










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.1 Bogotá Jan./Apr. 2015
https://doi.org/10.15446/abc.v20n1.41610
Artículo de investigación
ANÁLISIS GENÉTICO DE LA POBLACIÓN DE GATOS DEL NORTE Y SUR DE CALI, COLOMBIA
Genetic Analysis of the Cat Population of North and South of Cali, Colombia
Andrés F. PEÑA-CRUZ1, Stephania SANDOVAL ARANGO1, Angie PATIÑO MONTOYA1, Mailyn BEDOYA1, Alejandra RODRÍGUEZ ORTIZ1, Juan ORJUELA VASQUEZ1, Alejandra ORTEGA1, Juan F. LÓPEZ1, Edward MOLINA HENAO1, Angélica GUZMÁN1, Julian GIL1, Heiber CÁRDENAS HENAO2.
1Grupo de Estudio y Trabajo en Genética (GETEG). Universidad del Valle., Calle 13 No. 100-00. Cali, Colombia.
2 Grupo de Investigación en Ecogenética y Biología Molecular. Universidad del Valle. Cali, Colombia.
For correspondence. andres.pcruz@hotmail.com
Received 16th January 2014, Returned for revision 13th May 2014, Accepted 13th June 2014.
Citation / Citar este artículo como: Peña-Cruz AF, Sandoval Arango S, Patiño Montoya A, Bedoya M, Rodríguez Ortiz A, Orjuela Vasquez J, Ortega A, López JF, Molina Henao E, Guzmán A, Gil J, Cárdenas Henao H. Análisis genético de la población de gatos del norte y sur de Cali, Colombia. Acta biol. Colomb. 2015;20(1):109-116. doi: http://dx.doi.org/10.15446/abc.v20n1.41610.
RESUMEN
Los marcadores genéticos del pelaje y malformaciones óseas han permitido caracterizar el perfil genético de más de 400 poblaciones del gato doméstico alrededor del mundo. Hace 15 años se estableció dicho perfil en la ciudad de Cali (Colombia). En este estudio se determinó si el norte y sur de Cali se comportan como subpoblaciones y se comparó el perfil total con el estudio pasado. Se encontró una disminución de la frecuencia alélica de a (no-agouti) y d (dilution), pero un aumento en cinco, especialmente en l (longhair) y cs (siamese). Dichas diferencias pueden atribuirse a la selección humana de características más atractivas y por el flujo génico resultante del crecimiento demográfico de la ciudad, lo que explicaría también el primer reporte de los alelos inhibitor y ticked abyssinian. Se evaluó el equilibrio Hardy-Weinberg para el norte, sur y las dos zonas juntas, usando los loci white spotting y orange, encontrándose desequilibrio en este último para las tres zonas evaluadas debido a un déficit de heterocigotos. Norte y sur se dividieron en dos, y cada sub-muestra presentó equilibrio Hardy-Weinberg, aunque las diferencias en las frecuencias alélicas y heterocigosidades resaltaron microestructura geográfica y una relación entre tiempo de fundación del barrio y heterocigosidad. Norte y sur resultaron ser una población y no subpoblaciones (FST= 0,0004, D= 0,0017), al igual que las nueve poblaciones colombianas con las que se comparó la presente ciudad. Se sugiere realizar un análisis microgeográfico de flujo génico y la definición de posibles colonias de gatos en Cali.
Palabras clave: Felis catus, genes del pelaje, genética de poblaciones, perfil genético, subpoblación.
ABSTRACT
The coat genetic markers and skeleton abnormalities have allowed characterize the profile from more than 400 domestic cat populations around the world. 15 years ago, that profile was established in the city of Cali (Colombia). In this study it was determined if north and south of the city are subpopulations and it was compared the total profile against past study. A decrease in allele frequency of a (non-agouti) and d (dilution) was found, but an increase of five alleles was found, especially in l (long hair) and cs (siamese). These differences could be attributed to human selection of more attractive characteristics and gene flow resulting from demographic growth city, which would also explain the first report of inhibitor and ticked abyssinian alleles. Hardy-Weinberg equilibrium was evaluated for the north, south and both areas together, using white spotting and orange loci, determining disequilibrium in orange for the three evaluated areas due to a heterozygotes deficit. North and south were divided into two, each sub-sample showed Hardy-Weinberg equilibrium, although allele frequencies and heterozygosities highlighted microgeographic structure and a relationship between founding time of the neighborhood and heterozygosity. North and south are a single population and aren´t subpopulations (FST= 0,0004, D= 0,0017), as well as nine Colombian populations with which this city was compared. It is suggested to make a microgeographical gene flow analysis and the definition of possible cat colonies in Cali.
Keywords: coat genes, Felis catus, genetic profile, population genetics, subpopulation.
INTRODUCCIÓN
El gato doméstico (Felis silvestris catus) (Linnaeus, 1758) presenta características que lo hacen interesante para el estudio de la genética de poblaciones, entre ellas: expresan apareamiento aleatorio, son cosmopolitas y exhiben un elevado flujo génico dentro y entre poblaciones, dado que el ser humano se transporta con ellos de un lugar a otro (Robinson, 1985; Ruiz-García y Klein, 1997; Ruiz-García y Álvarez, 2000). Desde 1947 se han llevado a cabo estudios en más de 400 poblaciones de gatos alrededor del mundo (Ruiz-García y Álvarez, 2000; Shostell et al., 2005; Ruiz-García y Álvarez, 2008), usando ciertos marcadores genéticos de fácil identificación fenotípica y que afectan los patrones de coloración del pelaje y malformaciones óseas (Ruiz-García, 1997).
Las frecuencias de estos marcadores genéticos comúnmente usados para dichos estudios poblacionales, pueden verse afectadas por variables ambientales (Shostell et al., 2005) y/o por efectos migratorios que caracterizan la historia del establecimiento de las poblaciones humanas (Todd, 1977). Por lo anterior, procesos de conquista y colonización española y portuguesa en Latinoamérica, han sido de especial interés para el estudio de las poblaciones de gatos establecidas en el continente. En Colombia, se encuentran caracterizados los perfiles genéticos de 20 poblaciones de gatos según estudios de Ruiz-García y Álvarez (1999); Peñuela (2011) y Peñuela y Cárdenas (2012). Dado que siempre se usan para dichos análisis los genes: O (orange, gen ligado al sexo), Ta (tabby), A (agouti), D (dilution), L (longhair), S (white spotting), W (dominant white), C (colour repart) e I (inhibitor), es posible establecer comparaciones para construir árboles de distancias genéticas y FST, lo cual permite construir la historia de migración que ha ocurrido en dichas poblaciones, según como lo muestran Ruiz-García y Álvarez (1999) y Peñuela (2011). Sin embargo, no todas las poblaciones de gatos presentan los mismos eventos de evolución colonizadora (Ruiz-García, 1990; Ruiz-García y Álvarez, 2000).
Estudios de Ruiz-García y Álvarez (1999) reportan el perfil genético para la ciudad de Cali en el año 1997, aunque un estudio reciente realizado por Peñuela (2011) también lo detalla. No obstante, Ruíz-García y Klein (1997) y Ruiz-García y Álvarez (2000), muestran como las poblaciones de gatos, dada su condición de especie social, pueden presentar microestructura, teniéndose la formación de subpoblaciones e incluso de demos dentro de estas. La posible existencia de dicha estructura microgeográfica, llevó a que se planteara como objetivo principal de esta investigación, evaluar si la distancia geográfica que existe entre el norte y sur de la ciudad de Cali, puede ser un factor que pueda determinar si las dos localidades se comportan como subpoblaciones, según sus respectivos perfiles genéticos y análisis de diversidad y de Hardy-Weinberg. Por último, se elaboraron dendrogramas de distancia genética y FST entre los perfiles genéticos obtenidos para la población objeto de estudio y nueve más reportadas previamente para Colombia.
MATERIALES Y MÉTODOS
Se realizó la recolección de los datos durante un año en la ciudad de Cali (Colombia). Los barrios estudiados y su antigüedad fueron en el Norte: Villa del Prado (20 años), Santa Bárbara (18 años), El Sena (53 años), Salomia (60 años); y en el Sur: Meléndez (colonizado hace 73 años y reconocido como barrio desde 1965), Caney (20 años), Ciudadela Comfandi (alrededor de 20 años) La playa (colonizado hace 35 años aproximadamente), Las Vegas (alrededor de 23 años). Se tomó como supuesto que el conjunto de los barrios del norte conformaban una subpoblación, al igual que los del sur. La selección de estos barrios se debió a la gran densidad de viviendas, y además por la separación urbana de 13 km aproximadamente que hay entre norte y sur. El muestreo se llevó a cabo de casa en casa indagando por el sexo del animal y tomando registro fotográfico del patrón de pelaje del individuo. Cabe destacar que no se tenían en cuenta gatos de las calles, ni los que presentaran algún parentesco familiar.
A partir del registro fotográfico se identificaron fenotípicamente los siguientes loci, siguiendo la propuesta del Committee on Standardized Genetic Nomenclature for Cats (1968), excepto para el locus Tabby, pues recientemente Eizirik et al., (2010) por mapeo cromosómico separó el alelo causante del patrón abyssinian como un locus distinto al Tabby, denominándolo Ticked. Los marcadores genéticos inventariados fueron: O (Orange; ligado al sexo), y los autosómicos: B (B, b, bl; color negro vs. color chocolate vs. color carmelita), A (A, a; agouti vs. non-agouti), Ta (Ta(M), Ta(b); mackerel o atigrado vs. tabby blotched), Ti (Ti(A), Ti(+); abyssinian vs. no-abyssinian), D (D, d; full color vs. dilution), L (L, l; pelo corto vs. long hair o pelo largo), S (S, s; manchado de blanco o white spotting vs. no manchado), W (W, w; dominant white vs. color normal), C (C, cs; color normal vs. siamese) y I (I, i; plateado o inhibitor vs. no plateado).
El cálculo de las frecuencias alélicas se efectuó en los loci con dominancia completa asumiendo que existe equilibrio Hardy-Weinberg, según la fórmula: p2+2pq+q2=1. La frecuencia del alelo recesivo se obtuvo como la raíz cuadrada del conteo de homocigotos recesivos de ese locus sobre el total de individuos, mientras que la frecuencia del alelo dominante se obtuvo como p = 1-q. El cálculo de las frecuencias genotípicas y de los valores de chi-cuadrado para evaluar el equilibrio Hardy-Weinberg en los loci white spotting (S) y orange (O) se determinó siguiendo lo planteado por Christensen (2000). Se incluyen valores de heterocigosidad (2pq) para cada locus y de desviación estándar 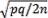 . Estos análisis se hicieron para el norte, sur y las dos zonas juntas (norte + sur). Como se explica más adelante el locus orange no presentó equilibrio Hardy-Weinberg, por lo que se subdividió norte y sur en dos respectivamente, de manera que se agruparon los barrios más cercanos para formar una submuestra, estableciéndose cuatro divisiones: Norte 1 (N1): Villa del Prado y Santa Bárbara, Norte 2 (N2): El Sena y Salomia, Sur 1 (S1): El Caney y Ciudadela Comfandi, Sur 2 (S2): Meléndez, La Playa y Las Vegas. Se evaluó si existía equilibrio Hardy-Weinberg en estas cuatro submuestras, tal cual como se expuso anteriormente.
. Estos análisis se hicieron para el norte, sur y las dos zonas juntas (norte + sur). Como se explica más adelante el locus orange no presentó equilibrio Hardy-Weinberg, por lo que se subdividió norte y sur en dos respectivamente, de manera que se agruparon los barrios más cercanos para formar una submuestra, estableciéndose cuatro divisiones: Norte 1 (N1): Villa del Prado y Santa Bárbara, Norte 2 (N2): El Sena y Salomia, Sur 1 (S1): El Caney y Ciudadela Comfandi, Sur 2 (S2): Meléndez, La Playa y Las Vegas. Se evaluó si existía equilibrio Hardy-Weinberg en estas cuatro submuestras, tal cual como se expuso anteriormente.
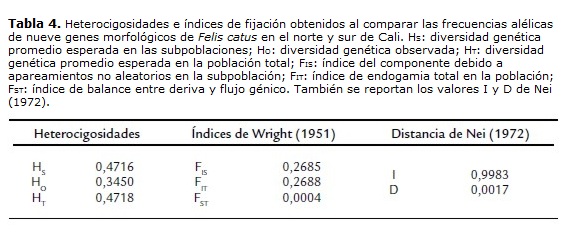
El análisis filogenético se realizó usando los índices de endogamia de Wright (1951) según como dicho estudio lo propone, calculándose la diversidad genética promedio esperada en las subpoblaciones (HS), la diversidad genética observada (HO), la diversidad genética promedio esperada en la población total (HT), el índice del componente debido a apareamientos no aleatorios en la subpoblación (FIS), el índice de endogamia total en la población (FIT) y el índice de balance entre deriva y flujo génico (FST). Además se calculó la distancia de Nei (1972), específicamente la identidad genética normalizada (I) y la distancia genética (D). Este análisis filogenético de Wright (1951) y Nei (1972) se elaboró para el norte y sur de Cali usando los loci O, a, Ta(b), d, l, S, W, cs, I; no obstante asumiendo que ambas zonas representaban a Cali, se comparó con nueve poblaciones colombianas reportadas por Ruiz-García y Álvarez (1999) y Peñuela y Cárdenas (2012), construyéndose árboles filogenéticos de FST y distancia genética (D) usando el algoritmo de Neighbor-Joining en el programa MEGA 5.0 (Tamura et al., 2011).
RESULTADOS
Se inventariaron un total de 263 individuos, 136 en el norte y 127 en el sur de la ciudad. En cuanto al sexo, en el norte se encontraron 41,2% machos y 58,8 % hembras, mientras que en el sur 41,7% machos y 58,3% hembras. Esta igualdad en el porcentaje de sexos encontrados en las dos zonas de la ciudad indican un buen tamaño de muestra para la realización del estudio.
Se encontró que tanto para norte, sur y las dos zonas juntas (norte + sur) los alelos siamese (cs) y long hair (l) presentaron las frecuencias más altas, siendo congruente también con las mayores heterocigosidades reportadas. Por otro lado, el Ti(A) se caracterizó por mostrar la frecuencia más baja en el norte de la ciudad, mientras que en el sur esto ocurrió para el alelo brown (b), teniendo ambos las menores heterocigosidades de su zona. No obstante, cuando se observan los dos sitios en conjunto (norte + sur), se encuentra que tanto el Ti(A) como el white (W) presentan las frecuencias alélicas y heterocigosidades más bajas. Cabe rescatar, empero, que el alelo inhibitor también presenta una de las más bajas frecuencias en el norte, sur y las zonas juntas (norte + sur) de la ciudad (tabla 1).
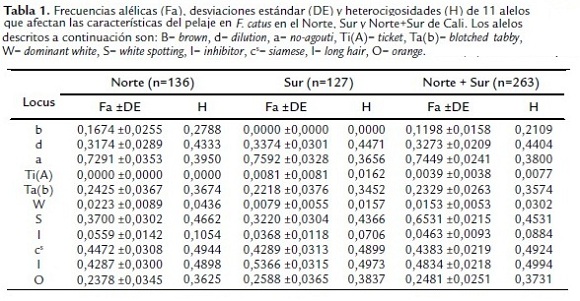
Además, en la tabla 1, se reporta que la desviación estándar mínima encontrada en norte fue para el alelo Ti(A), mientras que en sur fue en brown (b), no obstante, si se asume que norte y sur juntos (Norte + Sur) representan a Cali, se encuentra que es el alelo W el que presenta la menor desviación. Por otro lado, la máxima desviación estándar se encontró en Ta(b) tanto para norte, sur y las dos zonas juntas (norte + sur).
Por otro lado, la prueba de bondad de ajuste de chicuadrado determinó que el locus white spotting se encuentra en equilibrio Hardy-Weinberg tanto en norte, sur, como norte y sur juntas, pero no ocurrió lo mismo con el locus orange como se puede ver en la tabla 2 (no hay equilibrio Hardy-Weinberg en ninguna de las localidades evaluadas), de hecho se observa que hay una deficiencia de heterocigotos considerable con respecto a los valores esperados en norte, sur y las dos juntas (norte + sur) para este locus.
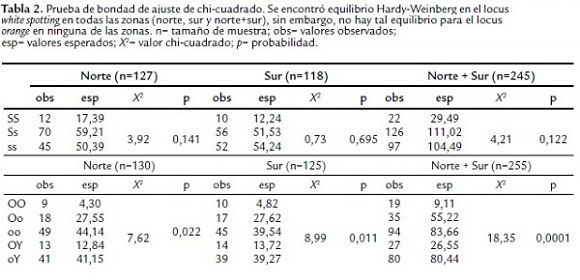
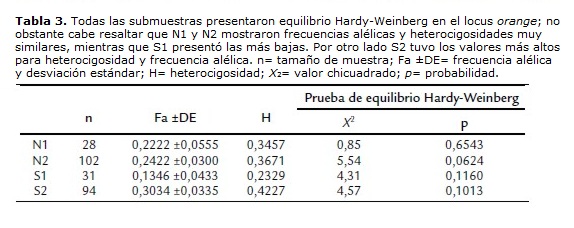
En vista que el locus orange no presentó equilibrio Hardy-Weinberg, se dividió norte y sur en dos respectivamente, agrupando los barrios más cercanos. De esta manera se obtuvieron cuatro submuestras compuestas por los siguientes barrios: Norte 1 (N1): Villa del Prado y Santa Bárbara, Norte 2 (N2): El Sena y Salomia, Sur 1 (S1): El Caney y Ciudadela Comfandi, Sur 2 (S2): Meléndez, La Playa y Las Vegas. Para estas cuatro submuestras se determinó equilibrio Hardy-Weinberg, encontrándose diferencias particulares entre estas.
Mientras que N1 y N2 presentaron frecuencias alélicas y heterocigosidades casi idénticas, S1 presentó las más bajas, y S2 exhibió las más altas (tabla 3).
El análisis filogenético (tabla 4) tuvo en cuenta los alelos más informativos: O= orange, a= no-agouti, tb= blotched tabby, d= dilution, l= long hair, S= white spotting, W= dominant white, cs = siamese e I= inhibitor. Se encontró que el norte y sur de Cali se comportan como una sola población y no como subpoblaciones, probando en este caso, que la distancia geográfica no ha sido un factor determinante en la subdivisión poblacional, según el análisis de FST de Wright (1951) y distancia de Nei (1972): Identidad genética normalizada (I) y Distancia genética (D) (FST= 0,0004; I= 0,9983; D= 0,0017). La HT y HS indican que no ha habido diferenciación genética entre las frecuencias alélicas del norte y sur de Cali, no obstante la heterocigosis observada (HO) es menor a las esperadas (HT y HS) lo cual muestra una diversidad genética menor de la que se esperaba, explicándose por un posible efecto de endogamia, que aunque es pequeño (FIT= 0,2688; FIS= 0,2685) argumentaría lo observado.
Al compararse las frecuencias alélicas reportadas por Ruiz-García y Álvarez (1999) con las del presente estudio en la ciudad de Cali (asumiendo que norte y sur representan en conjunto a Cali) se encuentra que durante los años transcurridos los siguientes alelos se reportaron en mayor frecuencia, especialmente los alelos l y cs según como se menciona respectivamente: O (0,1930 vs. 0,2481), Ta(b) (0,1800 vs. 0,2329), l (0,3300 vs. 0,4834), cs (0,2690 vs. 0,4383), I (0,0000 vs. 0,0463). Mientras que los alelos a (0,8390 vs. 0,7449) y d (0,4710 vs. 0,3273) disminuyeron.
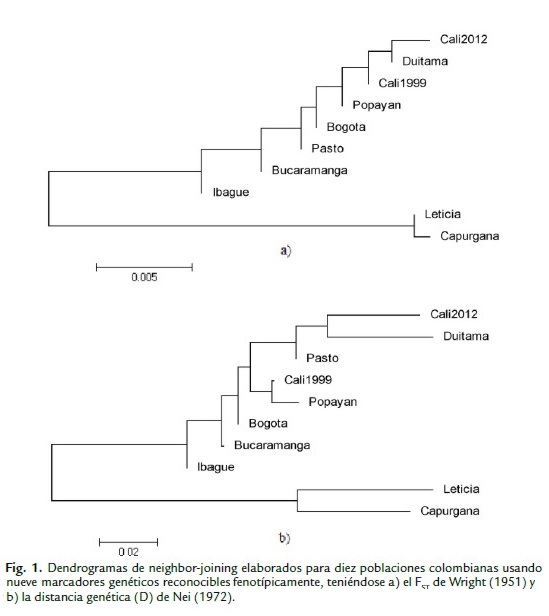
Los dendrogramas de Neighbor-Joining elaborados para los valores de FST de Wright (1951) y de distancia genética (D) de Nei (1972) para las diez poblaciones Colombianas, presentan un grupo externo formado por Leticia y Capurganá, siendo estás las que mayor diferenciación genética han presentado frente al resto. También, ambos dendrogramas agruparon a Cali 2012 con Duitama, dada la similaridad en las frecuencias alélicas encontradas para ambas (Figs. 1a y b). Cali 1999, Popayán y Bogotá formaron un grupo del cual Pasto estaba cerca, sin embargo, esta ciudad presenta una diferenciación genética similar a la de Bogotá (Fig. 1a), pero la distancia genética la agrupa con Duitama y Cali 2012 (Fig. 1b).
DISCUSIÓN
Los loci l, Ta(b), I, cs, O, a y d presentaron diferencias relevantes con respecto al estudio de Ruiz-García y Álvarez (1999) en la ciudad de Cali; esto podría explicarse quizá por el hecho que hay una selección por parte de los humanos hacia estos alelos por cuestiones estéticas, aumentando así las frecuencias al igual que la heterocigosis esperada. Además, otra posible respuesta podría ser el rápido crecimiento poblacional de gatos lo cual incrementa considerablemente el flujo genético e incrementa la panmixia. Por otro lado, Ruiz-García y Álvarez (1999) plantean que la escases del locus White puede ser usado como medidor de diversidad genética. Para el presente estudio se obtuvo una frecuencia del alelo W ligeramente mayor con respecto al estudio hecho en Cali por Ruiz-García y Álvarez (1999), lo cual según estos, puede ser un detector de la cantidad de intervención humana en las poblaciones de gatos por lo que se podría pensar que para este trabajo dicha intervención fue mayor. No obstante, el hecho que la frecuencia del alelo W sea tan baja en ambos estudios, puede ser atribuido a efectos pleiotrópicos sobre la audición (Geigy et al., 2007) lo cual podría causar problemas en los individuos, así como una muerte más temprana. Es posible, que el aumento de la frecuencia del locus long hair (l) se deba a la preferencia de los humanos por los gatos de pelo largo, dado que son más atractivos como mascotas; con esto han aumentado en Cali, las tiendas que se dedican a importar estos animales para la venta dentro y fuera de la ciudad. Otra hipótesis que podría explicar por que la frecuencia del long hair aumentó con respecto al estudio de Ruiz-García y Álvarez (1999), puede deberse a que recientemente Kehler et al., (2007) determinaron que dicho locus está presente en distintas formas fenotípicas, es decir que un gato puede tener el alelo que causa el pelo largo y expresarlo fenotípicamente en sólo una parte de su cuerpo, es posible que Ruiz-García y Álvarez (1999) ignoraran esto, pues no lo aclaran en la metodología. La baja frecuencia del alelo Ti(A) fue un resultado esperado teniendo en cuenta que no fue reportado en 1999 para Cali. La presencia de este carácter actualmente puede revelar la introgresión de dicho carácter asiático, como sucedió con México en el 2005 (Ruiz-García et al., 2005). Cabe aclarar que recientemente Eizirik et al., (2010) separó el patrón abisiano como un locus distinto al tabby.
El alelo que causa el color inhibitor (I), no se reportó en Ruiz-García y Álvarez (1999), pero en el presente estudio apareció con una frecuencia de 0,0463, lo cual podría justificarse por los cambios en la tasa de crecimiento y estructura de la población caleña, dado que desde hace 13 años muchas personas se trasladaron a la ciudad junto con sus mascotas; además, los desplazamientos llevados a cabo en Colombia por el conflicto armado se han intensificado en los últimos años, como consecuencia en el periodo de 19992005 llegaron a Santiago de Cali más de 55000 personas, de los cuales casi 20000 fueron en el año 2005 (Fundación Foro Nacional por Colombia, 2006). Los eventos de migración de los gatos están sometidos al traslado de sus dueños, siendo un factor importante en el cambio del flujo génico; una vez que una población humana alcanza los 30000 habitantes, su población de gatos puede haber alcanzado un número efectivo entre 1000 y 5000 individuos, por lo que el impacto de la deriva genética es bastante limitado, siendo la migración un factor más explicativo (Morril y Todd, 1978). Esta hipótesis de migraciones humanas (incluidos los desplazamientos) puede aplicarse a todos los loci, pero en especial puede ser una explicación para argumentar por qué se reportó el inhibitor 15 años después. No se descarta igualmente, la selección por novedad, pues como se mencionó previamente, este carácter, pudo ser seleccionado por razones estéticas por los humanos, de tal manera que sus frecuencias fueron elevándose progresivamente (Ruiz-García y Álvarez, 1999), aun así no se tienen pruebas suficientes para determinar que esta haya sido la razón.
Usualmente, como menciona Ruiz-García y Álvarez (1999), los dueños de este tipo de mascotas, se movilizan por toda la ciudad con éstos animales, abandonando las crías o regalándolas, de esta forma el flujo génico es alto y era de esperarse que las frecuencias alélicas de los genes evaluados no hayan diferido sustancialmente entre los muestreos del Sur y el Norte de Cali, lo cual fue corroborado por el análisis de FST, ya que ambas zonas se comportan como una sola población (FST=0,0004). Se pudo comprobar el equilibrio H-W para el locus white spotting tanto en las dos localidades (norte y sur) separadas, como en conjunto; este gen por ser autosómico pudo alcanzar el equilibrio rápidamente (Hardy, 1908), dado que el tiempo de ingreso de los gatos a América fue aproximadamente hace 500 años, traídos de España y Portugal por actividades comerciales (Ruiz y Álvarez, 2003). Por el contrario, cuando las frecuencias alélicas difieren entre sexos, el equilibrio H-W no se alcanza en una generación; para encontrarse en equilibrio deben pasar aproximadamente cinco generaciones, lo que se cumple si las frecuencias alélicas están en grandes números y si la población es panmíctica (Soler, 2003). Entonces el locus orange debería estar en equilibrio, pues muchas generaciones han pasado durante 500 años, no obstante, debe considerarse la microestructura e historia de los barrios muestreados, como Meléndez, el cual empezó su asentamiento a finales de la década de 1940 (Dagma, 2009).
Observando los valores de heterocigosidad para el locus orange, se detalló que esta fue mayor en S2 (0,4227) en comparación con los otros lugares (N1: 0,3456, N2: 0,3671, S1: 0,2329). Una posible razón de esto, es que S2 fue poblado desde la década de 1940, mientras que los otros comenzaron a ser colonizados varias décadas después; por ejemplo, S1, correspondiente al barrio El Caney se fundó en 1993 (Vásquez, 2001), siendo el barrio más nuevo de los implicados en el muestreo y con el menor valor de heterocigosidad. Si se asume una relación entre el tiempo de fundación de los barrios y el flujo génico, podría considerarse que entre mayor sea esta mayor será el flujo; de acuerdo con ello, podría ser una explicación al hecho de que el barrio más antiguo presente mayor heterocigosidad. Igualmente, de acuerdo a Ruiz-García (1994) este locus puede afectarse debido una preferencia fenotípica por parte de la ciudadanos, a favor de los homocigotos.
El valor de FST en conjunto con el de distancia genética (FST= 0,0004; I=0,0017) obtenido para norte y sur refleja que ambas localidades no son subpoblaciones lo cual puede argumentarse por varias razones: por un control de los seres humanos en la generación de camadas con paternidades simultáneas, incrementando así en gran manera el flujo genético dentro de la poblaciones, reduciendo la endogamia e incrementando la panmixia; además, no hay barreras que limiten el flujo de los gatos, por medio de sus dueños de un lado de la ciudad al otro. Sin embargo, el hecho que el FIS y FIT presenten valores de 0,268 indica que si hay cierto nivel de endogamia en las poblaciones locales norte y sur, evidenciando una deficiencia en la heterocigosidad. Cabe aclarar que el valor del FST se debe al sistema de apareamiento y no a la subdivisión poblacional.
Ahora bien, al comparar la ciudad de Cali con otras ciudades colombianas se encontró que todas son similares genéticamente. La alta homogeneidad genética encontrada en gran parte de Colombia puede estar relacionada con los movimientos rápidos de los conquistadores españoles a través del río Magdalena, los cuales establecieron muchas ciudades y villas en poco tiempo (Ruiz-García et al., 2005). Sin embargo, Cali 2012 se observa muy cercana a Duitama y Cali de 1999 pero también a Popayán, lo que concuerda con lo encontrado por Ruiz-García y Álvarez (1999); esto puede deberse a que los municipios de Popayán y Cali fueron centros administrativos territoriales de España durante la colonia en el Valle geográfico del río Cauca y al ser las únicas poblaciones presentes en la época, el flujo de habitantes, y con ellos sus animales domésticos, debió ser permanente (Peñuela, 2011); se podría pensar entonces que su permanente comunicación por el curso del río Cauca permitió una similitud genética entre las poblaciones fuentes de gatos que darían origen al resto.
Generalmente nunca se ha visto agrupada a Duitama con otras ciudades colombianas, a lo que se ha atribuido un fuerte efecto fundador y deriva genética (Ruiz-García y Álvarez, 1999; Ruiz-García y Álvarez, 2008). El que se encuentre agrupado con Cali podría deberse a que la introgresión de genes como Inhibitor, afectó en gran parte la diversidad genética de Cali, semejándola a Duitama, lo que pudo ser maximizado por otras variables como la selección por novedad o los fuertes desplazamientos que se han vivido en el presente siglo. Por otro lado, se observa a Leticia completamente separada del resto de ciudades, lo que concuerda con estudios anteriores; Ruiz-García y Álvarez (1999) reportaron a Leticia como la población más diferente de otras 20 poblaciones latinoamericanas estudiadas y, sugirieron un fuerte efecto fundador asociado a deriva genética y/o un probable origen diferente a las otras poblaciones, además posible pérdida de variabilidad genética. Peñuela y Cárdenas (2012) reportaron igualmente que Capurganá, se agrupa con Leticia, lo cual es incoherente si se tiene en cuenta que ambas poblaciones son las más distantes geográficamente. Tampoco podría explicarse la agrupación por las migraciones humanas, pues Ruiz-García et al. (2005), Peñuela (2011) y Peñuela y Cárdenas (2012) señalan que Capurganá y Leticia no solo difieren en su tiempo de fundación, sino que también en la historia de su colonización y de relaciones comerciales con otras regiones. Ahora bien, la perdida de alelos parece ser la explicación más acertada por la cual Leticia y Capurganá hayan agrupado. Aunque las diez poblaciones evaluadas presenten una historia de colonización diferente o quizá similar, dados los movimientos de conquista españoles al interior de la nación, todas reflejan un origen común, lo cual explicaría por qué el grado de diferenciación y distancia genética entre ellas es reducido.
La distancia de Nei (1972) indicó que Cali se comporta genéticamente como una sola población, y al compararla con otras ciudades, hubo una alta similitud entre ellas; confirmando lo encontrado por el FST. En general, la similitud genética entre las poblaciones latinoamericanas puede deberse a un evento fundador en común, que refleja su origen idéntico en fuentes españolas (Ruiz-García et al., 2002). Ruiz-García et al., (2005) argumentan que la comparación de las frecuencias alélicas en las poblaciones de gatos, ha llevado a la identificación de relaciones genéticas entre las poblaciones y las rutas de migración de los europeos. Esta especie no estaba presente en el continente americano hasta la llegada de los europeos y su introducción debió estar profundamente ligada a las rutas colonizadoras y comerciales de las diferentes potencias europeas en el Nuevo Mundo (Ruiz-García y Álvarez, 1999), como lo mencionado para Cali y Popayán. Es por esto que poblaciones del suroccidente colombiano guardan gran similitud, ya que históricamente fueron originadas por la misma población de origen; los pequeños grupos genéticos que se forman pueden deberse a frecuencias de alelos raros que son comunes entre ellos.
CONCLUSIONES
El norte y sur de Cali no han presentado diferenciación genética, por lo que se comportan como la misma población y no como subpoblaciones, explicando así la facilidad con la que existe flujo genético entre los barrios de la ciudad; aunque, debe tenerse en cuenta que la diversidad genética parece verse influenciada por el tiempo de construcción o colonización de los barrios, sin embargo no se descarta que estén ocurriendo otros eventos a nivel microgeográfico que caractericen cada lugar. Los procesos de inmigración y emigración humana en la ciudad son un factor importante que ha determinado procesos de flujo génico, por lo que no es de extrañar, que a pesar de todo, las frecuencias alélicas puedan variar con el pasar de los años, como se determinó al compararse con el estudio realizado hace aproximadamente 15 años en Cali. No obstante, este cambio en las frecuencias parece ser más visibles a un nivel microgeográfico, por lo que un estudio detallado de la microestructura a nivel de "colonias" podría dar luz para entender que procesos de colonización y de flujo ocurren en la microestructura de la población de la ciudad de Cali. Por último, los valores de distancia genética y FST son muy similares entre las poblaciones colombianas evaluadas, reflejando así un origen común.
AGRADECIMIENTOS
Agradecemos a Mauricio Peñuela Aristizabal por la asesoría brindada para la elaboración del análisis filogenético y la redacción del documento, también agradecemos a los integrantes del Grupo de Estudio y Trabajo En Genética (GETEG), por su apoyo logístico durante la recolección de datos y a los dueños de las mascotas, quienes permitieron que este estudio fuera posible.
REFERENCIAS
Christensen A. Cats as an aid to teaching genetics. Genetics. 2000;155(3):999-1004. [ Links ]
Committee On Standardized Genetic Nomenclature For Cats. Standardized genetic nomenclature for the domestic cat. J Hered. 1968;59(1):39-49. [ Links ]
Dagma. Agenda ambiental, Comuna 18. Consejo Ambiental de Cali 2009. Available at: URL: http://consejoambiental.files.wordpress.com/2009/10/comuna-181.pdf. Cited 9 Jun 2014. [ Links ]
Eizirik E, David V, Buckley-Beason V, Roelke M, Schäffer A, Hannah S, et al. Defining and mapping mammalian coat pattern genes: multiple genomic regions implicated in domestic cat stripes and spots. Genetics. 2010;184(1):267-275. Doi: 10.1534/genetics.109.109629. [ Links ]
Fundación Foro Nacional Por Colombia. Foro debates (No. 5): pobreza y exclusión social en Bogotá, Medellín y Cali. Fundación Foro Nacional por Colombia. Bogotá, D. C.; 2006. p. 72-73. [ Links ]
Geigy C, Heid S, Steffen F, Danielson K, Jaggy A, Gaillard C. Does a pleiotropic gene explain deafness and blue irises in white cats?. Vet J. 2007;173(3):548–553. Doi: 10.1016/j.tvjl.2006.07.021. [ Links ]
Kehler JS, David V, Schäffer A, Bajema K, Eizirik E, Ryugo D, et al. Four independent mutations in the feline fibroblast growth factor 5 gene determine the long-haired phenotype in domestic cats. J Hered. 2007;98(6):555-566. Doi: 10.1093/jhered/esm072. [ Links ]
Morril R, Todd N. Mutant allele frequencies in the domestic cats of Denver, Colorado. J Hered. 1978;69(2):131-134. [ Links ]
Nei M. Genetic distance between populations. Am Nat. 1972;106(949):283-292. Doi: 10.1086/282771. [ Links ]
Peñuela M. Marcadores genéticos del pelaje del gato doméstico Felis catus (FELIDAE) en el suroccidente colombiano (Tesis de pregrado). Cali: Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle; 2011. 75 p. [ Links ]
Peñuela M, Cárdenas H. Marcadores genéticos del pelaje en gatos domésticos de Capurganá-Colombia. Momentos de Ciencia. 2012;9(1):28-33. [ Links ]
Robinson R. The domestic cat in genetic research. Trends Genet. 1985;1:236-239. Doi: 10.1016/0168-9525(85)90090-3. [ Links ]
Ruiz-García M. Frecuencias alélicas en la población de gatos domésticos de la isla de Menorca (Baleares): diferentes modelos de evolución colonizadora. Evol Biol. 1990;4:307-342. [ Links ]
Ruiz-García M. Genetic profiles from coat genes of natural Balearic cat populations: an eastern Mediterranean and North-African origin. Genet Sel Evol. 1994;26(1):39-64. Doi: 10.1186/1297-9686-26-1-39. [ Links ]
Ruiz-García M. Genetic relationships among some new cat populations sampled in Europe: a spatial autocorrelation analysis. J Genet. 1997;76(1):1-24. Doi: 10.1007/BF02931765. [ Links ]
Ruiz-García M, Álvarez D. Análisis filogenético de 21 poblaciones latinoamericanas de gatos, mediante 10 loci morfológicos, utilizando métodos de matrices de distancias genéticas y de máxima parsimonia. Bol R Soc Esp Hist Nat (Sec Biol). 1999;95(3-4):139-164. [ Links ]
Ruiz-García M, Álvarez D. Genetic microstructure in two spanish cat populations. I: genic diversity, gene flow and selection. Genes Genet Syst. 2000;75(5):269-280. Doi: 10.1266/ggs.75.269. [ Links ]
Ruiz-García M, Álvarez D. Análisis de seis poblaciones latinoamericanas de gatos mediante genes del pelaje y marcadores microsatélites. Acta Zool Mex (ns). 2003;89:261-286. [ Links ]
Ruiz-García M, Álvarez D. A biogeographical population genetics perspective of the colonization of cats in Latin America and temporal genetic changes in Brazilian cat populations. Genet Mol Biol. 2008;31(3):772-782. Doi: 10.1590/S1415-47572008000400026. [ Links ]
Ruiz-García M, Álvarez D, Shostell J. Population genetic analysis of cat populations from México, Colombia, Bolivia, and the Dominican Republic: identification of different gene pools in Latin America. J Genet. 2005;84(2):147–171. Doi: 10.1007/BF02715841. [ Links ]
Ruiz-García M, Campos H, Álvarez D, Kajon A, Díaz S. Coat gene profiles of several cat populations in Cuba, Costa Rica, Colombia, Paraguay, Chile and Argentina, and possible genetic origins of these cat populations. Russ J Genet. 2002;38(2):165-180. Doi: 10.1023/A:1014334111142. [ Links ]
Ruiz-García M, Klein K. Genetic structure of populations of the domestic cat in Catalonia (Spain) and upper midwestern USA: a microgeographic and macrogeographic study. J Genet. 1997;76(2):99-115. Doi: 10.1007/BF02923555. [ Links ]
Shostell J, Staudinger J, Ruiz-García M. Mutant allele frequencies in domestic cat populations in Arkansas and Tennessee. J Hered. 2005;96(5):557-565. Doi: 10.1093/jhered/esi072. [ Links ]
Soler M, editor. Evolución la base de la biología. Barcelona, España: Proyecto sur de ediciones, S.A.L.; 2003. 559 p. [ Links ]
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731-2739. Doi: 10.1093/molbev/msr121. [ Links ]
Todd N. Cats and commerce. Sci Am. 1977;237(5):100-107. Doi: 10.1038/scientificamerican1177-100. [ Links ]
Vásquez E. Historia de Cali en el siglo 20: sociedad, economía, cultura y espacio. Cali, Colombia: Universidad del Valle; 2001. p. 293-294. [ Links ]
Wright S. The genetical structure of populations. Ann Eugen. 1951;15(1):323-354. Doi: 10.1111/j.1469-1809.1949.tb02451.x. [ Links ]
