla 3). El orden Isopoda estuvo representado por tres especies y dos familias, constituyendo el 1,5 % del total de crustáceos registrados, de está Cirolana sp.1 (14 ind.) y Excorallana sp. (11 ind.) fueron las especies más abundantes. En el orden Amphipoda, se identificó un total de cinco especies con igual número de familias, contribuyendo con el 0,6 % del total de crustáceos recolectados, de estas Corophium sp. (4 ind.), Ampelisca sp. (3 ind.) fueron las especies más abundantes durante el estudio (Tabla 3).
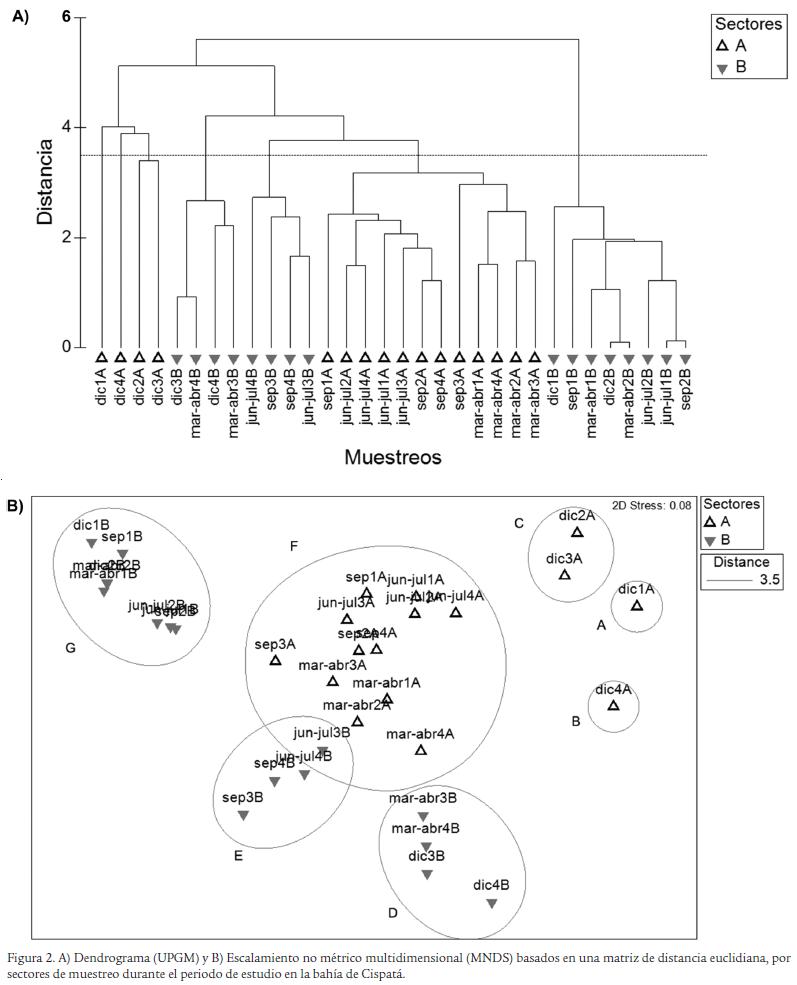
La conformación de los grupos se efectuó con un corte en la escala de distancia euclidiana de 3,5, según el método de promedios pares no ponderados (UPGMA) (Fig. 2A), con base en lo cual se conformaron siete grupos bien diferenciados en el escalamiento no métrico multidimensional (NMDS) (Fig. 2B). El grupo A, formado por una estación, se registraron los decápodos Alpheus heterochaelis y Hippolyte curacaoensis como especies exclusivas. El grupo B, igualmente formado por una estación, lo conforman el anfípodo caprélido Paracaprella sp. y el mejillón verde Perna viridis como especies exclusivas. En el grupo C, que reunió dos estaciones, lo constituye Melongena melongena como una especie característica, mientras que el anfípodo Melita palmata y el cangrejo Panopeus herbstii se presentaron como especies exclusivas. En el grupo D y E, no se registraron especies características y exclusivas, al no presentar un patrón de abundancia que explique la formación de la asociaciones del dendrograma. El grupo F, que reunió a doce de las estaciones, se destacó por presentar a los decápodos Menippe nodifrons, P. occidentalis y Synalpheus apioceros como especies características. El grupo G, que asoció a ocho estaciones, se caracterizó por la presencia de los moluscos Marisa cornuarietis y Hydrobia sp. y el anfípodo Corophium sp. como especies exclusivas para este sector que recibe la influencia de aguas continentales, lo que confirma que estas especies están adaptadas a bajas concentraciones de salinidad (Tabla 4).
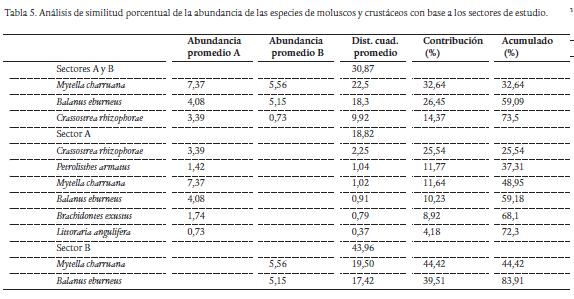
El análisis de similitud porcentual (SIMPER) registró en los dos sectores de muestreo un promedio de distancia euclidiana de 30,9, en la que M. charruana, B. eburneus y C. rhizophorae mostraron el acumulado más alto de similitud (73,5 %), indicando que fueron las especies más importantes en términos de abundancia en casi todas las estaciones de la bahía de Cispatá. El promedio bajo para la distancia (18,8) en el sector A, se fundamenta en la asociación de estas tres especies con P. armatus, L. angulifera y B. exustus, mientras que el alto promedio de distancia (44) en el sector B, indica que M. charruana y B. eburneus fueron las especies dominantes en todas las estaciones de dicho sector (Tabla 5).
DISCUSIÓN
El número de especies (38 en total, 14 especies de moluscos y 24 de crustáceos) en las raíces de R. mangle en la bahía de Cispatá fue menor al reportado por Reyes y Campos (1992b) quienes identificaron 80 especies colonizadoras en la bahía de Chengue, de las cuales 15 especies fueron moluscos y 35 crustáceos, coincidiendo en diez especies con este estudio. No obstante, el número total de especies, tanto de moluscos como de crustáceos, fue mayor al compararlo con el estudio de Quinceno y Palacio (2008), donde recolectaron en la ciénaga La Boquilla (Sucre), un total 33 especies, cinco moluscos y 16 crustáceos. Así como también el realizado por García y Palacio (2008), quienes caracterizaron los macroinvertebrados asociados a las raíces del mangle rojo en dos bahías del golfo de Urabá, identificando 11 moluscos y 12 crustáceos en la bahía Turbo, y 11 moluscos y 14 crustáceos en la bahía El Uno. Los valores registrados en el presente estudio y comparados con otras áreas del Caribe colombiano, pueden deberse a las características de la bahía en particular, la cual presenta varios ambientes que favorecen la existencia de una heterogeneidad de nichos e.g. pastos marinos y sustratos fangosos y arenofangosos.
De las 14 especies de moluscos identificadas, los bivalvos fueron la clase dominante, siendo Mytella charruana la especie abundante y constante en los dos sectores durante el periodo de estudio, por lo que puede considerarse de gran importancia bioecológica y en algunos casos económica (Freitas et al., 2012). En inventarios exhaustivos de la malacofauna del Caribe colombiano (Díaz y Puyana, 1994; Echeverría et al., 2007; Meza et al., 2007), no se había registrado la presencia de M. charruana en la región. Los hallazgos de autores como Gillis et al. (2009) y Puyana et al. (2012) parecen sugerir que M. charruana fue naturalizada en la bahía de Cartagena antes de 2008, ampliando el ámbito de distribución original de la especie para el Caribe colombiano. Este es el primer registro de la especie en Córdoba como un componente dominante de la comunidad de organismos sésiles asociado a las raíces sumergidas de R. mangle.
Otros bivalvos representativos en las raíces R. mangle fueron las especies C. rhizophorae y B. exustus, las cuales se registraron en los sectores de muestreo. Según Reyes y Campos (1992a) y Romero y Polanía (2008), señalan que estas especies son un componente importante de los ecosistemas estuarinos, indicando que su presencia en aguas salobres da una idea de la amplia tolerancia de estas a factores abióticos. Márquez y Jiménez (2002) en su estudio en el golfo de Santa Fe, Venezuela, catalogan a C. rhizophorae y B. exustus como especies de amplia tolerancia a las fluctuaciones de salinidad, considerándolas como eurihalinas. Prüsmann y Palacio (2008), destacan la presencia de B. exustus, como la más abundante en las raíces de mangle rojo en una laguna costera del golfo de Morrosquillo.
Los gastrópodos Nerita fulgurans, Murex sp. y L. angulifera, se presentaron ocasionalmente con abundancias bajas en el estudio. Según Romero y Polanía (2008) estas especies se caracterizan entre otros aspectos por adaptarse a sustratos duros para su adhesión. N. fulgurans prefiere adherirse a la sección sumergida de las raíces de R. mangle, mientras que L. angulifera lo hace por encima del nivel del agua. La presencia de otros gasterópodos reportados, como es el caso de Hydrobia sp. y M. cornuarietis, se basa de acuerdo con Cosel (1986) en la tolerancia a rangos amplios de salinidad por periodos prolongados, así como altas concentraciones de materia orgánica que acumulan las raíces del mangle rojo.
De las 24 especies de crustáceos identificados, los cirripedios fueron el grupo dominante del total de crustáceos registrados en las raíces, siendo B. eburneus, la especie que siempre se recolectó en las raíces. Este cirripedio se encontró formando grandes agrupaciones, compitiendo por espacio y alimento con los mejillones B. exustus y M. charruana. Autores como Romero y Polanía (2008) y Prüsmann y Palacio (2008) señalan que los cirripedios crecen mejor en aguas turbias y tranquilas, debido a que se alimentan de material en suspensión; características que se ajustan con las estaciones de muestreo ubicadas en la bahía de Cispatá. Además, estos resultados confirman lo registrado por Cedeño et al. (2010) quienes reportaron a B. eburneus como una de las especie de mayor dominancia en la laguna de Bocaripo, Venezuela. Por otro lado, Reyes (1991), menciona que esta especie es común en ambientes con cierto grado de contaminación, por lo que es capaz de tolerar y desarrollarse en dichas áreas.
Dentro de los crustáceos decápodos, A. pisonii y P. armatus, presentaron la mayor abundancia en el sector A. Morao (1983), encontró estas especies en la laguna de la Restinga, indicando que su presencia en aguas salobres proporciona una idea de la amplia tolerancia a la salinidad. Márquez et al. (2006) en su estudio en el golfo de Santa Fe señalan a P. armatus como una especie eurihalina. Lalana et al. (1985) y Lacerda et al. (2001), hacen mención que estos crustáceos decápodos son organismos que se alimentan principalmente de detritos. Sin embargo A. pisonii también es catalogada como una especie de hábitos herbívoros, alimentándose de las hojas frescas de R. mangle (Prahl y Sánchez, 1985). Corophium sp. y Ampelisca sp., son especies eurihalinas y características de ambientes de bosque de manglar (Ellison y Farnsworth, 1990). En el presente estudio estas especies se encontraron entre los densos agregados de ostras y mejillones, principalmente en el sector con mayor influencia de agua dulce. Según Barnard y Agard (1986) y Martín et al. (2002), estos anfípodos habitan en fondos someros, en donde por lo general se desplazan libremente sobre algas o cualquier otro tipo de sustrato vivo que puedan encontrar. Excorallana sp. y Cirolana sp.1, también se encontraron en las estaciones con el mayor aporte de aguas continentales, como aquellas provenientes del río Sinú. Según Brusca et al. (2005), estas especies son capaces de tolerar rangos amplios de salinidad, por lo que son frecuentes encontrarlas también en sistemas estuarinos y algunas veces en lagos. Ellison y Farnsworth (1990) mencionan que especies como las descritas contribuyen con el proceso de degradación de los árboles de mangle al perforarlos para la construcción de sus madrigueras.
La distribución de moluscos y crustáceos en las raíces de R. mangle en la bahía de Cispatá, estuvo determinada por un gradiente espacio temporal, como resultado de las diferencias en composición y abundancia de especies. Romero y Polanía (2008) mencionan que en los ecosistemas estuarinos inciden factores ambientales como las concentraciones de salinidad y partículas disueltas, que son regulados por las temporadas de sequía y lluvias, condicionado un patrón de distribución y abundancia de las especies en los manglares. Reyes y Campos (1992a) señalan que la fauna epibionte varía como consecuencia de los hábitos de alimentación, migraciones y depredación, así como la interacción con otras especies provenientes de ecosistemas vecinos. Los mayores valores de riqueza tanto de moluscos como de crustáceos en el sector A, podría estar asociado a una mayor complejidad de hábitat, ofreciendo más espacios potencialmente disponibles, importantes para el establecimiento de epibiontes (Kovalenko et al., 2012). En los sectores A y B tres especies registraron las más altas abundancias; M. charruana (9128 ind.), B. eburneus (1440 ind.) y C. rhizophorae (895 ind.). Poza y Rodríguez (1987) y Díaz y Puyana (1994) señalan que estas especies se desarrollan en ambientes estables. No obstante, también presentan una gran capacidad de adaptación y ajuste a las variaciones hidroclimáticas, mostrándose como especies fuertemente eurihalinas; lo que también se reflejó en la alta contribución de similitud (> 80%) de M. charruana y B. eburneus en el sector B. La presencia del porcelánido P. armatus (135 ind.) y Aratus pisonii (79 ind.) indica que son especies de bosques de manglar con gran movilidad y mecanismos de adaptación fisiológicos. William (1984) señala que estas especies son abundantes en las raíces sumergidas de R. mangle, ya que se encuentra entre las colonias de mitílidos y dentro del tejido de las esponjas, siendo corroborado este comportamiento, exclusivamente en todas las estaciones del sector A.
AGRADECIMIENTOS
Los autores agradecemos a la Universidad de Córdoba por los laboratorios, equipos y todo su apoyo para la realización de este trabajo; así como a la Facultad de Ciencias Básicas y el Departamento de Biología por el apoyo logístico brindado durante la investigación.
BIBLIOGRAFÍA
Abbott R. American Seashells: The Marine Mollusca of the Atlantic and Pacific Coasts of North America. New York: Van Nostrand Reinhold Company; 1974. p. 663. [ Links ]
Báez D. Estructura de la biocenosis asociada a raíces sumergidas de mangle rojo (Rhizophora mangle) en la isla Tintipán, archipiélago de San Bernardo, Caribe colombiano. [trabajo de grado. Bogotá: Facultad de Ciencias, Pontificia Universidad Javeriana; 2001. [ Links ]
Barnard J. The Families and Genera of Marine Gammaridean Amphipoda. Washington D.C.: Smithsonian Institution Press; 1969. p. 271. [ Links ]
Barnard J, Agard J. A new species of Ampelisca (Crustacea: Amphipoda) from Trinidad. Bull Mar Sci. 1986;39(3): 630-636. [ Links ]
Brusca R., Findley L, Hastings P, Hendrickx M, Torre J, Van Der Heiden A. Macrofaunal diversity in the Gulf of California. En: Cartron JL, Ceballos G, Felger R. editors. Biodiversity, ecosystems and conservation in northern Mexico. Oxford: Oxford University Press; 2005. p. 179-202. [ Links ]
Carballo J. Distribución de Ecteinascidia turbinata (Ascidiacea: Perophoridae) en los manglares de la Península de Yucatán, México. Rev Biol Trop. 2000;48 (2-3):365-369. [ Links ]
Castaño A, Urrego L, Bernal G. Dinámica del manglar en el complejo lagunar de Cispatá (Caribe colombiano) en los últimos 900 años. Rev Biol Trop. 2010;58(4):1347-1366. [ Links ]
Cedeño J, Jiménez M, Pereda L, Allen T. Abundancia y riqueza de moluscos y crustáceos asociados a las raíces sumergidas del mangle rojo (Rhizophora mangle) en la laguna de Bocaripo, Sucre, Venezuela. Rev Biol Trop. 2010;58 (suppl.3):213-226. [ Links ]
Clarke K, Gorley R. PRIMER v.6: User Manual / Tutorial. PRIMER-E Ltda. UK: Plymouth; 2005. p. 192. [ Links ]
Cosel R. Moluscos de la región de la ciénaga Grande de Santa Marta (costa del Caribe colombiano). An Inst Inv Mar Punta de Betín. 1986;15-16:79-370. [ Links ]
Díaz J, Puyana M. Moluscos del Caribe colombiano. Un catálogo ilustrado. Bogotá: COLCIENCIAS, Fundación Natura e INVEMAR; 1994. p. 291. [ Links ]
Dittmann S. Abundance and distribution of small infauna in mangroves of Missionary Bay, North Queensland, Australia. Rev Biol Trop. 2001;49(2):535-544. [ Links ]
Echeverría F, Aguirre N, Castaño J, Valderrama A, Peña J, Giudice C. Caracterización fisicoquímica y biológica de la bahía de Cartagena en la zona de Mamonal para la evaluación de pinturas antiincrustantes en condiciones estáticas. Rev Fac Ing Univ Antioquia. 2007;39:7-20. [ Links ]
Ellison A, Farnsworth E. The ecology of Belizean mangrove-root fouling communities. I. Epibenthic Fauna is barriers to isopod attack of mangrove roots. J Exp Mar Biol Ecol. 1990;142(1):91-104. [ Links ]
Farnsworth E, Ellison A. Scale-dependent spatial and temporal variability in biogeography of mangrove root epibiont communities. Ecol Monogr. 1996;66(1):45-66. [ Links ]
Freitas S, Pamplin P, Legat J, Fogaca F, Barros R. Conhecimento tradicional das marisqueiras de Barra Grande, área de proteção ambiental do delta do Rio Parnaíba, Piauí, Brasil. Ambient Soc. 2012;15(2):91-112. [ Links ]
García JM, Palacio J. Macroinvertebrados asociados a las raíces sumergidas del mangle roja (Rhizophora mangle) en las Bahías Turbo y El Uno, golfo de Urabá, Caribe colombiano. Gestión y Ambiente. 2008;11(3):55-66. [ Links ]
Gillis N, Walters L, Fernandes F, Hoffman E. Higher genetic diversity in introduced than in native populations of the mussel Mytella charruana: evidence of population admixture at introduction sites. Diversity Distrib. 2009;15(5):784-795. [ Links ]
Granadillo L, Urosa L. La familia Balanidae (Cirripedia, Thoracica) en el oriente de Venezuela. Bol Inst Oceanogr Venezuela. 1984;23(1-2):15-41. [ Links ]
Kaandorp J. Rocky substrate communities of the infralitoral fringe of the boundaries coast, NW France: A quantitative survey. Mar Biol. 1986;92:22-256. [ Links ]
Kensley B, Schotte M. Guide to the marine isopod crustaceans of the Caribbean. Washington, D.C. and London; Smithsonian Institution Press; 1989. p. 308. [ Links ]
Kovalenko K, Thomaz S, Warfe D. Habitat complexity: approaches and future directions. Editorial Review. Hydrobiologia. 2012;685(1):1-17. [ Links ]
Lacerda L, Conde J, Kjerfve B, Álvarez-León R, Alarcón C, Polanía J. American Mangroves. En: Lacerda LD. editor. Mangrove Ecosystem: Function and Management. Berlín: Springer-Verlag; 2001. p. 1-62. [ Links ]
Lalana R, Álvarez M, Ortíz M, Pérez M, Veledo T. Organismos asociados a las raíces de mangle, Rhizophora mangle, en lagunas costeras y de cayos de Cuba. Rev Invest Mar. 1985;VI(2-3):59-71. [ Links ]
Lodeiros C, Marín B, Prieto A. Catálogo de moluscos marinos de las costas nororientales de Venezuela: Clase Bivalvia. Caracas: Ediciones Apudons; 1999. p. 109. [ Links ]
Márquez B, Jiménez M. Moluscos asociados a las raíces sumergidas del mangle rojo Rhizophora mangle, en el golfo de Santa Fe, Estado Sucre, Venezuela. Rev. Biol. Trop. 2002;50(3):1101-1112. [ Links ]
Márquez B, Blanco J, Jiménez M, Allen T. Crustáceos asociados a las raíces del mangle rojo Rhizophora mangle (L.) en el golfo de Santa Fe, Estado Sucre, Venezuela. Ciencia. 2006;14 (1):12-27. [ Links ]
Martín A, Ortiz M, Díaz Y. Nuevos registros de anfípodos (Crustacea: Peracarida) para Venezuela. Bol Invest Mar Cost. 2002;31(1):15-24. [ Links ]
Meza M, Calixto D, Velosa L, Acosta E, Puyana M, Morales A, Panqueva J. Valoración de la efectividad antiincrustante de recubrimientos aplicados a embarcaciones que operan en la bahía de Cartagena. Ciencia y Tecnología de Buques. 2007;1(1):17-26. [ Links ]
Morao A. Diversidad y fauna de rnoluscos y crustáceos asociados a las raíces sumergidas del mangle rojo, Rhizophora mangle en la Laguna de la Restinga. [trabajo de grado. Cumaná: Biología Marina, Núcleo Sucre, Universidad de Oriente; 1983. [ Links ]
Ortiz M. Clave gráfica para la identificación de familias y géneros de anfípodos del suborden Gammaridea del Atlántico Occidental Tropical. An Inst Invest Mar Punta Betín. 1994;23(1):59-101. [ Links ]
Patiño F, Flórez F. Estudio ecológico del golfo de Morrosquillo. Bogotá: Universidad Nacional, Fondo FEN; 1993. p. 109. [ Links ]
Pérez M. Caracterización del ecosistema de manglar en el golfo de Morrosquillo (San Onofre, Sucre) con el propósito de propiciar su conservación. [trabajo de grado. Medellín: Universidad Nacional de Colombia; 1998. [ Links ]
Poza B, Rodríguez R. Supervivencia de la ostra de mangle Crassostrea rhizophorae (Gilding, 1828) a las variaciones de temperatura, salinidad y pH. Soc Cienc Nat La Salle. 1987;18(127-128):217-231. [ Links ]
Prahl H, Sanchéz O. Cangrejos grápsidos (Crustacea: Decapoda: Grapsidae) del Pacífico colombiano. Boletín Ecotrópica. 1985;14(1):21-33. [ Links ]
Prüsmann J, Palacio J. Colonización de moluscos y crustáceos en raíces de mangle rojo en una laguna costera de la punta norte del golfo de Morrosquillo. Gestión y Ambiente. 2008;11(3):77-86. [ Links ]
Puyana M, Prato J, Díaz JM. Mytella charruana (D'Orbigny) (Mollusca: Bivalvia: Mytilidae) en la bahía de Cartagena, Colombia. Bol Invest Mar Cost. 2012;41(1):213-217. [ Links ]
Quiceno P, Palacio J. Aporte al conocimiento de los macroinvertebrados asociados a las raíces del mangle (Rhizophora mangle) en la cienaga La Boquilla, municipio de San Onofre, sucre. Gestión y Ambiente. 2008;11(3):67-78. [ Links ]
Rathbun M. The cancroid crabs of America of the families Euryalidae, Portunidae, Atelecyclidae, Cancridae and Xanthidae. Washington D.C.: Smithsonian Institution Press; 1930. p. 609. [ Links ]
Reyes R. Macroinvertebrados asociados a las raíces de Rhizophora mangle Linnaeus, 1753 (mangle rojo), en la ciénaga Grande de Santa Marta y en la bahía de Chengue y Nenguange, Caribe colombiano. [trabajo de maestría. Bogotá: Universidad Nacional de Colombia; 1991. [ Links ] Reyes R, Campos N. Moluscos, anélidos y crustáceos asociados a las raíces de Rhizophora mangle Linnaeus, en la región de Santa Marta, Caribe colombiano. Caldasia. 1992a;17(1):133-148. [ Links ]
Reyes R, Campos N. Macroinvertebrados colonizadores de raíces de Rhizophora mangle en la bahía de Chengue, Caribe colombiano. An Inst Inv Mar Punta Betín. 1992b; 21(1):101-116. [ Links ]
Romero P, Polanía J. Sucesión temprana de la taxocenosis Mollusca-Annelida-Crustacea en raíces sumergidas de mangle rojo en San Andrés isla, Caribe colombiano. Revista de Biología Marina y Oceanografía. 2008;43(1): 63-74. [ Links ]
Ruíz M, Bernal G, Polanía J. Influencia del río Sinú y el mar Caribe en el sistema lagunar de Cispatá. Bol Invest Mar Cost. 2008;37(1):29-49. [ Links ]
Warmke M, Abbott R. Caribbean seashells. A guide to the Marine mollusca of Puerto Rico and other West Indian Islands. Bermuda and the Lower Florida Keys; 1975. p. 165. [ Links ]
Williams A. Shrimps, Lobsters, and Crabs of the Atlantic Coast of the Eastern United State, Maine to Florida. Washington: Smithsonian Ins. Press; 1984. p. 550. [ Links ]
