










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.2 Bogotá May/Aug. 2015
https://doi.org/10.15446/abc.v20n2.43130
Doi: http://dx.doi.org/10.15446/abc.v20n2.43130
Artículo de investigación
COMPARING INOCULATION METHODS TO EVALUATE THE GROWTH OF Xanthomonas axonopodis pv. manihotis ON CASSAVA PLANTS
Comparación de métodos de inoculación para evaluar el crecimiento de Xanthomonas axonopodis pv. manihotis en plantas de yuca
Alejandra MUÑOZ BODNAR1, Leidy Mariel CRUZ GÓMEZ2, Adriana BERNAL3, Boris SZUREK4, Camilo Ernesto LÓPEZ CARRASCAL1.
1Manihot Biotec, Departamento de Biología. Universidad Nacional de Colombia. Cra. 30 nº. 45-03. Bogotá, Colombia.
2Universidad Pedagógica y Tecnológica de Colombia. Avenida Central del Norte 39-115. Tunja, Colombia.
3LAMFU, Departamento de Biología. Universidad de los Andes. Carrera Primera nº. 18A-12. Bogotá, Colombia.
4Résistance des Plantes aux Bioagresseurs (RPB). Institut de Recherche pour le Développement. 911, Av. Agropolis BP 64501. Montpellier, Francia.
For correspondence. celopezc@unal.edu.co
Received: 24 April 2014; Returned for revision: 23 September 2014; Accepted: 3 November 2014.
Associate Editor: Leonardo Galindo.
Citation / Citar este artículo como: Muñoz Bodnar A, Cruz Gómez LM, Bernal A, Szurek B, López Carrascal CE. Comparing inoculation methods to evaluate the growth of Xanthomonas axonopodis pv. manihotis on cassava plants. Acta biol. Colomb. 2015;20(2):47-55. doi: http://dx.doi.org/10.15446/abc.v20n2.43130
RESUMEN
Xanthomonas axonopodis pv. manihotis (Xam) es el agente causal del tizón bacteriano de la yuca, una de las principales enfermedades de los cultivos de yuca en América del Sur y África. Hasta ahora, el desarrollo de la enfermedad se mide a través de AUDPC (Area Under Disease Progress curve), pero no hay disponibles métodos cuantitativos fiables, esto debido posiblemente a la alta variabilidad del crecimiento bacteriano en la planta. Para establecer un método exacto para la cuantificación bacteriana durante el curso de la infección Xam dentro de los tejidos del huésped, se analizaron las poblaciones de bacterias sobre tallo y hojas, así como corte de hojas de las variedades de yuca MCOL1522 y SG107-35 con la cepa virulenta CIO151 Xam. En esta investigación se muestra que el movimiento de las bacterias a lo largo de los tejidos y especialmente en las hojas es estocástico. Por otra parte, hemos podido demostrar el crecimiento diferencial de la cepa virulenta Xam CIO151 tras la punción al tallo y la cuantificación de la bacteria a 6 cm de distancia del punto de inoculación de dos variedades que presentan niveles contrastantes de susceptibilidad.
Palabras clave: crecimiento bacteriano, métodos de inoculación, yuca.
ABSTRACT
Xanthomonas axonopodis pv. manihotis (Xam) is the causal agent of cassava bacterial blight (CBB), a major disease for cassava crops in South America and Africa. Until now the development of the disease is measured via AUDPC (Area Under Disease Progress Curve) but no reliable quantitative methods are available probably due to high variability of bacterial growth in planta. To establish an accurate method for bacterial quantification during the course of Xam infection within the host tissues, we analyzed bacterial populations upon stem and leaf-puncturing as well as leaf-clipping of cassava varieties MCOL1522 and SG107-35 challenged with the virulent Xam strain CIO151. Here, we show that the movement of bacteria along the tissues and especially in leaves is stochastic. Moreover, we were able to demonstrate differential growth of virulent Xam strain CIO151 upon stem-puncturing and quantification of bacteria 6 cm. away from the inoculation point of two varieties displaying contrasting levels of susceptibility.
Keywords: bacterial growth, cassava, inoculation methods.
INTRODUCTION
Cassava Bacterial Blight (CBB) is one of the most prevalent bacterial diseases of cassava, which can severely compromise its production, and therefore food security for more than 1000 million people, who depend on cassava starch as main source of calories (FAO, 2011; López and Bernal, 2012). CBB is caused by the gram-negative bacteria Xanthomonas axonopodis pv. manihotis (Xam) which enters plant tissues via wounding and colonizes the vascular tissues ( Verdier, 2002). Xam is a highly diverse pathogen in Latin America, as well as in Africa, albeit to a lower extend (López and Bernal, 2012). The losses caused by Xam can reach 100 % of cassava production after three cycles of production if the environmental conditions are favorable (Verdier, 2002). Only a few resistant cassava varieties have been reported, which are mainly non-commercial varieties (Restrepo et al., 2000a). Histological and molecular defense responses of cassava against Xam have shown to be activated in both resistant and susceptible varieties, but are faster and more intense in resistant plants (Kpémoua et al., 1996; López et al., 2005 ). As an important tool for improving the study of the cassava-Xam pathosystem, the cassava genome sequence was recently released (http://www.phytozome.net, Prochnik et al., 2012). Also, the draft genome sequences of 65 Xam strains as well as the complete genome sequence of the reference Xam strain CIO151 were recently reported and are available (Bart et al., 2012; Arrieta et al., 2013), These important genomic resources will be crucial for the identification of key cassava and Xam genes determining resistance and susceptibility of cassava plants challenged with Xam.
To phenotypically validate the function of putative cassava resistance genes, it is important to evaluate their effect on the growth of Xam populations in planta. Similarly, to evaluate the role played by Xam virulence/pathogenicity candidate genes, it is essential to evaluate the effect of the inactivation of these genes by following the growth of relevant mutant strains in susceptible cassava plants. Thus, the quantification of bacterial populations in planta is pivotal to demonstrate the function of plant and bacterial genes during infection, and complementary to the observation of macroscopic plant phenotypes. Traditionally, bacterial populations are quantified by evaluating a dilution series of infected plant tissues extracts plated on selective medium, but the inoculation methods vary depending on the studied pathosystem. The target tissues that are naturally colonized by the pathogen as well as the plant anatomy are key criteria to chose the most appropriate inoculation method (Meyer etal., 2005). For example, in the Arabidopsis-Pseudomonas syringae pathosystem depending on the inoculation method, resistance mechanisms may or may not be triggered (Melotto etal., 2006). In other pathosystems, such as in the interaction of Xanthomonas campestris pv. campestris with Arabidopsis, several inoculation methods are reported, but some are more prone to generate variability in plant responses and bacterial growth (Meyer et al., 2005). To tackle this problem, Meyer et al., (2005) reported on wound inoculation as the optimal method to avoid variability Interestingly, the function of the avirulence gene avrACXcc8004 in this pathosystem could only be evidenced upon puncturing but not by leaf infiltration, revealing that avrACXcc8004 is specifically recognized within vascular tissues (Xu et al., 2007). Hence, the method of inoculation plays an essential role in the study of a pathosystem of interest and it is, therefore essential to find what is the best one mimicking the pathogen infection under natural conditions.
Wounding, syringe-infiltration and spray inoculation are a few of the most prevalent methods to inoculate bacteria. In tomato, Xanthomonasaxonopodis pv. vesicatoria, which colonizes the apoplast of the leaf mesophyl, is often inoculated by leaf infiltration (Kim et al., 2008). Xanthomonas oryzae pv. oryzae colonizes the vascular tissues of rice and the most common inoculation method for quantitative assessment is leaf-clipping, which consists in cutting leaf tips with scissors previously immersed in a bacterial suspension. In cassava, several methods of inoculation of Xam into the plant have been evaluated in material grown from stem cuttings or in vitro (Jorge and Verdier, 2002). These methods include clipping leaves with contaminated scissors (Lozano and Laberry, 1982), spraying foliage, depositing bacterial suspensions on mechanically generated leaf holes and stem puncturing (Restrepo et al., 2000b; Jorge and Verdier, 2002). Stem inoculation of plants grown in vitro was used to describe Xam colonization inside the plant through histological approaches (Kpémoua et al., 1996). Interestingly, this study reported the presence of Xam agglomerates randomly distributed along the tissues. This could explain why most of the methods employed to quantify bacteria in planta produce a high variability, thereby making them inadequate for resistance screening and loss of pathogenicity assays. Additionally, microarray studies demonstrated that cassava gene expression upon Xam infection is quite delayed as compared to other pathosystems (López et al., 2005). This is also correlated to the observation that primary symptoms appear only 8 days-post inoculation of 4-month old susceptible cassava plants with a virulent strain of Xam (López et al., 2005). At present, the reference method to evaluate cassava resistance is through area under the disease progress curve (AUDPC). However, this method is based on the visual inspection of disease intensity over time and does not measure bacterial populations in planta, which is particularly relevant considering the quantitative nature of the resistance to CBB (Jorge and Verdier, 2002).
The aim of this work was to evaluate different inoculation methods relevant to the Xam-cassava pathosystem, quantifying bacterial growth and following up the symptomatic development in order to establish an accurate method with reduced variability. Hence, despite the observation of high variability in bacterial populations, we show that the most accurate evaluation method is by determining bacterial growth away from the site of inoculation point in the stem of cassava plants grown in greenhouse.
MATERIAL AND METHODS
Bacterial strains
Xam strain CIO151 was streaked onto YPGA medium (5 % yeast extract, 5 % peptone, 5 % glucose and 15 % agar) and incubated at 28 °C during 48 hours. One colony was grown onto YPG medium and incubated at 28 °C for 18 h before inoculation. Liquid inoculums were calibrated at OD600nm= 0.002, corresponding to 106 colony forming units (CFU).
Greenhouse plants assays
Cassava varieties MCOL1522 and SG10735 were grown in greenhouse conditions (day/night temperature = 28/19ºC, 12h photoperiod, and 80 % relative humidity). Leaves of 8-weeks-old plants were used to test the puncturing and clipping inoculation methods. For puncturing, a sterile needle was used to make a the incision on the main nerve of the central lobe. Five microliters of calibrated inoculums (OD600nm=0.002) were placed on the puncture. For clipping, scissors were immersed in the inoculum solutions and a transversal section was made 1 cm. below the tip of the leaf. For both methods, the bacterial population was quantified at 0, 7, 14 and 21 days post inoculation (dpi). Depending on the experiment, different sections of stem or leaves were collected.
For stem inoculation, eight week-old plants were used. Puncturing with a sterile toothpick was made on the third internode from the apical region. Ten microliters of calibrated inoculums were placed on the puncture. Different leaf or stem sections of were collected at 0, 7, 14 and 21 dpi, depending on the experiment.
AUDPC
Eight week-old cassava plants were inoculated in the stem with the strain CIO151. Plants were inoculated and evaluated at 0, 7, 14, 21 and 28 dpi following a severity scale 0-5 (Jorge et al., 2000) where 0- healthy plant, no reaction observed, 1- dark area or necrosis around the inoculation point; 2- gum exudates on the stem; 3- wilting of one or two leaves and exudates; 4- more than two leaves wilted and 5- complete wilting and dieback. Ten biological replicates were used for each variety AUDPC was calculated as described previously (Jorge et al., 2000).
Quantification of bacterial population
For every experiment, three technical repetitions from three distinct plants of each variety were used. Leaves and stem sections were collected, cleaned in 70 % ethanol and sterile water and disinfected with 2 % sodium hypochlorite and ground in sterile water. Serial dilutions were plated on YPGA medium and incubated at 28 °C. After 48 hours Xam colonies formed were counted. Non-inoculated plants were used as controls.
All the assays were performed with three biological repetitions.
Statistical analyses
The statistical Wilcoxon test was used as a non-parametric test to compare related samples. For AUDPC, a t-test was employed. Data from three biological repetitions (consisting of three technical replicate) were generated to complete statistical analyses.
RESULTS
Susceptibility level
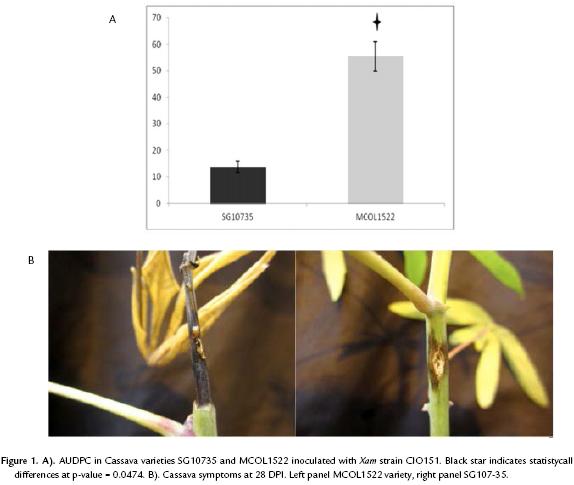
In order to compare the susceptibility level of the two cassava varieties that were used in our study, we applied a qualitative standard method (AUDPC), to discriminate between resistant and susceptible, which relies on the use of a symptoms scale. Through this approach the phenotype of a given variety can be quantified in a rank from 0 to 122.5, 0 indicating the plant to be highly resistant and 122.5 highly susceptible (Jorge and Verdier, 2002). As a result of the analysis of three independent biological replicates, the obtained average AUDPC value were 13.8 and 55.5 for the varieties SG107-35 and MCOL1522, respectively (Fig. 1A and B, indicative of a significant difference in the susceptibility level of the two cassava varieties to Xam strain CIO151 (t-test with p-value=0.0474). Interestingly, symptoms observed in MCOL1522 appear earlier than in SG10735, where the plants remain healthy at 7 dpi and symptoms increase quickly between 14 and 21 dpi. More than one plant of COL1522 variety showed a symptom scale superior to 4, which is the criterion employed by Restrepo et al., (2000b) to be considered as susceptible. On the other hand, none SG107-35 plants showed this symptom scale and in consequence following the same criterion this variety is considered as resistant. Overall, these data demonstrate that MCOL1522 and SG10735 are useful varieties for our purpose to evaluate the sensitivity of different inoculation methods.
Leaf-clipping inoculation
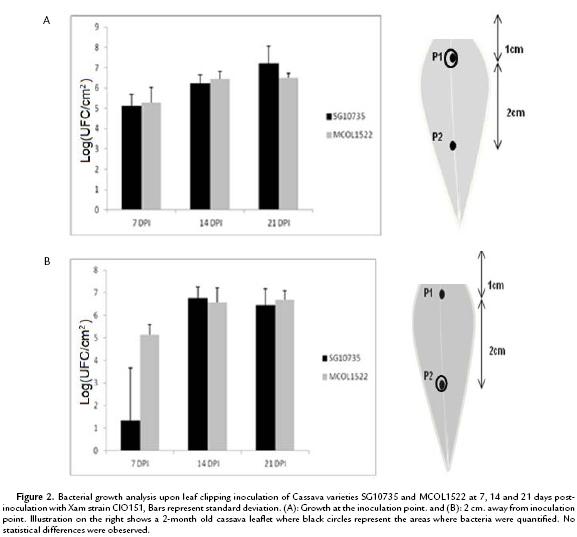
To establish a reliable method with reduced variability to quantify bacterial growth in cassava, we first evaluated leaf-clipping inoculation. As shown in Figure 2A, bacterial numbers at the inoculation point were similar in both cassava varieties MCOL1522 and SG10735, which is in opposition to what was expected from the AUDPC analysis. Inspection of bacterial growth at 2 cm. away from the inoculation point also showed a similar trend in both varieties (Fig. 2B), indicating that there is no differences in bacterial numbers.
Leaf puncturing inoculation
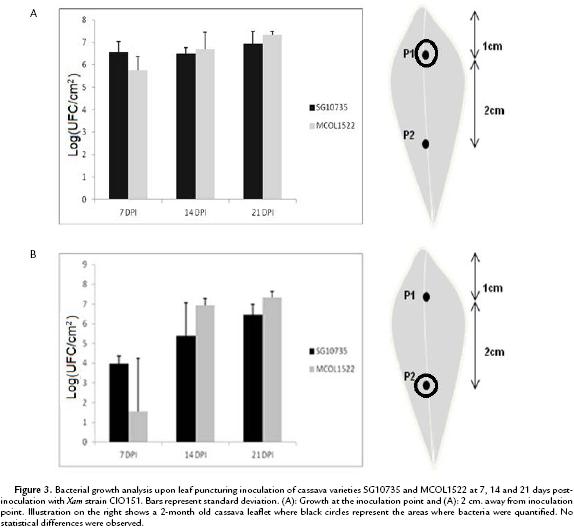
Because Xam is a vascular pathogen, we next investigated the effect of puncturing the main vein of 8 week old cassava leaves. As shown in Fig. 3A, bacterial growth was monitored over time within a 5 mm2 leaf fragment comprising the inoculation point in the main vein. Again, measurements performed at 7, 14 and 21 days post-infection indicated that bacteria multiplied at similar level in both MCOL1522 and SG10735 varieties. Similarly, when bacterial numbers were monitored 2 cms away from the point of inoculation no difference could be observed between the two varieties (Fig. 3B). Hence, neither leaf-clipping nor leaf puncturing inoculation revealed differences in Xam growth within leaves of the two cassava varieties.
Stem inoculation
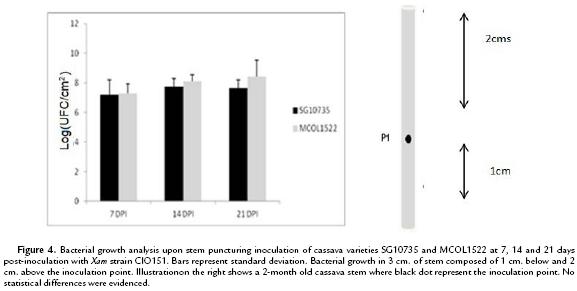
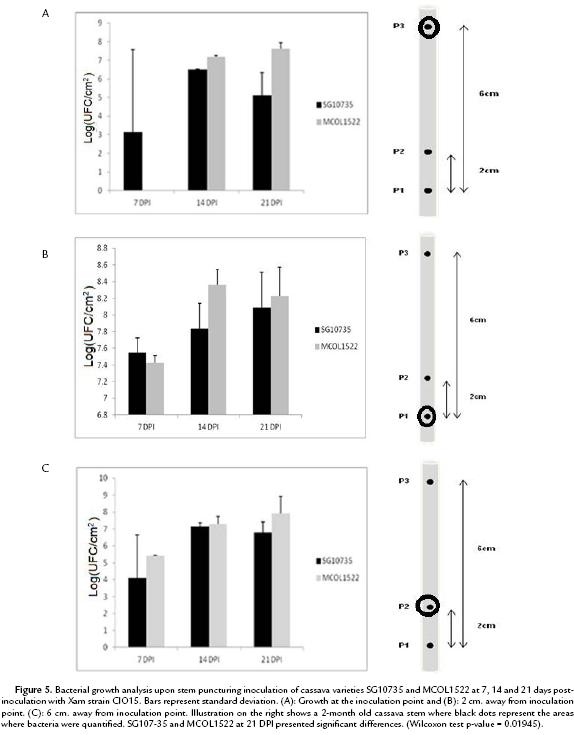
Xam naturally colonizes cassava stems and techniques have been previously established to inoculate stems under laboratory conditions (Restrepo et al., 2000b; Jorge and Verdier, 2002). We therefore evaluated bacterial growth of Xam after stem puncture inoculation. In a first attempt, bacterial growth was monitored at 7, 14 and 21 dpi within a 3 cm long stem fragment including 1 cm below and 2 cm above the inoculation puncture, respectively. As shown in Fig. 4, bacterial number was similar to those observed upon leaf-clip inoculation with no differences observed among varieties. In a second attempt, the same puncture inoculation method was employed but bacterial multiplication was monitored within different stem fragments. Stem fragments of 1 cm-long were collected at the puncture of inoculation, at 2 cm and 6 cm above it. As shown in Fig. 5A and 5B, bacteria grew similarly in both varieties when measurements were performed from the inoculation site and 2 cm away from it. Interestingly, as shown in Fig. 5C, quantification of bacterial at 6 cm above the puncture of inoculation highlighted a 4-log difference at 21 dpi of Xam populations in variety MCOL1522 as compared to SG10735. These differences are significant (W-test at p-value=0.01945) and confirm the results obtained by AUDPC that differentiate between the two cassava varieties.
DISCUSSION
With the advent of cassava and Xam genome sequences it should be possible to identify resistance and pathogenicity candidate genes. The functional validation of both, bacterial and plant genes will require studying their phenotypes when these are overexpressed or mutated. To achieve this objective, a robust system of inoculation to observe the loss or gain on resistance to Xam is required. To establish a method for quantification of bacterial multiplication, we studied different methods previously reported in the literature for cassava and/or other pathosystems (Jorge and Verdier, 2002; Wydra et al., 2004; Meyer et al., 2005 Kim et al., 2008 ).
Stem inoculation is a widely used method to evaluate the resistance/susceptibility of cassava plants and it represents the more natural way of Xam spread in the crop (Wydra et al., 2004). However few studies have quantified the bacteria present in these tissues and the only optimal way to do it is the qualitative AUDPC approach (Jorge and Verdier, 2002). In the present study, we observed quantitative differences in the bacterial numbers between the two varieties used in this study only by taking a portion of stem, 6 cm away from inoculation point. The results obtained with this method were comparable with the AUDPC results. Hence, we argue that the methodology presented here constitutes a sensitive method allowing detection of small resistance/susceptibility differences between varieties. Furthermore, this result suggests that Xam, as a vascular pathogen, moves preferentially across the stem and not as effectively on leaves, and supports the idea of non-regular bacterial aggregation across the tissue but only close to the inoculation point.
Evaluation of symptoms in leaves, as it is established in several pathosystems, would be more economical and practical for testing phenotypes in cassava plants. Nevertheless, probably because of the high content of polysaccharides present in the cell wall of cassava leaves, methods such as infiltration are very difficult and variable between cassava varieties. To do an accurate bacterial quantification, leaf puncturing and leaf clipping methods were evaluated. The results obtained using these methods, resulted in high variability, and lead us to evaluate bacterial movement in plant tissues. Xam is a vascular pathogen that colonizes the xylem (Verdier, 2002). Therefore, the variability reported here, can be explained considering the possibility that the flow of water and other substances through the vascular vessels could increase bacterial mobility. In consequence, bacterial aggregation varies in space and time resulting in sampling variability. Thus, the quantification of the bacteria at distant sections from the inoculation point will be a stochastic phenomenon appropriate for Xam.
Even if a high variability was observed, we did not observe any differences in the cassava varieties after using the inoculation methods on leaves, although by AUDPC, the traditional and standard method to evaluate resistance, the two varieties were different. Previous studies using other cassava varieties indicated that the cassava responses are similar in the resistant and susceptible varieties in particular in the mesophyll, despite not evaluating different regions of the plant or different times post inoculation.
Despite of these facts, we could observed the bacterium was not able to grow in distant regions from the inoculation point in stem. Interestingly, it has been reported that leaf inoculation could be a useful method to evaluate mutant pathogenicity (Castiblanco et al., 2013). Nevertheless based on our results, we argue that this technique is not useful to evaluate resistant and susceptible cassava varieties. Consequently, the mobility assays could be important to compare the growth of pathogenic Xam strains in contrasting varieties. Recent PCR-based strategies have been developed to detect Xanthomonas in planta (Albuquerque et al., 2012; Lang et al., 2014). Further studies could be directed to quantify bacteria employing real time PCR.
CONCLUSIONS
In conclusion and taking together these results, we propose as a quantitative evaluation, complementary to AUDPC, to measure the bacterial growth taking 1 cm of tissue 6 cmaway from the stem inoculation point at 21 dpi.
AKNOLWDGMENTS
W would like to acknowledge Paula Diaz, Andrea Vasquez, Elizabeth Contreras, Andrea del Pilar Barrera, Juan Camilo Ochoa and Ana Maria Bossa from the Manihot Biotech group of the National University from Colombia, for their cooperation allowing the successful obtention of the results generated in this study. We wish to thank Liliana López for giving advices in statistical analysis. We thanks the anonymous reviewer and editor for helpful comments and suggestions to improve the quality of the manuscript. Funding was obtained from the National University from Colombia under the quipu project number 201010016732 and ECOS-Nord project C11A02.
REFERENCES
Albuquerque P, Caridade C, Rodrígues A, Marcal A, Cruz J, Cruz L, et al.. Evolutionary and experimental assessment of novel markers for detection of Xanthomonas euvesicatoria in plant samples. PLoS ONE. 2012;7(5):e37836. Doi: http://dx.doi.org/10.1371/journal.pone.0037836. [ Links ]
Arrieta-Ortíz M, Rodriguez-R L, Pérez-Quintero A, Poulin L, Díaz A, Arias N, et al. Genomic Survey of Pathogenicity Determinants and VNTR Markers in the Cassava Bacterial Pathogen Xanthomonas axonopodis pv. manihotis Strain CIO151. PLoS ONE. 2013;8(11): e79704. Doi: 10.1371/journal.pone.0079704. [ Links ]
Bart R, Cohn M, Kassen A, Mccallum E, Shybut M, Petriello A, et al. High-throughput genomic sequencing of cassava bacterial blight strains identifies conserved effectors to target for durable resistance. PNAS. 2012;129(28):1972-1979. Doi: 10.1073/pnas.1208003109. [ Links ]
Castiblanco L, Gil R, Osorio D, Perez A, Munoz A, Restrepo S, et al. TALE1 from Xanthomonas axonopodispv.manihotis acts as a transcriptional activator in plant cells and is important for pathogenicity in cassava plants. Mol Plant Pathol. 2013;14(1):84-95. Doi: 10.1111/j.1364-3703.2012.00830.x. [ Links ]
FAO. Food Outlook, Global Market Analysis. Cassava. 2011;39-44. [ Links ]
Jorge V, Fregene M, Duque M, Bonierbale M, Tohme J, Verdier V. Genetic mapping of resistance to bacterial blight disease in cassava (Manihot esculenta Crantz). Theor Appl. Genet. 2000;101:865-872. Doi: 10.1007/s001220051554. [ Links ]
Jorge V, Verdier V. Qualitative and quantitative evaluation of cassava bacterial blight resistance in F 1 progeny of a cross between elite cassava clones. Euphytica 2002;123(1): 41-48. Doi: http://dx.doi.org/10.1023/A:1014400823817. [ Links ]
Kim J, Taylor K, Hotson A, Keegan M, Schmelz E A, Mudgett M. XopD SUMO Protease Affects Host Transcription, Promotes Pathogen Growth, and Delays Symptom Development in Xanthomonas-Infected Tomato Leaves. Plant Cell. 2008;20(7):1915-1929. Doi: http://dx.doi.org/10.1105/tpc.108.058529. [ Links ]
Kpémoua K, Boher B, Nicole M, Calatayud P, Geiger J. Cytochemistry of defense responses in cassava infected by Xanthomonas campestris pv. manihotis. Can J Microbiol. 1996;42(11):1131-1143. Doi: 10.1139/m96-145. [ Links ]
Lang J, Langlois P, Nguyen M, Triplett L, Purdie L, Holton T, et al. Sensitive detection of Xanthomonas oryzae Pathovars oryzae and oryzicola by loop-mediated isothermal amplification. Appl Environ Microbiol. 2014; 80(15):4519-30. Doi:10.1128/AEM.00274-14. [ Links ]
López C, Soto M, Restrepo S, Piegu B, Cooke R, Delseny M, et al. Gene expression profile in response to Xanthomonas axonopodis pv. manihotis infection in cassava using a cDNA microarray. Plant Mol Biol. 2005;57(3):393-410. Doi: 10.1007/s11103-004-7819-3. [ Links ]
López C, Bernal A. Cassava Bacterial Blight: Using Genomics for the Elucidation and Management of an Old Problem. Trop Plant Biol. 2012;5(1):117-126. Doi: 10.1007/s12042-011-9092-3. [ Links ]
Lozano J, Laberry R. Screening for resistance to cassava bacterial blight. Plant Dis. 1982; 66(4):316-318. [ Links ]
Melotto M, Underwood W, Koczan J, Nomura K, He S. Plant stomata function in innate immunity against bacterial invasion. Cell. 2006;126(5):969-980. [ Links ]
Meyer D, Lauber E, Roby D, Arlat M, Kroj T. Optimization of pathogenicity assays to study the Arabidopsis thaliana-Xanthomonas campestris pv. campestris pathosystem. Mol. Plant Pathol. 2005;6(3):327-333. Doi: 10.1111/j.1364-3703.2005.00287.x. [ Links ]
Prochnik S, Reddy P, Desany B, Rabinowicz P, Mohiuddin M, Rodriguez F, et al. The Cassava Genome: Current Progress, Future Directions. Trop Plant Biol. 2012;5(1):88-94. Doi: 10.1007/s12042-011-9088-z. [ Links ]
Restrepo S, Duque M, Verdier V. Resistance spectrum of selected Manihot esculenta genotypes under field conditions. Field Crops Res. 2000a; 65(1):69-77. Doi:10.1016/S0378-4290(99)00072-6. [ Links ]
Restrepo S, Duque M, Verdier V. Characterization of pathotypes among isolates of Xanthomonas axonopodis pv. manihotis in Colombia. Plant Pathol. 2000b; 49(6):680-687. Doi: 10.1046/j.1365-3059.2000.00513.x. [ Links ]
Verdier, V. Bacteriosis vascular (o añublo bacteriano) de la yuca causada por Xanthomonas axonopodis pv. manihotis. CIAT (eds.). La yuca en el Tercer Milenio: Sistemas modernos de producción, procesamiento, utilización y comercialización. [online] 2002. [cited 15 October 2013]. Available from: http://www.agrifoodgateway.com/sites/default/files/articles/bacteriosis_vascular.pdf. [ Links ]
Wydra K, Zinsou V, Jorge V, Verdier V. Identification of pathotypes of Xanthomonas axonopodis pv. manihotis in Africa and detection of quantitative trait loci and markers for resistance to bacterial blight of cassava. Phytopathology. 2004; 94(10):1084-1093. [ Links ]
Xu R, Blanvillain S, Feng J, Jiang B, Li X, Wei H Y, et al. AvrACXcc8004, a type III effector with a leucine-rich repeat domain from Xanthomonas campestris pathovar campestris confers avirulence in vascular tissues of Arabidopsis thaliana ecotype Col-0. J Bacteriol. 2007;190(1):343-355. Doi:10.1128/JB.00978-07. [ Links ]
