










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.1 Bogotá Jan./Apr. 2015
https://doi.org/10.15446/abc.v20n1.41099
Artículo de investigación
USO DOS RECURSOS POR DUAS ESPÉCIES SIMPÁTRICAS DE Ameivula (Squamata: Teiidae) EM UM ECÓTONO DE MATA ATLÂNTICA-CAATINGA
Use of Resources by Two Sympatric Species of Ameivula (Squamata: Teiidae) in an Atlantic Forest-Caatinga Ecotone
Uso de recursos por dos especies simpátricas de Ameivula (Squamata: Teiidae) en un ecotono de bosque Atlántico-Caatinga
Régis Vinícius Souza SANTOS1; Crizanto Brito DE-CARVALHO2; Evellyn Borges de FREITAS2; Fernanda Barros GUEIROS2; Renato Gomes FARIA2.
1 Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil.
2 Laboratório de Herpetologia, Departamento de Biologia, Centro de Ciências Biológicas e da Saúde, Universidade Federal de Sergipe, São Cristóvão, SE, Brasil.
For correspondence: regisvinicius@gmail.com
Received 6th December 2013, Returned 6th April for revisión 2014, accepted 5th May 2014.
Citation / Citar este artículo como: Santos RVS, De-Carvalho CB, Freitas EB, Gueiros FB, Faria RG. Uso dos recursos por duas espécies simpátricas de Ameivula (Squamata: Teiidae) em um ecótono de Mata Atlântica-Caatinga. Acta biol. Colomb. 2015;20(1):67-77. doi: http://dx.doi.org/10.15446/abc.v20n1.41099.
RESUMO
As formas de utilização dos recursos espaciais e temporais por duas espécies simpátricas de Ameivula foram estudadas em um ecótono de Mata Atlântica e Caatinga, situado no Parque Nacional Serra de Itabaiana, Sergipe, Brasil. O estudo foi desenvolvido entre fevereiro de 2008 e janeiro de 2009. O método adotado foi o de busca ativa limitado por tempo, sendo utilizado o período das 06:00 às 18:00 h. As duas espécies foram verificadas ao longo de todos os meses de coleta, sendo A. ocellifera a com maior número de registros. Apesar da alta sobreposição de nicho temporal observada (Ojk = 0,90), A. ocellifera e A. abaetensis diferiram em relação aos horários de maior atividade. A. ocellifera foi avistado com maiores frequências (≥10 %) entre 08:00 e 13:00 h, enquanto que A. abaetensis, entre as 09:00 às 13:00 h. As duas espécies foram mais frequentes em dias ensolarados, com padrões similares de atividade no momento do avistamento, tendo as mesmas sido verificadas, em geral, andando sobre areia (A. ocellifera) ou folhas próximas à base de arbustos (A. abaetensis). Os comportamentos adotados pelas duas espécies de Ameivula em resposta à aproximação do observador foram diferenciados e estiveram relacionados ao grau de exposição e, consequentemente, de vulnerabilidade à predação. A coexistência de A. ocellifera e A. abaetensis na área de estudo está possivelmente relacionada a pequenas divergências no padrão de uso de recursos temporais e espaciais pelas duas espécies, sendo provavelmente essas diferenças resultantes de interações ecológicas atuais. Os dados obtidos neste estudo contribuem para o conhecimento sobre a história natural e ecologia das espécies de Ameivula e, principalmente, geram informações úteis para a conservação destas espécies em hábitats de transição Mata Atlântica-Caatinga.
Palavras-chave: Ameivula abaetensis, Ameivula ocellifera, coexistência, ecologia, lagartos.
ABSTRACT
The forms of spatial and temporal resources use of two sympatric species of Ameivula were studied in an Atlantic Forest-Caatinga ecotone located in Parque Nacional Serra de Itabaiana, Sergipe, Brazil. The study was conducted between February 2008 and January 2009. The method used was an active search for a limited time, from 06:00 am to 06:00 pm. The species were observed during all months sampled, and A. ocellifera presented the highest number of sightings. Despite the high temporal niche overlap observed (Ojk = 0.90), A. ocellifera and A. abaetensis differed regarding the times of higher activity. A. ocellifera was most seen (≥ 10%) between 08:00 am and 01:00 pm, and A. abaetensis from 09:00 am to 01:00 pm. Both species were found more frequently on sunny days, with similar patterns of activity at the time of sighting, being usually observed walking on sand (A. ocellifera) or leaves near the base of shrubs (A. abaetensis). The behaviors demonstrated by both species of Ameivula in response to the approach of the observer were different and were related to the level of exposure along with the vulnerability to predation. The coexistence of A. ocellifera and A. abaetensis in the study area is possibly related to minor differences in the use pattern of temporal and spatial resources by both species, and these differences are probably due to the current ecological interactions. The results obtained in this study contribute to the understanding of the natural history and ecology of the species of Ameivula and mainly generate useful information for the conservation of these species in habitats transitional of Atlantic Forest-Caatinga.
Keywords: Ameivula abaetensis, Ameivula ocellifera, coexistence, ecology, lizards.
RESUMEN
En un ecotono de bosque atlántico y Caatinga, ubicado en el Parque Nacional Serra de Itabaiana, Sergipe, Brasil, fueron estudiadas las formas de uso de recursos espaciales y temporales de dos espécies simpátricas de Ameivula El estudio se realizó entre febrero de 2008 y enero de 2009. El método adoptado fue la búsqueda activa con control de tiempo, en el periodo de las 06:00-18:00 horas. Se observó que la especie con más registros fue A. ocellifera. A pesar del alto solapamiento de nicho temporal observado (Ojk = 0,90), A. ocellifera y A. abaetensis difirieron en relación con los momentos de mayor actividad. A. ocellifera se observó con mayor frecuencia (≥ 10 %) 08:00-13:00, mientras que A. abaetensis de 09:00 a 13:00 horas. Ambas especies fueron más frecuentes en los días soleados, con patrones similares de actividad en el momento de la observación, fueron vistas por lo general caminando sobre la arena (A. ocellifera) o en las hojas cerca de la base de arbustos (A. abaetensis). Conductas adoptadas por las dos especies Ameivula en respuesta al enfoque del observador eran diferentes y estaban relacionados con el grado de exposición de la vulnerabilidad a la depredación. La coexistencia de A. ocellifera y A. abaetensis en el área de estudio se relaciona posiblemente con pequeñas diferencias en el patrón de tiempo de uso de los recursos y el espacio de las dos especies, estas diferencias son probablemente el resultado de las interacciones ecológicas actuales. Los resultados obtenidos en este estudio contribuyen a la comprensión de la historia natural y ecología de las especies de Ameivula y generan principalmente información útil para la conservación de estas especies en hábitats de transición de la Mata Atlántica-Caatinga.
Palabras clave: Ameivula abaetensis, Ameivula ocellifera, coexistencia, ecología, lagartos.
INTRODUÇÃO
A alta similaridade no padrão de uso dos recursos por espécies semelhantes (morfologicamente, filogeneticamente e comportamentalmente) em condição de simpatria, conduz a uma segregação interativa que, de acordo com o Princípio da Exclusão Competitiva (Gause, 1934; Pianka, 2000), é fundamental para prevenir a extinção do competidor mais fraco. Assim, variações na forma de utilização de ao menos uma das três dimensões primárias de recurso (espacial, temporal ou trófica) são esperadas entre espécies que coexistem na mesma comunidade ecológica (Pianka, 1973; Schoener, 1974). A partição de recursos é, portanto, um mecanismo desenvolvido por espécies potencialmente concorrentes para utilizar recursos não aproveitados por membros de outras espécies (Griffin e Silliman, 2011) que leva a um aumento líquido da exploração de recursos pelas comunidades de consumidores e a uma maior biodiversidade (Finke e Snyder, 2008).
No ecótono Mata Atlântica-Caatinga do Parque Nacional Serra de Itabaiana no estado de Sergipe, Brasil, os lagartos teiídeos Ameivula ocellifera (Spix, 1825), espécie com ampla distribuição, e A. abaetensis (Dias, Rocha e Vrcibradic, 2002), incluído recentemente na lista de espécies ameaçadas de extinção (Machado et al., 2008), coexistem na área do Parque, evento incomum em toda a região tropical do continente americano (Vitt et al., 2000). No Brasil, até o presente momento, o único registro de coexistência destas espécies foi para a restinga do Abaeté na Bahia (Dias e Rocha, 2007), não existindo, contudo, informações sobre como elas diferem em características ecológicas enquanto simpátricas em habitats de transição.
Este trabalho teve como objetivo caracterizar as formas de utilização dos recursos espaciais e temporais das espécies simpátricas Ameivula ocellifera e A. abaetensis em um ecótono Mata Atlântica-Caatinga, e verificar a existência de variações nos padrões de uso dos recursos que contribuem para a coexistência dessas espécies na área.
MATERIAL E MÉTODOS
Área de estudo
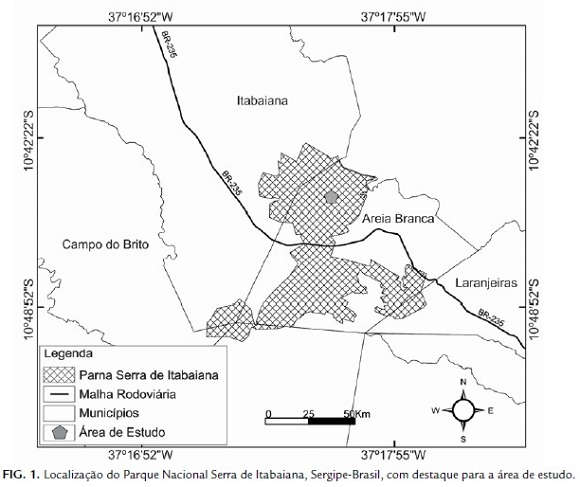
Ameivula ocellifera e A. abaetensis foram estudados no Parque Nacional Serra de Itabaiana–PNSI (10°40' S, 37°25' O, 7966 ha, 200-670 m s.n.m., Fig. 1), localizado no Estado de Sergipe entre os municípios de Areia Branca, Itabaiana, Laranjeiras, Itaporanga D'ajuda e Campo do Brito, abrangendo as Serras de Itabaiana, Comprida e do Cajueiro (Brasil, 2005; Carvalho e Vilar, 2005; Dantas e Ribeiro, 2010). O clima regional é do tipo As, de acordo com a classificação de Köppen (Kottek et al., 2006), com excedente hídrico de inverno moderado. A vegetação é caracterizada por diferentes fisionomias marcadas por fragmentos de Mata Atlântica e de Caatinga (Ab'saber, 1967; Vicente, 1997; Carvalho e Vilar, 2005). Entre esses fragmentos é observada uma zona de transição ecológica composta por áreas fechadas, com vegetação arbórea secundária, localizadas principalmente nas encostas acompanhando os riachos que recortam as serras; e áreas abertas, que também ocorrem nas encostas e nas partes mais altas, com uma vegetação rasteira diversificada, arbustos e arvoretas, que se desenvolvem em solos de areias brancas (Vicente et al., 1997).
Métodos de coleta e análise de dados
Os dados foram coletados quinzenalmente em campanhas de dois dias consecutivos, entre fevereiro de 2008 e janeiro de 2009, numa área de transição Mata Atlântica-Caatinga de 2,08 ha, situada a 10°44'56 S e 37°20'31 W. O método adotado foi o de busca ativa limitado por tempo, sendo utilizado o período entre as 06:00 e 18:00 h. O esforço amostral efetivo total foi de 576 horas/homem de observação.
Para cada animal encontrado foram anotados os seguintes dados: espécie, data, horário, condição do dia (ensolarado, nublado ou chovendo), posição do lagarto em relação ao sol (sol, sombra ou mosaico de sol e sombra), microhabitats utilizados durante a observação e após a aproximação do observador (microhábitat 1 e 2, respectivamente; areia, gramínea ou serrapilheira) e as atividades desenvolvidas durante esses momentos (atividade 1 e 2, respectivamente; parado, andando, forrageando ou correndo). Para a atividade 1, a categoria "correndo" não foi considerada por se tratar de um comportamento de fuga adotado pelos indivíduos frente aos predadores, não sendo uma atividade comum na ausência desse tipo de estímulo.
Dados de temperatura do ar (°C), precipitação (mm) e umidade relativa do ar (%) foram obtidos na estação agrometeorológica mais próxima da área de estudo (Itabaiana/ SE) e cedidos pelo Centro de Previsão de Tempo e Estudos Climáticos–CPTEC, do Instituto Nacional de Pesquisas Espaciais (INPE). A relação entre as variáveis ambientais e as abundâncias mensais das espécies foi analisada através do teste de correlação de Spearman (Zar, 2010).
Variações entre as duas espécies de Ameivula na utilização do microhábitat, atividade desenvolvida antes e após a aproximação do observador, horário de atividade, condição do dia e exposição do lagarto ao sol foram testadas usando Qui-quadrado (X²) ou teste de G, variando de modo a atender às premissas de cada teste.
As amplitudes de nicho (B) espacial (microhábitat) e temporal (horários de atividade) foram calculadas utilizando oinversodoíndicedediversidadedeSimpson(Simpson,1949).
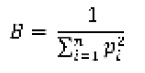
onde p é a proporção numérica da categoria i e n é o número de categorias. A amplitude de nicho B pode variar de 1 (utilização de uma única categoria de microhábitat) a n (utilização igual de todas categorias).
As sobreposições de nicho (Ojk) espacial (microhábitat) e temporal (horários de atividade) foram calculadas através do índice de sobreposição simétrica de Pianka (Pianka, 1973).
Para as análises estatísticas, os dados foram armazenados em planilhas do Excel 2010, e os testes realizados no programa STATISTICA 8.0 (Statsoft, 2008). O nível de significância adotado foi de 5 %.
A nomenclatura científica utilizada neste trabalho segue a proposta da Lista de Répteis da Sociedade Brasileira de Herpetologia (Bérnils e Costa, 2012).
RESULTADOS
Padrões de atividade
Os Ameivula do PNSI foram observados ativos ao longo de todo o ano do trabalho, provavelmente em decorrência das altas temperaturas e da pequena oscilação verificada. A exceção foi o mês de julho quando as duas espécies foram menos avistadas (Fig. 2).
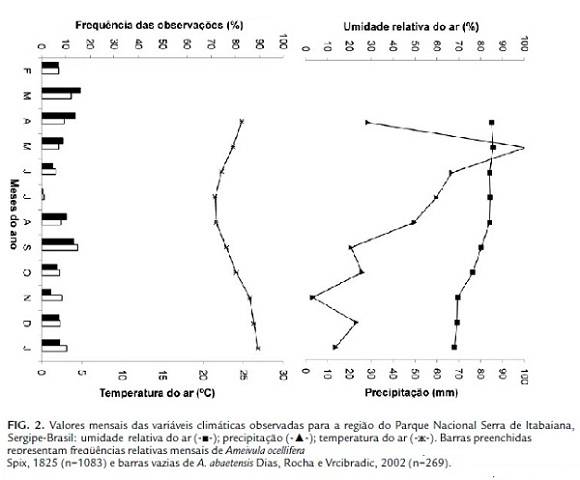
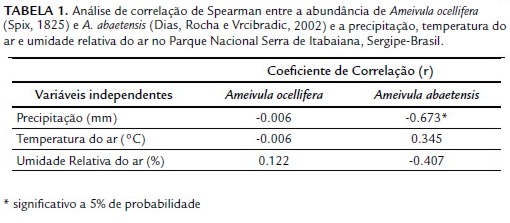
A correlação entre as abundâncias mensais de Ameivula e a precipitação foi negativa para as duas espécies, sendo significativa para A. abaetensis (Spearman, p<0,05) (Tabela 1). Não houve correlação significativa entre as abundâncias e a temperatura do ar e a umidade relativa do ar (Spearman, p>0,05).
Indivíduos de Ameivula ocellifera (A.o.) foram observados sempre em maiores proporções quando comparados a A. abaetensis (A.a.), sendo a razão média A.o./A.a. de 4:1 (Tabela 2).
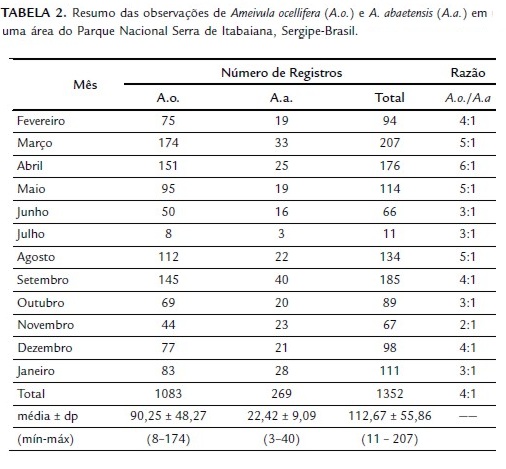
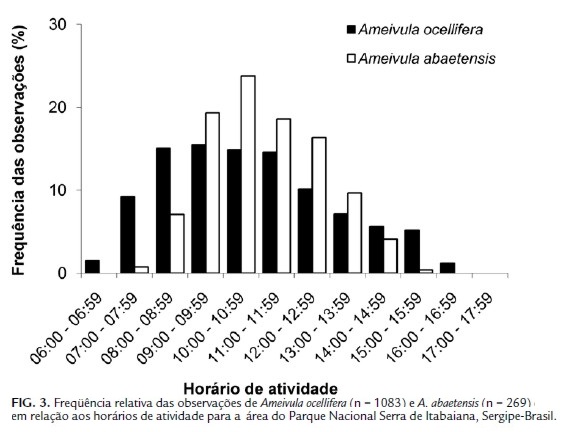
Apesar da alta sobreposição de nicho temporal observada (Ojk = 0,90), Ameivula ocellifera e A. abaetensis diferiram em relação aos horários de maior atividade (G = 96,6759; g.l. = 11; p<0,0001; Fig. 3). Ameivula ocellifera foi observado na área de estudo das 06:00 às 17:00 h, com maiores frequências (≥10 %) entre 08:00 e 13:00 h. Ameivula abaetensis foi avistado das 07:00 às 16:00 h, com maiores picos das 09:00 às 13:00 h. As amplitudes de nicho (B) temporal (horários de atividade) de Ameivula ocellifera e A. abaetensis foram respectivamente 8,32 e 5,84.
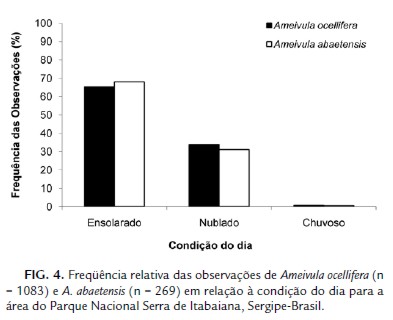
As duas espécies foram mais frequentes em dias ensolarados (G = 0,0864; g.l. = 2; p = 0,9577; BA. o.= 1,85 e BA.a.= 1,78; Ojk= 0,999; Fig. 4). Dos 1143 registros (1083 Ameivula ocellifera e 269 A. abaetensis), 65,83 % dos indivíduos de A. ocellifera e 68,03 % dos A. abaetensis foram encontrados nessa condição.
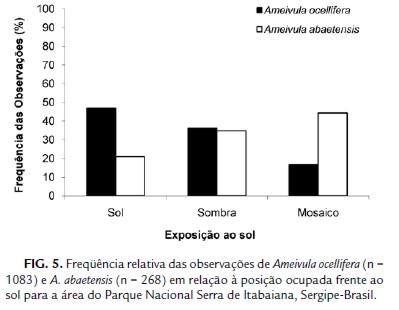
Ameivula ocellifera e A. abaetensis ocuparam diferentes posições frente ao sol (X2 = 107,832; g.l. = 2; p<0,0001), apesar da elevada sobreposição observada (Ojk= 0,81; BA. o. = 2,63 e BA.a.= 2,77). A maioria dos A. ocellifera (47,0%) foi encontrada exposta à luz solar ou na sombra (36,10 %), enquanto que A. abaetensis em áreas de mosaico de sol e sombra (44,40 %) ou sombreadas (34,70 %) (Fig. 5).
Padrões similares de atividade foram observados para as espécies deste estudo no momento da observação (X² = 1,789; g.l. = 2; p = 0,4087), tendo as mesmas sido verificadas, em geral, andando (Ameivula ocellifera – 73,46 % e A. abaetensis – 76,12 %; Fig. 6).
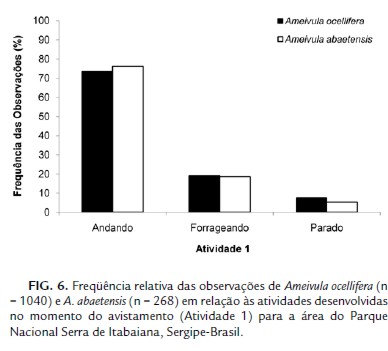
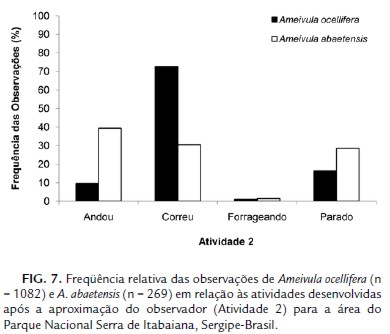
Os comportamentos adotados pelas duas espécies de Ameivula em resposta à aproximação do observador foram diferenciados (G = 182,7229; g.l. = 3; p < 0,0001; Fig. 7). A maior parte dos Ameivula ocellifera (72,74 %) correu à procura de abrigo, enquanto que A. abaetensis adotou principalmente três respostas: corridas (30,48 %), semelhante ao seu congênere ou permaneceu andando (39,41 %) ou parado (28,62 %).
Uso do espaço
No PNSI, as espécies estudadas apresentaram divergências quanto aos microhabitats utilizados no momento da observação (X2 = 479,828; g.l. = 3; p < 0,0001; BA. o.= 2,16 e BA. a.= 1,41; Ojk = 0,41), com Ameivula ocellifera sendo observado em maior proporção sobre areia (62,42 %) e A. abaetensis sobre folhas próximas à base de arbustos (serrapilheira) (82,46 %) (Fig. 8).
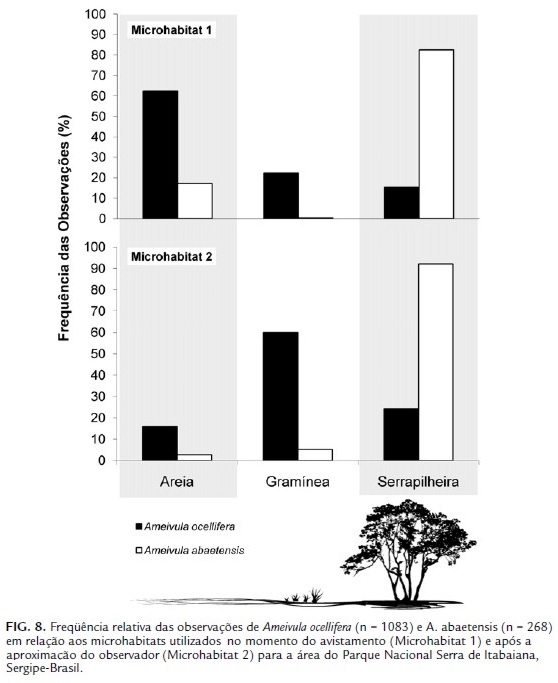
Os Ameivula do PNSI diferiram em relação aos sítios de refúgio adotados (X2 = 423,059; g.l. = 2; p < 0,0001; BA. o. = 2,25 e BA. a.= 1,17; Ojk= 0,42), Ameivula ocellifera procuraram, principalmente, locais com agrupamentos de gramíneas (60,02 %), enquanto que A. abaetensis permaneceram sobre a serrapilheira (92,13 %) (Fig. 8).
DISCUSSÃO
O nordeste brasileiro apresenta temperaturas relativamente elevadas ao longo de todos os meses, ao contrário do que ocorre nas regiões sul, sudeste e centro-oeste do Brasil, onde se observa um regime térmico de estações quentes e frias (Nimer, 1989). Estudos ecológicos nessas localidades têm demonstrado padrões comportamentais caracterizados por grande redução das atividades de diversas espécies de lagartos em épocas do ano nas quais são registradas grandes quedas de temperatura (e.g. Van Sluys, 1992; Zaluar e Rocha, 2000).
Em regiões onde não são registradas grandes diferenças nas temperaturas mensais, essa variável parece não interferir nos padrões de atividade adotados, como observado para os Ameivula do PNSI. Entretanto, um fator, a precipitação, apresentou correlação negativa significativa com a abundância de Ameivula abaetensis e, que pode ter contribuído para a diminuição nas observações das duas espécies no mês de julho. Apesar de neste mês ter sido registrado uma baixa precipitação, entretanto, as atividades de campo coincidiram com dias chuvosos na área (R. V. S. Santos, obs. pess.).
A alta umidade relativa do ar, verificada ao longo do ano, pode ter gerado condições propícias para a manutenção de grandes quantidades de artrópodes, principais itens das dietas dos lagartos (Silva e Araújo, 2008). Pois, em geral, é durante os períodos mais úmidos em que se observam nas áreas tropicais uma maior diversidade e abundância desses invertebrados (Janzen e Schoener, 1968). Deste modo, as atividades das duas espécies de Ameivula ao longo do ano podem ser mantidas em parte pelo grande estoque de presas.
Os padrões de atividade observados para Ameivula ocellifera e A. abaetensis foram semelhantes aos verificados para estas espécies na restinga do Abaeté, Bahia (Dias e Rocha, 2004). O padrão de atividade de um lagarto está fortemente ligado às suas necessidades térmicas, podendo ser influenciado pelas características do microhábitat utilizado (Pianka, 1973). Segundo Adolph (1990), a vegetação associada aos substratos adotados pode interferir na quantidade de luz e, consequentemente, de calor que alcança os lacertílios, levando a alterações na extensão de seu período de atividade. Temperaturas corporais elevadas são essenciais para manter os teiídeos mais ativos e velozes nas fugas e eficientes na perseguição do seu alimento (Freitas e Silva, 2007). Dessa forma, a redução de ambientes favoráveis à termorregulação poderia gerar uma diminuição das atividades desses animais. A influência da estrutura da vegetação no padrão de atividade de lagartos já foi sugerida para Sceloporus occidentalis e S. undulatus por Grover (1996) e para Ameivula ocellifera e A. abaetensis por Dias e Rocha (2007). Sendo assim, é possível que isso também venha ocorrendo com os Ameivula do PNSI, porém novas investigações são necessárias para a sua confirmação.
As duas espécies de Ameivula apresentaram padrão comportamental típico de animais heliófilos, o que é muito comum em representantes desse gênero (Vitt e Breitenbach, 1993). Provavelmente a escolha de locais onde haja maior incidência direta de raios solares seja a condição ótima, ou seja, a que mais se ajusta às necessidades metabólicas de forrageadores ativos (Magnusson et al., 1985). Porém, as elevadas temperaturas em determinados horários do dia, com temperaturas do solo podendo chegar até a 60°C (R. V. S. Santos, obs. pess.), podem ter levado ao grande número de registros das duas espécies de Ameivula ocupando regiões mais sombreadas, como forma de evitar o superaquecimento.
Padrões similares de atividade observados para as espécies deste estudo foram semelhantes ao registrado para grande parte das espécies deste gênero (e.g. Mesquita e Colli, 2003a; Mesquita e Colli, 2003b). Este comportamento é típico de forrageadores ativos, que necessitam de movimentos constantes para a busca de alimento (Pianka e Vitt, 2003).
As reações adotadas pelos indivíduos provavelmente estão relacionadas ao grau de exposição e, consequentemente, de vulnerabilidade à predação. Corridas foram adotadas normalmente pelos indivíduos que se encontravam em microhabitats mais expostos e consequentemente mais vulneráveis a predadores (e.g. areia). Enquanto que o ato de continuar simplesmente andando ou parado, foi mais comum naqueles que já se encontravam próximo ou mesmo em locais de vegetação mais complexa (e.g. gramíneas, folhas na base de arbustos), que oferecem certo grau de proteção. Estudos que fizessem uso das medidas da distância entre os indivíduos e a base dos arbustos mais próximos e as comparassem com os mecanismos de escape adotados por eles, possivelmente dariam uma maior sustentação a essa relação.
Os lagartos podem ocupar diversos hábitats, e dentro destes, microhabitats variados. No entanto, para que uma população consiga persistir ao longo do tempo é necessário que os custos relacionados com as atividades vitais, como forrageio e termorregulação, e com as interações ecológicas, não sejam muito elevados (Pianka e Vitt, 2003; Silva e Araújo, 2008).
Sabe-se que espécies relativamente próximas vivendo em simpatria divergem em pelo menos um dos três eixos de nicho: espacial, temporal ou trófico (Pianka, 1973), sendo a partilha dos recursos espaciais o fator mais importante de separação interespecífica na maioria dos répteis simpátricos (Toft, 1985). Assim, muitas vezes é possível observar em espécies simpátricas de lagartos o uso do espaço de modo diferenciado, o que sugere certa preferência, seleção ou especialização por hábitats ou microhabitats particulares, fruto de interações ecológicas atuais ou do passado, para minimizar os efeitos negativos da competição interespecífica (Vitt e Carvalho, 1995; Faria e Araújo, 2004).
Segundo Silva e Araújo (2008), uma diferença nos microhabitats utilizados pode estar associada à disponibilidade de locais igualmente adequados às duas espécies de lagartos ou então à superioridade competitiva de uma delas, que ocuparia os melhores lugares, deixando apenas remanescentes para a outra espécie.
Em áreas abertas como as do presente estudo, a heterogeneidade espacial (disponibilidade de habitats) é menor quando comparada a áreas florestadas, visto que as árvores são escassas e solos sem vegetação são mais comuns. Logo, a presença de microhábitats como os adotados por Ameivula abaetensis, que envolvem uma vegetação mais complexa, composta por áreas com arbustos e serrapilheira acumulada no chão, oferecem uma quantidade maior de recursos, tornando o uso desses locais necessário e adequado para muitas espécies de lagartos (Silva e Araújo, 2008).
A procura por locais com agrupamentos de gramíneas ou serrapilheira no momento da fuga parece ser típico de lagartos forrageadores ativos, como os do gênero Ameivula, que tendem a procurar por áreas mais cobertas e de vegetação um pouco mais densa (Pianka e Vitt, 2003), por serem, em geral, regiões de difícil acesso para predadores visualmente orientados, como aves.
Na área de estudo foram observados também alguns predadores com orientação mais química (e.g. Chironius bicarinatus, Liophis reginae, Liophis viridis, Oxybelis aeneus, Thamnodynastesspp.), sendo que, neste caso, o comportamento de fuga adotado (e.g. corrida) provavelmente tenha sido mais eficiente do que a própria escolha de refúgios, como anteriormente relatado.
CONCLUSÕES
Considerando a filogenia do gênero Ameivula (espécies pertencentes ao antigo gênero Cnemidophorus) (Santos, 2007) e como já afirmado por Carvalho et al., (2005), Ameivula ocellifera e A. abaetensis pertencem à mesma linhagem, a do grupo ocellifera. Deste modo, a partilha de recursos observada entre as duas espécies na área do PNSI, parece estar mais relacionada a interações ecológicas recentes, como a competição (e. g. Losos, 1995; Vitt, 1995; Vitt e Carvalho, 1995), do que a fatores históricos, resultantes de interações prévias responsáveis pela evolução das divergências hoje observadas ("the ghost of competition past" Connell, 1980). Entretanto, para confirmar tal hipótese, seriam necessárias investigações direcionadas, por exemplo, a populações alopátricas das duas espécies que vivem sob condições semelhantes àquelas encontradas na área do Parque, ou experimentos em laboratório que caracterizem tal situação. Por fim, os dados obtidos neste estudo contribuem para o conhecimento sobre a história natural e ecologia das espécies de Ameivula e, principalmente, geram informações úteis para a conservação destas espécies em hábitats de transição Mata Atlântica-Caatinga.
AGRADECIMENTOS
Ao Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) – PARNA Serra de Itabaiana (SE) pela estrutura de apoio aos pesquisadores oferecida durante a realização do trabalho de campo.
REFERÊNCIAS
Ab'saber AN. Domínios morfoclimáticos e províncias fitogeográficas do Brasil. Orientação. 1967;3:45-48. [ Links ]
Adolph SC. Influence of behavioral thermoregulation on microhabitat use by two Sceloporus lizards. Ecology. 1990;71(1):315-327. Doi: http://dx.doi.org/10.2307/1940271. [ Links ]
Bérnils RS, Costa HC. Répteis brasileiros: lista de espécies. Versão 2012.2. Sociedade Brasileira de Herpetologia. 2012 (Cited 22 Nov 2013). Available in: http://www.sbherpetologia.org.br/. [ Links ]
Brasil. Decreto de 15 de junho de 2005, Ministério do Meio Ambiente, criação do Parque Nacional Serra de Itabaiana. Diário Oficial da União. 2005;1(114):9-11. Available in: http://pesquisa.in.gov.br/imprensa/jsp/visualiza/index.jsp?data=16/06/2005&jornal=1&pagina=9&totalArquivos=72. [ Links ]
Carvalho CM, Vilar JC. Introdução – Levantamento da biota do Parque Nacional Serra de Itabaiana. In: Carvalho, CM, Vilar JC editors. Parque Nacional Serra de Itabaiana – Levantamento da Biota. Aracaju: IBAMA; 2005. p. 9-14. [ Links ]
Carvalho CM, Vilar JC, Oliveira FF. Répteis e anfíbios. In: Carvalho CM, Vilar JC, editors. Parque Nacional Serra de Itabaiana – Levantamento da Biota. Aracaju: IBAMA; 2005. p. 39-61. [ Links ]
Connell JH. Diversity and the coevolution of competitors, or the ghost of competition past. Oikos. 1980;35(3):131-138. [ Links ]
Dantas TVP, Ribeiro AS. Caracterização da vegetação do Parque Nacional Serra de Itabaiana, Sergipe – Brasil. Biotemas. 2010;23(4):9-18. [ Links ]
Dias EJR, Rocha CFD. Niche differences between two sympatric whiptail lizards (Cnemidophorus abaetensis and Cnemidophorus ocellifer, Teiidae) in the restinga habitat of northeastern Brazil. Braz J Biol. 2007;67(1):41-46. Doi: http://dx.doi.org/10.1590/S1519-69842007000100006. [ Links ]
Dias EJR, Rocha CFD. Thermal ecology, activity patterns, and microhabitat use by two sympatric whiptail lizards (Cnemidophorus abaetensis and Cnemidophorus ocellifer) from northeastern Brazil. J Herpetol. 2004;38(4):586-588. Doi: http://dx.doi.org/10.1670/80-03N. [ Links ]
Faria RG, Araújo AFB. Sintopy of two Tropidurus lizard species (Squamata: Tropiduridae) in a rocky cerrado habitat in central Brazil. Braz J Biol. 2004;64(4):775-786. Doi: http://dx.doi.org/10.1590/S1519-69842004000500007. [ Links ]
Finke DL, Snyder WE. Niche partitioning increases resource exploitation by diverse communities. Science. 2008;321(5895):1488–1490. Doi: 10.1126/science.1160854. [ Links ]
Freitas MA, Silva TFS. Guia ilustrado: A herpetofauna das caatingas e áreas de altitudes do nordeste brasileiro. Pelotas, Rio Grande do Sul: USEB. 2007. p. 384. [ Links ]
Gause GF. The Struggle for existence. Baltimore, MD, U.S.A: The Williams and Wilkins Company; 1934. p.163. [ Links ]
Griffin JN, Silliman BR. Resource Partitioning and Why it Matters. Nature Education Knowledge. 2011;3(10):49. [ Links ]
Grover MC. Microhabitat use and thermal ecology of two narrowly sympatric Sceloporus (Phrynosomatidae) lizards. J Herpetol. 1996;30(2):152-160. [ Links ]
Janzen DH, Schoener TW. Differences in insect abundance and diversity between wetter and drier sites during a tropical dry season. Ecology. 1968;49(1):96-110. [ Links ]
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F. World Map of the Köppen-Geiger climate classification updated. Meteorol Z. 2006;15(3):259-263. [ Links ]
Losos JB. Community evolution in greater Antillean Anolis lizards: Phylogenetic patterns and experimental tests. Philos. Trans R Soc Lond. 1995;349(1327):69-75. Doi: 10.1098/rstb.1995.0092. [ Links ]
Machado AMB, DRummond GM, Paglia AP. Livro vermelho da fauna brasileira ameaçada de extinção. 1 ed. Brasília: Fundação Biodiversitas; 2008. p. 1420. [ Links ]
Magnusson WE, Paiva LJ, Rocha RM, Franke CR, Kasper LA, Lima AP. The correlates of foraging mode in Brazilian lizards. Herpetologica. 1985;41:324-332. [ Links ]
Mesquita DO, Colli GR. Geographical variation in the ecology of populations of some Brazilian species of Cnemidophorus (Squamata, Teiidae). Copeia. 2003a;2003(2):285-298. Doi: http://dx.doi.org/10.1643/0045-8511(2003)003[0285:GVITEO]2.0.CO;2. [ Links ]
Mesquita DO, Colli GR. The ecology of Cnemidophorus ocellifer (Squamata, Teiidae) in a Neotropical savanna. J Herpetol. 2003b;37(3):498-509. Doi: http://dx.doi.org/10.1670/179-02A. [ Links ]
Nimer E. Climatologia do Brasil. 2 ed. Rio de Janeiro: IBGE, Departamento de Recursos Naturais e Estudos Ambientais; 1989. p. 422. [ Links ]
Pianka ER. The structure of lizards communities. Annu Rev Ecol Evol Syst. 1973;4:53-74. Doi: 10.1146/annurev.es.04.110173.000413. [ Links ]
Pianka ER. Evolutionary Ecology. 6th ed. San Francisco, CA: Benjamin Cummings; 2000. p.512. [ Links ]
Pianka ER, Vitt LJ. Lizards: Windows to the evolution of diversity. California: University of California Press; 2003. p. 331. [ Links ]
Santos RML. Estudos evolutivos em espécies de lacertílios brasileiros da família Teiidae (Squamata), com base em dados citogenéticos e moleculares (tese de doutorado). São Paulo: Instituto de Biociências, Universidade de São Paulo; 2007. p. 67. [ Links ]
Schoener TW. Resource partitioning in ecological communities. Science. 1974;185(4145):27–39. [ Links ]
Silva VN, Araújo AFB. Ecologia de lagartos brasileiros. 1 ed. Rio de Janeiro: Technical Books; 2008. p. 271. [ Links ]
Statsoft. Statistica data analysis system version 8.0. Tulsa: Statsoft. 2008. [ Links ]
Toft CA. Resource partitioning in amphibians and reptiles. Copeia. 1985;1985(1):1-21. [ Links ]
Van Sluys M. Aspectos da ecologia do lagarto Tropidurus itambere (Tropiduridae), em uma área do sudeste do Brasil. Braz J Biol. 1992;52(1):181-185. [ Links ]
Vicente A. Levantamento fitossociológico de mata mesófila decídua na Serra de Itabaiana, Sergipe. Publicações Avulsas do Centro Acadêmico Livre de Biologia da Universidade Federal de Sergipe. 1997;1:23-27. [ Links ]
Vicente A, Araújo GMM, Lírio Júnior GP, Santos SC. Descrição parcial e preliminar dos habitats da Serra de Itabaiana, Sergipe. Publicações Avulsas do Centro Acadêmico Livre de Biologia da Universidade Federal de Sergipe. 1997;1:7-21. [ Links ]
Vitt LJ. The ecology of tropical lizards in the Caatinga of northeast Brazil. Occasional Papers of the Oklahoma Museum of Natural History. 1995;1:1-29. [ Links ]
Vitt LJ, Breitenbach GL. Life histories and reproductive tactics among lizards in the genus Cnemidophorus (Sauria: Teiidae). In: Wright JW, Vitt LJ, editors. Biology of whiptail lizards (Genus Cnemidophorus). United States of America: Okalahoma Museum of Natural History; 1993. p. 211-243. [ Links ]
Vitt LJ, Carvalho CM. Niche partitioning in a tropical wet season: Lizards in the lavrado area of northern Brazil. Copeia. 1995;1995(2):305-329. [ Links ]
Vitt LJ, Sartorius SS, Avila-Pires TCS, Espósito MC, Miles DB. Niche segregation among sympatric Amazonian teiid lizards. Oecologia. 2000;122(3):410-420. Doi: 10.1007/s004420050047 [ Links ]
Zaluar HLT, Rocha CFD. Ecology of the wide-foraging lizard Ameiva Ameiva (Teiidae) in a sand dune habitat of Southeast Brazil: ontogenetic, sexual and seasonal trends in food habits, activity, thermal biology and microhabitat use. Ciência e Cultura. 2000;52(2):101-107. [ Links ]
Zar JH. Biostatistical Analysis. 5 ed. New Jersey: Prentice-Hall; 2010. p. 944. [ Links ]
