Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.2 Bogotá May/Aug. 2015
https://doi.org/10.15446/abc.v20n2.46057
Doi: http://dx.doi.org/10.15446/abc.v20n2.46057
Artículo de investigación
SOBREVIVENCIA Y CRECIMIENTO DE PLÁNTULAS DE TRES ESPECIES ARBÓREAS EN ÁREAS DE BOSQUE MONTANO ANDINO DEGRADADAS POR GANADERÍA EN COLOMBIA
Survival and Growth of Tree Seedlings from Three species in Degraded Andean Mountain Forest for Cattle Grazing in Colombia
Deisy Johana MORENO BETANCUR1, Sandra Eugenia CUARTAS HERNÁNDEZ1
1Instituto de Biología, Universidad de Antioquia. Calle 67 nº 53-108. Medellín, Colombia. For correspondence. scuartashdez@gmail.com
Received: 7 October 2014; Returned for revision: 8 November 2014; Accepted: 6 December 2014.
Associate Editor: Susana Raquel Feldman.
Citation / Citar este artículo como: Moreno Betancur DJ, Cuartas Hernández SE. Sobrevivencia y crecimiento de plántulas de tres especies arbóreas en áreas de Bosque Montano Andino degradadas por ganadería en Colombia. Acta biol. Colomb. 2015;20(2):85-100. doi: http://dx.doi.org/10.15446/abc.v20n2.46057
RESUMEN
En la región lechera del centro de Antioquia el paisaje ha sido ampliamente transformado de bosques montanos a pastizales para pastoreo de ganado vacuno, siendo la restauración del paisaje en esta zona una acción primordial que debe ser incorporada en el plan de uso de la tierra. El objetivo general de este estudio fue seleccionar tres especies arbóreas que promuevan el proceso de regeneración de bosques montanos y evaluar el efecto de diferentes estrategias de siembra sobre el establecimiento de las plántulas. Se consideró el efecto del tipo de fertilización, combinación de especies, disponibilidad lumínica y niveles de precipitación sobre la sobrevivencia y crecimiento de las plántulas. Quercus humboldtii, Weinmannia pubescens y Myrsine coriacea fueron las especies seleccionadas por presentar alto índice de valor de importancia ecológica y alta disponibilidad de semillas. La fertilización orgánica favoreció la sobrevivencia de las tres especies evaluadas y el crecimiento para W. pubescens. La combinación de especies no afectó la tasa de crecimiento pero sí la sobrevivencia. Se detectó también mayor sobrevivencia en las plántulas de las tres especies que recibieron sombra en algún momento del día; al contrario, disminuyó en la época de sequía. El éxito en el establecimiento de Q. humboldtii, la tolerancia de W. pubescens a las heladas y la dispersión de semillas de M. coriacea por aves son características que en conjunto convierten a estas especies en promisoras para procesos de restauración ecológica en áreas de bosque montano.
Palabras clave: combinación de especies, fertilización orgánica, índice de valor de importancia, restauración ecológica.
ABSTRACT
Mountain forest landscapes of the central region of Antioquia department have been largely transformed into pastures; therefore, ecological restoration must be included as a primordial strategy. The aim of this study was the selection of three tree species that help to improve natural regeneration of mountain forest and to evaluate the effect of different sowing strategies on the seedling`s establishment. We assessed the effect of fertilization type, combination of species, light availability and monthly precipitation on seedling survival and growth. Myrsine coriacea, Quercus humboldtii, and Weinmannia pubescens were selected on the basis of high importance value index and high seed availability. Survival of the three species was higher when seedlings were fertilized with organic matter. Also, growth was higher for W. pubescens. The combination of species had no effect on the growth rate of the three species, but it affected negatively the survival of Q. humboldtii and W. pubescens growing with M. coriacea. Survival was higher for seedlings of W. pubescens and M. coriacea out of the sun at some moment during the day. Also, survival diminished during drought period. Successful establishment of Q. humboldtii and tolerance of W. pubescens to frost and regrowth after freeze period and seed dispersal of M. coriacea by birds constitute a set of characteristics that make of these species promissory in the ecological restoration process in degraded mountain forest areas.
Keywords: ecological restoration, importance value index, mixed-species, organic fertilization.
INTRODUCCIÓN
Los Andes tropicales constituyen una región con una alta diversidad de hábitats, producto de complejos gradientes espaciales y ambientales. En el mosaico de paisajes de esta región, los bosques son considerados una prioridad de conservación debido a su elevada diversidad biológica, caracterizada por su alto grado de singularidad y de endemismos (Cuesta et al., 2009). Los bosques montanos son ecosistemas frágiles y estratégicos debido a su importante papel para la provisión de servicios ecosistémicos, principalmente los vinculados a la regulación hídrica (Bubb et al., 2004), a la regulación climática regional y a la captura y almacenamiento de carbono (Cuesta et al., 2009; Tejedor et al., 2012). Al mismo tiempo, estos ecosistemas se encuentran en estado de amenaza a nivel global debido a las dinámicas de cambios de cobertura y uso de la tierra (Webster, 1995; Bubb et al., 2004). En la región norte de los Andes en Colombia, el patrón de uso de la tierra ha creado un mosaico de hábitats transformados para la expansión de las actividades agrícolas, las plantaciones forestales, el establecimiento de cultivos ilícitos, la minería, el desarrollo urbano y predominantemente la ganadería (Instituto Agustín Codazi, 2002; Tejedor et al., 2012). De esta forma, la degradación de bosques tiene su origen en una sobreexplotación de los recursos naturales como resultado del crecimiento poblacional y la demanda alimenticia (Meli, 2003).
En algunas situaciones, la regeneración natural del bosque en áreas previamente degradadas es posible cuando existen condiciones favorables que permitan su recuperación sin intervención humana, como por ejemplo la permanencia del banco de semilla, la colonización por especies de plantas pioneras o la dispersión de semillas por aves, entre otras. Este mecanismo no requiere de manipulación ni costo alguno. Sin embargo, en áreas severamente degradadas, especialmente aquellas dedicadas a la cría de ganado vacuno, la regeneración es lenta o nula debido a barreras químicas, físicas o biológicas (Shono et al., 2007). Una de las barreras para el establecimiento exitoso de las plántulas de especies de árboles nativos son las condiciones adversas de los micrositios como resultado del pisoteo, la herbivoría y la compactación del suelo (Ramírez et al., 2003). También la disponibilidad limitada de semillas y su bajo poder germinativo pueden restringir el proceso de regeneración natural (Omeja et al., 2011). Por lo tanto, la restauración ecológica como alternativa para iniciar la regeneración es una estrategia valiosa para la recuperación de la cobertura del suelo y del bosque a través del establecimiento de especies de plantas nativas (Ramírez-Marcial et al., 2006; Ferreira y Santos, 2012). La restauración de ecosistemas forestales en zonas degradadas aspira a simular e incluso acelerar los procesos que dirigen la sucesión secundaria con el objetivo de devolver el ecosistema perturbado a un estado semejante a su condición original, reconstruyendo la estructura y funciones perdidas (Bradshaw, 1987; Society Ecological Restoration, 2004).
La introducción artificial de plántulas o la siembra directa de las semillas de especies nativas puede funcionar como mecanismo para facilitar la regeneración natural, acelerando la acumulación de biomasa y diversidad (Omeja et al., 2011; Ferreira y Santos, 2012), y al mismo tiempo favoreciendo el establecimiento de estas especies cuando compiten con especies exóticas (Lesica y Allendorf, 1999). El éxito del proceso de recuperación de áreas degradadas depende de la selección adecuada de especies (Vargas, 2011) y un modelo de plantación eficiente (Jesus y Rolim, 2005). En el proceso de selección de especies se debe considerar su potencial de crecer en zonas alteradas y su trayectoria sucesional, de manera que permitan recuperar la fertilidad del suelo y parte de la diversidad de plantas y animales (Vásquez et al., 1999). Para aumentar la eficiencia en la plantación es necesario conocer la autoecología de las especies leñosas, desde las condiciones adecuadas de germinación y manejo, hasta la tasa de sobrevivencia y crecimiento a través de gradientes ambientales (Lesica y Allendorf 1999; Ramírez-Marcial et al., 2006). Dichos aspectos ecológicos son pobremente conocidos para gran parte de la vegetación arbórea en los bosques montanos Andinos, y de igual manera se tiene poca información sobre procesos ecológicos básicos que permita predecir las tendencias del cambio sucesional para el diseño de las medidas técnicas pertinentes para la recuperación, conservación y aprovechamiento de los recursos forestales (Gonzáles-Espinosa et al., 2007).
Existe una variedad de factores bióticos y abióticos que determinan el éxito de establecimiento de plántulas en el proceso de restauración, tales como la incidencia lumínica, los niveles de precipitación (Gonzáles-Espinosa et al., 2007), el suplemento de nutrientes al suelo, la cantidad y diversidad de semillas en el suelo (Baker y Berendse, 1999), longevidad de las semillas, disponibilidad de micrositios, las interacciones bióticas como presencia de polinizadores, dispersores, competencia y depredación (Osunkoya et al., 1992) y la presencia de plantas niñeras (Padilla y Pugnaire, 2006; Vargas, 2011), entre otras. Adicionalmente, se ha observado que las plantaciones con una mezcla de especies pueden alcanzar mayor productividad que los monocultivos (Erskine et al., 2006).
Por lo tanto, con el objetivo de conocer las condiciones adecuadas para el establecimiento de tres especies arbóreas nativas, se evaluó el efecto simultáneo del suplemento de nutrientes al suelo, la mezcla de especies vegetales y la incidencia lumínica, sobre la sobrevivencia y crecimiento de estas, simulando la variación en algunas condiciones naturales de los micrositios en los cuales deben establecerse las plántulas. También se evaluó la relación entre la sobrevivencia y el crecimiento con la precipitación.
Para alcanzar el propósito de este estudio se realizó un inventario y análisis de la vegetación en remanentes de bosque de la región, con la finalidad de seleccionar especies con alto valor de importancia en la estructura de dichas comunidades forestales. Se planteó el ensayo de restauración incluyendo la germinación de las semillas, la siembra en campo y el monitoreo de la sobrevivencia y crecimiento de tres especies arbóreas seleccionadas, de acuerdo a lo planteado por Howell et al., (2012).
MATERIALES Y MÉTODOS
Sitio de Estudio
El área de estudio se localiza en el Municipio de Belmira en la cordillera Central en el Departamento de Antioquia (6°36´2´´N, y 75°38´17´´W). Corresponde al altiplano de la región norte. Su topografía es quebrada, con alturas entre los 2500 y 3270 m s.n.m., con una precipitación media anual de 1787 mm distribuidos en dos períodos de lluvia entre los meses de abril, octubre y noviembre. Es una zona de gran riqueza hídrica, allí nacen varios ríos fundamentales para el abastecimiento de agua de la ciudad de Medellín, lo cual la constituye en un área de gran importancia para conservar los remanentes de bosque y restaurar las áreas degradadas por diversas actividades antrópicas. Los bosques dominados por Quercus humboltii (Bonpl) constituyen la vegetación predominante de la región. Estos bosques han sufrido un continuo proceso de fragmentación y sustitución por áreas de cultivo y pasto para ganadería extensiva durante los últimos 60 años, lo cual ha conducido a un acelerado deterioro de los recursos forestales (Corporación Autónoma Regional del Centro de Antioquia, 2009).
El área de siembra de 1,2 ha se localiza en el Predio Montañita perteneciente al Municipio de Belmira, el cual se encuentra bajo la jurisdicción de la Corporación Autónoma Regional del Centro de Antioquia (CORANTIOQUIA) y ha sido señalado como zona de prioridad de conservación y restauración. El predio consiste en un área de potrero en uso hasta pocos meses antes de la siembra de las plántulas.
Selección de especies arbóreas
Para la selección de especies arbóreas nativas se evaluó la composición florística de bosques en la región central del Departamento de Antioquia, específicamente en los Municipios de Santa Rosa de Osos, Donmatías, San Pedro de los Milagros, Entrerríos y Belmira localizados sobre la Cordillera Central. En esta zona a pesar de la intensa transformación del paisaje debido a la intervención antrópica aún permanecen algunos parches de bosque con bajos niveles de perturbación que albergan especies vegetales propias la región.
Para la caracterización de la vegetación presente en los Municipios de estudio se siguió la propuesta metodológica basada en los inventarios RAP (Rapid Assesment Program) propuesta por Gentry (1995). Esta metodología consiste en establecer diez parcelas de 2 × 50 m (0,1 ha), muestreando todos los individuos con un DAP ≥ 2,5 cm. Las muestras botánicas fueron sometidas al proceso de herborización e identificación taxonómica en el Herbario de la Universidad de Antioquia HUA y con la asesoría de botánicos especialistas.
Para tres de los cinco municipios, Don Matías, San Pedro de los Milagros y Belmira, la información de los transectos RAP fue suministrada por la Corporación Autónoma Regional del Centro de Antioquia CORANTIOQUIA, con base en estudios previos (Lázaro y Echeverry, 1998; Robles, 2006). Para los Municipios de Entrerríos y Santa Rosa de Osos, los transectos fueron realizados en este estudio en las Veredas La Ponderosa y La Pontezuela, respectivamente, en el mes de mayo del 2011.
El criterio de selección de especies se basó en el índice de valor de importancia ecológica (Mostacedo y Fredericksen, 2000), debido a que este índice incorpora parámetros ecológicos como la frecuencia, densidad y dominancia relativa de cada especie, que dan cuenta de la relevancia que cada especie tiene en la estructura de la comunidad. Se consideraron en primer lugar aquellas especies con mayor índice de valor de importancia. En segundo lugar, que tuvieran una alta disponibilidad de semillas y fueran reportadas en la literatura como tolerantes a la exposición lumínica en la fase de establecimiento de las plántulas. En tercer lugar, que fueran de importancia ecológica o económica, previendo que en el futuro estas características aumenten el interés de los habitantes locales para su utilización en proyectos locales de restauración.
Germinación de semillas en vivero y trasplante de plántulas
Una vez seleccionadas las tres especies se procedió con la colección de semillas mediante caminatas en los parches de bosque aledaños al área de siembra. Las semillas se colectaron de varios árboles parentales para incorporar variabilidad genética y prevenir los efectos negativos de la endogamia sobre la diversidad genética de las poblaciones a largo plazo. Se sembraron alrededor de 600 semillas de cada especie en bolsas con suelo del bosque colindante y una vez germinadas, las plántulas se mantuvieron en vivero durante seis meses hasta alcanzar una altura promedio de 20 cm para su posterior trasplante al campo. Se registró el porcentaje de germinación para cada especie considerando el número de semillas germinadas después de cinco meses de siembra en invernadero.
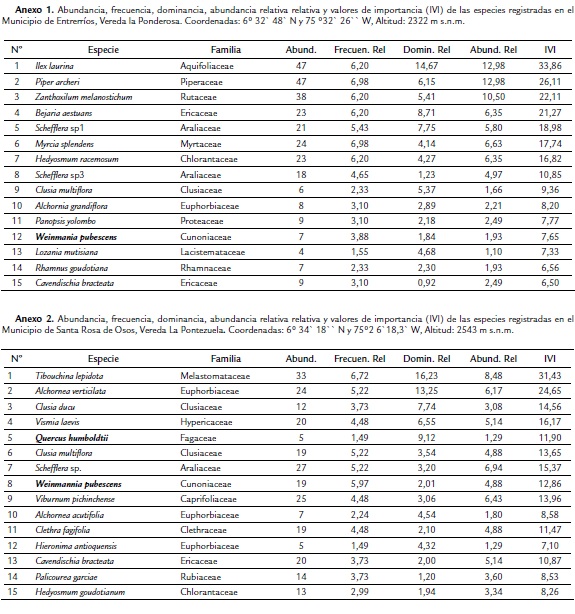
En diciembre de 2011 se trasplantaron 1440 plántulas a la parcela de 1,2 hectáreas, la cual fue dividida en 18 cuadrantes, cada uno con un área de aproximadamente 576 m2. En cada cuadrante se sembraron 80 plántulas, con una separación de 2,0 m siguiendo el método de siembra tresbolillo (Fig. 1A). En este sistema las plantas se siembran formando un triángulo equilátero en el campo, permitiendo sembrar un 15 % más de árboles comparado con el sistema de cuadrado (Centro Agronómico Tropical de Investigación y Enseñanza, 2009), además de permitir simular condiciones más parecidas a la regeneración natural en el sentido que las plantas no crecen ordenadamente formando cuadrados sino que puede darse una distribución aleatoria. En el caso de los cuadrantes en los cuales se sembraron dos o tres especies mezcladas estas se distribuyeron sistemáticamente evitando que individuos de la misma especie estuvieran agrupados.
Efecto de la fertilización, la combinación de especies y la disponibilidad lumínica sobre el establecimiento de plántulas
Se evaluó simultáneamente el efecto de la fertilización, mezcla de especies e incidencia lumínica sobre la sobrevivencia y crecimiento de cada plántula, con el objetivo de simular las condiciones naturales de los micrositios donde las plántulas deben establecerse. Para evaluar el efecto de la combinación de especies se sembraron dos cuadrantes con todas las plántulas de la misma especie (2 cuadrantes × 3 especies = 6 cuadrantes); en otros seis cuadrantes se sembraron plántulas combinadas de dos especies (40 de cada una) y en los seis cuadrantes restantes se sembraron plántulas de las tres especies (27 de cada una) para un total de 480 plántulas por especie (Fig. 1B). Para evaluar el efecto del tipo de fertilización del suelo, nueve de estos cuadrantes (uno con cada combinación de especies) fueron destinados al tratamiento de fertilización química (Q) y los nueve restantes a fertilización orgánica (O).
Para definir de manera adecuada la cantidad de nutrientes necesarios en la fertilización química se realizó un análisis fisicoquímico del suelo y atendiendo a los requerimientos de nutrientes de este se usó la dosis recomendada para los tres macroelementos químicos principales que requieren las plantas, nitrógeno, fósforo y potasio (Bonilla, 2008) con fertilizante NPK 15-15-15 (40 g/planta) y microelementos a partir de fertilizantes menores Rio Claro (25 g/planta).
La fertilización orgánica se realizó con gallinaza (120 g/ planta). Las plántulas fueron marcadas para su monitoreo con placas metálicas cerca de la base del tallo.
Además, se ha observado que las plántulas que crecen a la sombra tienen un mejor desempeño (Gonzáles-Espinosa et al., 2007) y en el área de siembra, la vegetación alrededor de cada cuadrante fue variable (árboles aislados, borde de bosque, etc.) influenciando la cantidad de luz que recibió cada plántula durante el día. De esta forma, se registró la disponibilidad lumínica para cada plántula como una variable categórica con dos niveles, sin sombra (S) para aquellos individuos que crecen totalmente expuestos al sol, y con sombra (C) cuando reciben sombra en algún momento del día.
El éxito del establecimiento se midió a través de la sobrevivencia y el crecimiento para cada uno de los individuos. El número de plántulas sobrevivientes (sobrevivencia) fue registrado cada quince días y la altura de la rama principal (crecimiento) cada mes durante un período de 22 semanas desde el momento de siembra (diciembre de 2011) hasta junio de 2012. Una plántula se consideró muerta cuando sus hojas y tallo estaban secos o habían desaparecido. Los intervalos de tiempo para el monitoreo de sobrevivencia y crecimiento difieren debido a que la sobrevivencia puede presentar cambios en pocos días, mientras que el crecimiento por ser un proceso lento requiere de un período de tiempo mayor que permita observar pequeñas variaciones.
Análisis de datos
En el diseño de siembra se cuenta solamente con una réplica para la combinación de los tratamientos (Especie × Combinación de especies × Tipo de Fertilización), excepto para la combinación de las tres especies para el cual hay tres réplicas. Al mismo tiempo, dentro de cada cuadrante, los datos tomados en cada plántula no son independientes, por lo tanto éstas no constituyen verdaderas réplicas (Hurlbert, 1984; Quinn y Keough, 2002). El uso de réplicas sensu stricto permite estimar el efecto del tratamiento entre unidades experimentales, aumenta la precisión de los estimados en sistemas biológicos que son altamente variables y es ideal en cualquier diseño experimental. No obstante, se reconoce la dificultad de obtener réplicas en experimentos ecológicos, especialmente cuando éstos son a gran escala, debido a que la combinación de grandes escalas espaciales y temporales con la replicación es costosa logística y económicamente (Oksanen, 2001). En este estudio, fue posible evaluar el efecto de los tratamientos a la escala adecuada (576 m2 y n = 180 plántulas en promedio) pero no replicarlos debido a la limitación de espacio en el área de siembra para la dispersión espacial de los tratamientos, la limitación de material vegetal, de capital económico y humano. A pesar de las limitaciones estadísticas debidas a la falta de replicación de los tratamientos, la cual impone una restricción para la aplicación de la estadística inferencial (Hurlbert, 1984), la interpretación ecológica de los datos es factible (Morgan y Lunt, 1999) como aproximación valiosa para el entendimiento del comportamiento del crecimiento y la sobrevivencia en la variedad de condiciones evaluadas.
De esta forma, se evaluó si existen diferencias en la sobrevivencia y la tasa de crecimiento de las plántulas entre especies y para cada especie entre tipo de fertilización, combinación de especies y disponibilidad lumínica, considerando el efecto de cada factor de manera independiente. Para el factor "tipo de fertilización" con dos niveles (orgánica y química) se realizó una prueba t. Para el factor "combinación de especies" con cuatro niveles (sola, con una de las dos especies restantes, con la otra especie o las tres especies) y para la comparación entre especies (tres especies) se utilizó un análisis de varianza. Para el factor disponibilidad lumínica, debido a que se registró después de la siembra de las plántulas, el número de individuos en cada tratamiento (con y sin sombra) fue diferente (i.e., diseño desbalanceado), y además no cumplió el supuesto de homocedasticidad, por lo tanto se aplicó una prueba t de Welch (Zar, 1999). También se realizó un análisis de varianza para evaluar el efecto de la disponibilidad de luz como factor anidado dentro del factor fertilización o combinación de especies sobre la tasa de crecimiento o sobrevivencia. La hipótesis nula evaluada es que no existe ningún efecto de crecer con o sin sombra dentro de cualquier tipo de fertilización o combinación de especies. En el caso de la sobrevivencia que es una variable categórica el ajuste de los dos modelos (i.e., el modelo que incluye el efecto de cualquier interacción –`full model´- o asociación entre las variables y el que no lo incluye –`reduced model´-.) se utilizó el estadístico "likelihood ratio" (Λ∼G2) (Quinn y Keough, 2002).
Aunque en algunos casos las variables no presentan distribución normal, en cada nivel de los factores analizados el tamaño muestral es grande (N>80) lo cual permite asumir un comportamiento normal de las variables y utilizar pruebas paramétricas (Charles y Laurie, 1997). Para los casos en que se encontraron diferencias entre los niveles del factor se realizaron pruebas de Tukey para identificar los grupos homogéneos.
Adicionalmente, las plántulas de cada especie fueron agrupadas dependiendo de los niveles de cada factor asignados, de manera que cada grupo corresponde a una especie en condiciones de fertilización, combinación de especies y disponibilidad de luz particulares. Por ejemplo, uno de los posibles grupos formados puede ser: plántulas de Quercus humboldtii (Q) con fertilización orgánica (O), creciendo en combinación con Myrsine coriacea Sw (M) y sin sombra (S). Este grupo estaría representado por el código QM-O-S. El número total posible de grupos es 48 de los cuales 16 grupos son de cada una de las tres especies (3 especies × 2 tipos de fertilización × 4 tipos de mezcla de especies × 2 tipos de disponibilidad de luz). Sin embargo, como la categoría de recibir o no sombra no derivó de un diseño estadístico sino de las condiciones naturales en campo, se obtuvieron menos grupos del total posible. Las diferencias en tasa de crecimiento entre grupos se evaluaron usando un análisis de varianza. Para evaluar la asociación entre la sobrevivencia y los grupos de plántulas conformados se utilizó una tabla de contingencia. Se usó una prueba "likelihood ratio" (Λ∼G2) para determinar el ajuste del modelo que incluye la interacción entre las variables y el que no lo incluye (Quinn y Keough, 2002).
También se realizaron curvas de sobrevivencia de Kaplan Meier (Arribalzaga, 2007), las cuales permiten analizar el tiempo transcurrido para que ocurra un suceso; en este caso la muerte de las plántulas, considerando además los datos censurados, es decir, cada uno de los individuos que hasta el último momento fueron observados y no habían desarrollado el suceso -permanecieron vivos durante el periodo de muestreo-. Se usó la prueba estadística Log- Rank para comparar curvas de sobrevivencia entre los niveles de cada factor. La tasa de crecimiento se calculó como el crecimiento absoluto mediante la formula (Barrera et al., 2010):.
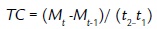
Siendo: Mt: Longitud final; Mt-1: Longitud inicial; t: Número de días.
Todos los análisis fueron realizados usando el paquete estadístico SPSS versión 17 (SPSS, 2007) y las gráficas se construyeron con el Software Sigmaplot versión 12 (Sigmaplot, 2014).
Se realizaron análisis de regresión lineal y cuadrática para observar si existe relación entre los niveles de precipitación promedio con la sobrevivencia promedio en intervalos de tiempo dos semanas y con la tasa de crecimiento promedio de las plántulas en intervalos de tiempo mensuales. Los registros de precipitación mensual para el período de estudio en la localidad fueron suministrados por el Departamento de Calidad Ambiental de la Corporación Autónoma Regional del Centro de Antioquia del Municipio de Belmira.
RESULTADOS
Selección de especies arbóreas
Como resultado de los muestreos florísticos para los Municipios de Entrerríos y Santa Rosa de Osos, se encontró un total de 751 individuos con diámetro igual o superior a 2,5 cm, pertenecientes a 69 especies de 36 familias. En el Municipio de Entrerríos se registraron 362 individuos representados por un mayor número de especies y familias respecto a los encontrados en Santa Rosa de Osos. Para el primero los individuos fueron identificados dentro de 31 familias y 46 especies, mientras los individuos del segundo municipio se clasificaron en 24 familias y 41 especies. En los anexos 1 y 2 se muestran las primeras 15 especies con mayor índice de valor de importancia ecológica encontradas en estos dos municipios.
Con base en la información obtenida del trabajo de campo y la reportada por Lázaro y Echeverry (1998) y Robles (2006) quienes desarrollaron transectos RAP en los Municipios de Belmira, Don Matías y San Pedro de los milagros, se seleccionaron de manera preliminar varias especies con mayor índice de valor de importancia en las cinco localidades. La selección final se realizó con base en información obtenida de la comunidad local sobre una alta disponibilidad de semillas e importancia económica (i.e., maderable o leña). Las especies seleccionadas para la evaluación de su desempeño en campo fueron: Quercus humboldtii, Weinmannia pubescens Kunt y Myrsine coriacea (Fig. 2 A, B y C, respectivamente).

Efecto de la fertilización, la combinación de especies y la disponibilidad lumínica sobre el establecimiento de las plántulas
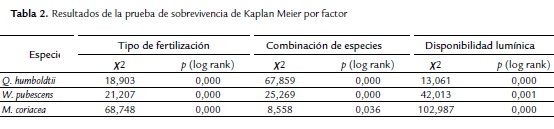
El porcentaje de germinación presentó variación entre especies. La especie con menor porcentaje de germinación fue W. pubescens (26 %), comparado con M. coriacea (62 %) y Q. humboldtii (69 %). Se observó una variación en la tasa de crecimiento (F=114,5; g.l.=2; p=0,000) y sobrevivencia (F =141,0; g.l. =2; p=0,000) entre las especies. Q. humboldtii presentó mayor crecimiento (0,036 cm/día) y sobrevivencia promedio (20,87 semanas), mientras que W. pubescens y M. coriacea presentaron el menor número promedio de semanas sobreviviendo (16,7 y 14,5 semanas, respectivamente) y menor tasa de crecimiento (0,016 y 0,020 cm/día, respectivamente). En general, los individuos permanecieron vivos un mayor número de semanas al fertilizar el suelo de manera orgánica (18,9 semanas) comparado con la fertilización química (17,8 semanas), observándose esta tendencia para las tres especies. Esta tendencia es consistente al evaluar el efecto de la fertilización sobre cada una de las tres especies. La tasa de crecimiento fue significativamente mayor en plántulas de Q. humboldtii con fertilización química, contrario a W. pubescens, la cual tuvo mayor crecimiento con fertilización orgánica. M. coriaceae no presentó diferencias en crecimiento entre los niveles de este factor (Tabla 1).
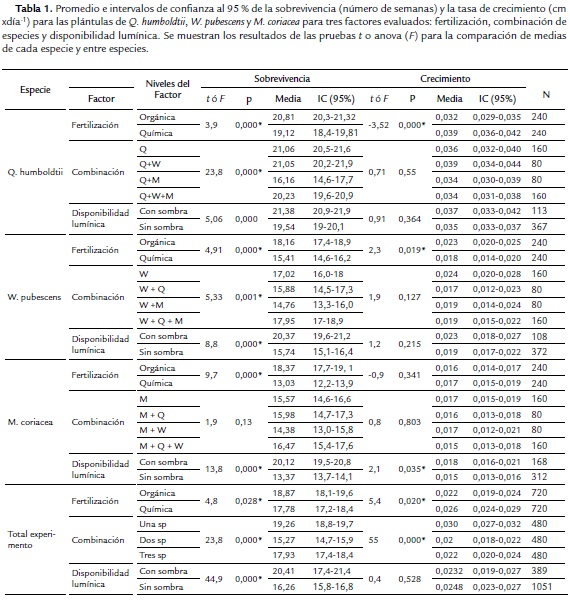
La sobrevivencia de los individuos de Q. humboldtii fue menor en combinación con plántulas de M. coriacea en comparación con las demás combinaciones que no mostraron diferencias entre sí (Prueba de Tukey t = 106,887; p = 0,528). W. pubescens fue afectada negativamente en sobrevivencia al crecer en combinación con M. coriaceae o Q. humboldtii (Prueba de Tukey t = 107,09; p≤0,001). En contraste, M. coriacea no presentó diferencias en sobrevivencia por efecto de las especies vecinas (Prueba de Tukey t =107,42; p=0,092). La tasa de crecimiento no presentó diferencias en ninguna de las especies por efecto de la mezcla de especies (p>0,05).
Recibir sombra en algún momento del día favoreció la sobrevivencia de W. pubescens y Q. humboldtii sin efecto en la tasa de crecimiento. Al mismo tiempo favoreció el crecimiento y sobrevivencia de M. coriacea (Tabla 1).
El análisis de varianza anidado permitió detectar que en general para las tres especies existe un efecto positivo de recibir sombra sobre la sobrevivencia en cualquier tratamiento de fertilización (Q. humboldtii: Likelihood Ratio LR- χ2=15,35; g.l.=2; p=0,0050; W. pubescens: LR- χ2=42,05; g.l.=2; p<0,0001; M. coriacea: LR- χ2=63,39; g.l.=2; p<0,0001) y de combinación de especies (Q. humboldtii: Likelihood Ratio LR- χ2=15,61; g.l =4; p=0,0036; W. pubescens: LR- χ2=35,48; g.l.=4; p<0,0001; M. coriacea: LR- χ2=104,63; g.l.=4; p<0,0001). Sin embargo, el efecto de recibir sombra no fue consistente entre especies en cualquier tratamiento de fertilización (Q. humboldtii: F=1,45; g.l.=2; p=0,2339; W. pubescens: F=0,37; g.l.=2; p<0,6867; M. coriacea: F=4,81; g.l.=2; p<0,0085) o combinación de especies (Q. humboldtii: F=1,21; g.l.=4; p=0,3042; W. pubescens: F=2,86; g.l.=4; p<0,0230; M. coriacea: F=3,62; g.l.= 4; p<0,0064). Específicamente, la tasa de crecimiento de Q. humboldtii no fue afectada por la disponibilidad de luz en ningún tratamiento de fertilización o combinación de especies. En W. pubescens sólo hubo efecto de la disponibilidad de luz dentro de los tratamientos de fertilización y en M. coriacea se detectó efecto de la sombra dentro de los tratamientos de fertilización y combinación de especies.
De los 16 grupos posibles que se pueden conformar de acuerdo al tipo de fertilización recibida, combinación de especies y disponibilidad lumínica experimentadas por cada plántula, se conformaron 14 grupos tanto para Q. humboldtii como para M. coriacea y 15 grupos para W. pubescens. En promedio cada grupo estuvo formado por 34 individuos. El análisis de varianza indica que existen diferencias en la tasa de crecimiento entre los grupos (F=8,53; g.l.=42; p<0,0001). En la figura 3A se observa el promedio para la tasa de crecimiento de cada grupo, observándose en general una tendencia de mayor crecimiento para la especie Q. humboldtii en todos los grupos comparado con M. coriaceae y W. pubescens. En la figura 3B se observa el porcentaje de sobrevivencia para los grupos, en la cual se evidencia que en general las plántulas de Q. humboldtii tienen menor mortalidad que M. coriacea y W. pubescens (Log-likelihood= 258,60; g.l.=42; χ2=517,2; p<0,001). Se observa una tendencia a que las plántulas con fertilización química y creciendo sin sombra (S) presentan alta mortalidad. Específicamente, Q. humboldtii y W. pubescens presentaron baja sobrevivencia en las condiciones anteriores y en presencia de M. coriacea. También se observó que en los grupos con combinación de las tres especies, fertilización química y sombra, W. pubescens y M. coriacea no presentaron mortalidad. En contraste, las plántulas de Q. humboldtii sobrevivieron mejor en combinación con sólo una de las otras dos especies.
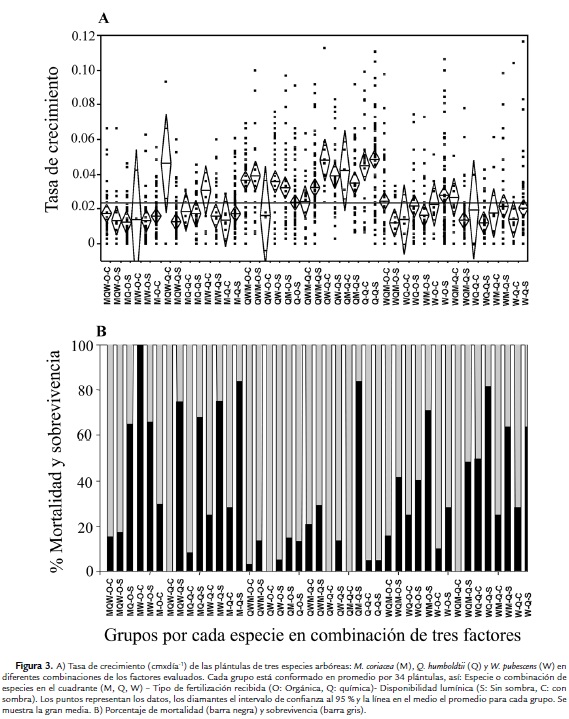
Curvas de sobrevivencia
Las curvas de sobrevivencia obtenidas mostraron diferencias entre los niveles de cada factor evaluado (Tabla 2). Es decir, el porcentaje de sobrevivencia a lo largo del tiempo de seguimiento varió de acuerdo al tipo de fertilización, combinación de especies y disponibilidad lumínica para cada especie (Fig. 4).
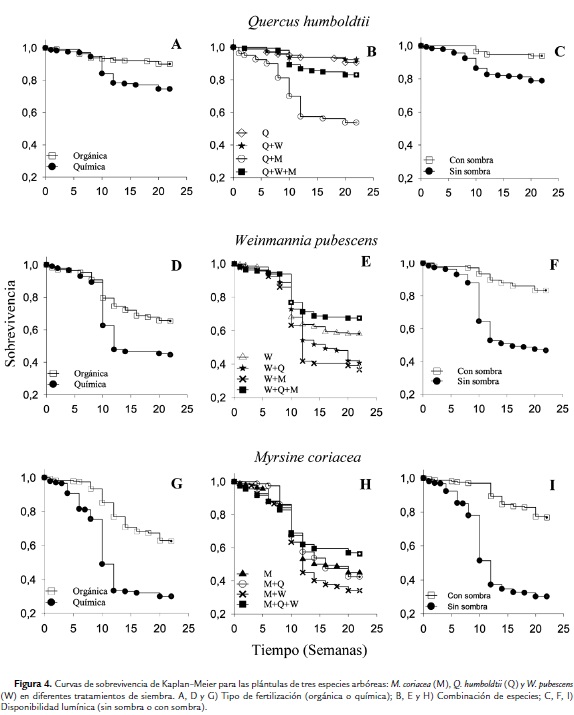
Variación de la mortalidad de acuerdo a los niveles de precipitación
La variación en la mortalidad y la tasa de crecimiento a lo largo del período de monitoreo no estuvo relacionada con la precipitación de manera lineal ni cuadrática. Esta tendencia se observó para las tres especies (p>0,05; 0,1<r2<0,5 para cada caso). Sin embargo, se observa en general, que un aumento en los niveles de precipitación es acompañado de una disminución en mortalidad (Figs. 5A, 5C, 5E) y un aumento en la tasa de crecimiento (Figs. 5B, 5D, 5F). También se observa que el número de individuos muertos aumentó entre las semanas diez y 14, época de mayor sequía.
DISCUSIÓN
Selección de especies arbóreas
La evaluación de la estructura florística de la región posibilitó la identificación de especies clave en el proceso de restauración como componentes fundamentales del ecosistema de referencia. En toda práctica de restauración ecológica la identificación de especies es uno de los puntos de partida crucial para el éxito de ésta, puesto que una elección equivocada de especies en tanto a sus requerimientos ambientales y su hábitat natural puede hacer que los esfuerzos sean insuficientes (Whisenant, 1999).
Las tres especies arbóreas seleccionadas no solo han sido reportadas por su importancia ecológica y socioeconómica (Galindo et al., 2003; Slocum et al., 2004; Solange y Petry, 2009; Avellana y Cárdenas, 2010; Gómez y Vargas, 2011) sino además por su grado de vulnerabilidad al ser parte de ecosistemas que han sido ampliamente degradados y como resultado del uso indiscriminado principalmente como especies maderables y su utilización como leña (Avellana y Cárdenas, 2010; Guerrero et al., 2010). En Colombia, se encuentran bosques dominados principalmente por la especie Q. humboldtii conocidos como robledales (Galindo et al., 2003), y considerados como hábitat de especies vegetales amenazadas y endémicas dentro de las cuales se encuentran especies pertenecientes a los géneros Weinmannia y Myrsine, entre otras. Además de ser el hábitat de una gran diversidad de fauna albergando más de 200 especies de aves, mamíferos y anfibios altamente dependientes de este ecosistema (Avellana y Cárdenas, 2010). La dominancia de Q. humboldtii en bosques andinos se refleja en su alto índice de valor de importancia ecológica en cuatro de los cinco bosques considerados. Por otro lado, W. pubescens se ha considerado como especie clave en procesos de restauración por su potencial de recuperar zonas degradadas o alteradas (Vargas y Gómez, 2011), debido principalmente a su capacidad de crecer en bordes de bosques, caminos e incluso invadiendo potreros (Vargas, 2002). El banco de plántulas de esta especie se encuentra en barrancos y orillas de camino. De igual forma, según Slocum et al., (2004) M. coriacea es una especie pionera importante para la restauración natural de bosques montanos, reportada por su importante asociación con aves. Solange y Petry (2009) encontraron 31 especies de aves que visitaron individuos de esta especie para consumir sus frutos, usarlas como perchas o para forrajear. Debido a su importancia como fuente de alimento y hábitat para las especies de aves es de gran prioridad en proyectos de restauración que permita la recuperación de especies vegetales y además la fauna y procesos de dispersión de semillas asociados a ella.
Efecto de la fertilización, combinación de especies y disponibilidad lumínica sobre el establecimiento de las plántulas
En general la mayor mortalidad de los individuos fertilizados con fertilizante químico para todo el experimento podría estar relacionada de manera indirecta con la competencia del pasto, especialmente de Holcus lanatus, puesto que el aumento de nutrientes suplementados al suelo favoreció su crecimiento. Esta especie es considerada altamente invasora compitiendo fuertemente con las plántulas de otras especies (Díaz et al., 2004; Martínez, 2008). Contrario a esto, en los cuadrantes con fertilización orgánica el crecimiento de pastos y otras malezas fue menor, disminuyendo la competencia con las plántulas sembradas y por lo tanto su mayor sobrevivencia. Esto puede ser debido a que la concentración y solubilidad de los fertilizantes químicos es mayor y fácilmente aprovechable por los pastos, los cuales se encontraron altamente representados en el banco de semillas del área evaluado en un estudio paralelo.
Los resultados presentados en este estudio se basan en la etapa inicial del establecimiento de las plántulas las cuales se encuentran a dos metros de distancia unas de otras, lo cual reduce la posibilidad de que se establezca competencia entre individuos de una misma especie o especies diferentes. Sin embargo, se encontró que la sobrevivencia de las plántulas de Q. humboldtii y W. pubescens es afectada negativamente cuando crecen en combinación con M. coriacea. Adicional a esto, la tasa de crecimiento no presentó diferencias en respuesta a la combinación de especies, a diferencia de lo observado por Cabezas et al., (2008), quienes reportaron menor tasa de crecimiento de Q. humboldtii en el caso de siembra monoespecífica, debido principalmente a que esta se favorece al crecer en mezcla de especies de mayor tasa de crecimiento que faciliten la producción rápida de sombra. Así, la mezcla de especies tiene un efecto indirecto en el crecimiento de plántulas al proveer sombra a estas. El efecto de sombra por las especies usadas en este estudio no se detecta aún debido al tamaño pequeño de las plántulas y a que están separadas dos metros unas de otras, de manera que no se proporcionan beneficio o competencia por luz; sin embargo, se evidencia el efecto positivo de la sombra proporcionado por pocas plantas de otras especies dentro y alrededor del área de siembra sobre la sobrevivencia de las tres especies de estudio.
Por esta razón, una estrategia de restauración que involucre a estas especies debe incluir un sistema que provea sombra necesaria para el establecimiento de las plántulas, como puede ser el uso de plantas niñeras o métodos artificiales utilizando mallasombra. Se ha encontrado que las plantas niñeras proporcionan microhábitat favorables para las plántulas al reducir altas temperaturas de la superficie del suelo e incrementar los nutrientes disponibles, particularmente el nitrógeno (Franco y Novel, 1989; Padilla y Pugnaire, 2006; Ren et al,. 2008). Si bien W. pubescens y M. coriacea han sido reportadas por su capacidad de crecer en áreas abiertas (Vargas, 2002; Lázaro y Vanegas, 2003), los resultados de este estudio sugieren que el suministro de sombra mejoraría su desempeño como especies claves en procesos de restauración. En otros ecosistemas se ha encontrado la facilitación de arbustos pioneros (i.e., plantas nodrizas) sobre las plántulas de especies maderables de estadíos succesionales tardíos como una interacción común en una variedad de ambientes, siendo más acentuada en sitios más soleados y secos (Gómez-Aparicio et al., 2004). En el sitio de estudio, algunos arbustos pioneros de la familia Melastomataceae podrían funcionar como facilitadores, sin embargo, es necesario evaluar este aspecto en el futuro.
Quercus humboldtti ha sido reportada por su capacidad de crecer en diferentes tipos de suelos, desde los medianamente fértiles hasta los degradados y casi estériles (Aguilar, 2009), lo que permite explicar la tolerancia que presenta frente a diversas condiciones edáficas como su alta probabilidad de sobrevivencia tanto para la fertilización orgánica como química comparado con las otras dos especies estudiadas. A diferencia de Ramírez et al., (2003) y Cabezas et al., (2008) quienes sugieren una mayor tasa de crecimiento para la especie al suministrarle sombra a los individuos durante los primeros años de crecimiento, los registros durante las 22 semanas de monitoreo no reflejan un efecto de la sombra sobre el crecimiento pero si en sobrevivencia. No obstante, es probable que las exigencias de la luz solar requerida pueda ser mayor en etapas más avanzadas de crecimiento (Aguilar, 2009).
La especie que presentó mayor porcentaje de mortalidad fue W. pubescens, principalmente en la época de heladas (semanas diez a 14), indicando que es muy susceptible a cambios ambientales como la disminución de la temperatura durante la noche del verano. Sin embargo, se observó que en 20 % de los individuos reportados como muertos pudieron emerger nuevos brotes una vez pasada la época de heladas. Esta fue la especie con mayor dificultad de manejo desde la etapa de germinación hasta el trasplante en campo debido a su baja tasa de sobrevivencia. A pesar de que las especies del género Weinmannia se han reportado como poco exigentes a las condiciones de suelo y luminosidad (Vargas, 2002), W. pubescens tuvo mejor desempeño con fertilización orgánica y sombra.
En un estudio complementario, se evaluaron algunas de las barreras que impiden la restauración de la zona, una de ellas fue que en el banco de plántulas dominan plántula de M. coriacea (75 %), debido posiblemente a que son dispersadas por aves. Estas se alimentan de sus frutos y una vez las semillas pasan por su tracto digestivo son devueltas al medio y germinan fácilmente (Solange y Petry, 2009). Esta observación indica la importancia de la especie en procesos de regeneración de bosques.
CONCLUSIONES
Entre las características que hacen a las especies evaluadas apropiadas para potenciar el proceso de recuperación de áreas degradadas en bosques montanos de zonas Andinas se encuentran la alta sobrevivencia y crecimiento de Q. humboldtii en cualquier combinación de los factores considerados, la estrecha interacción de M. coriacea con aves y la tolerancia de W. pubescens a las heladas. Teniendo en cuenta la problemática ambiental a nivel mundial ocasionada por el uso excesivo de fertilizantes químicos, es de resaltar que este tipo de fertilización solo favoreció el crecimiento de Q. humboldtii y afectó la sobrevivencia de las tres especies de manera negativa, lo que sugiere que es posible potenciar el uso de fertilización orgánica como alternativa de suplemento del suelo en proyectos que impliquen la siembra de estas especies en áreas extensas.
Finalmente, las condiciones recomendadas para iniciar un proceso de restauración en esta zona usando las tres especies evaluadas es realizar la siembra durante la época de mayor precipitación para asegurar una baja mortalidad de plántulas y promover una mayor tasa de crecimiento. De esta forma se favorece la adaptación de las plántulas en campo antes de la temporada de sequía donde ocurre la mayor mortalidad por causa de las heladas. Además, se sugiere el uso de fertilizante orgánico y suministro de sombra debido a que estos favorecen la sobrevivencia de las tres especies al reducir la competencia con los pastos.
AGRADECIMIENTOS
Al Comité para el Desarrollo de la Investigación (CODI) de la Universidad de Antioquia por la financiación del proyecto: "Ensayo de restauración de bosques Andinos en la región lechera del centro de Antioquia" CPT-1004. A la Corporación Autónoma Regional del Centro de Antioquia CORANTIOQUIA por facilitar el predio para la siembra de las plántulas y el apoyo logístico y al cabildo verde de Belmira por su ayuda en el trabajo de campo.
REFERENCIAS
Aguilar M. Plan de restitución poblacional de Quercus humboldtii Bonpl. En la reserva forestal protectora el robledal, Cundinamarca, Colombia. Universidad Alcalá de Henares, Madrid. 2009. p. 1-32. [ Links ]
Arribalzaga E. Interpretación de las curvas de sobrevivencia. Rev Chil Cir. 2007. 59(1):75-83. Doi: http://dx.doi.org/10.4067/S0718-40262007000100013. [ Links ]
Avellana A, Cárdenas L. Conservación y uso sostenible de los bosques de Roble en el corredor de conservación Guantiva- La Rusia- Iguaque, Departamentos de Santander y Boyacá, Colombia. Colomb for. 2010;13(1):5-25. Doi: http://dx.doi.org/10.14483/udistrital.jour.colomb.for.2010.1.a01. [ Links ]
Baker J, Berendse F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends Ecol Evol. 1999;14(2):63-68. Doi: http://dx.doi.org/10.1016/S0169-5347(98)01544-4. [ Links ]
Barrera J, Suárez D, Melgarejo L. Análisis de crecimiento en plantas. In: Experimentos en fisiología vegetal. Melgarejo D, editor. Primera edición. Universidad Nacional de Colombia. 2010. p. 25-38. [ Links ]
Bonilla I. Introducción a la nutrición mineral de las plantas. Los elementos minerales. In: Azcón J, Talón M, editors. Fundamentos de fisiología vegetal. Segunda edición. Barcelona; 2008. p. 103-121. [ Links ]
Bradshaw A. Reclamation of derelict land and the ecology of ecosystems. In: Jordan W, Gilpin M, Aber J, editors. Restoration ecology: a synthetic approach to ecological research. Cambridge University Press, Cambridge, Reino Unido; 1987. p. 53-74. [ Links ]
Bubb P, May I, Miles L, Sayer J. Cloud Forest Agenda. UNEPWCMC. Cambridge, UK; 2004. p. 5-18. [ Links ]
Cabezas M, Peña F, Día C, Moreno A. Dosel de tres especies forestales y su relación con la adaptación a suelos degradados por erosión. Rev udecactualidad divulg cient. 2008; 11(2):175-185. [ Links ]
Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Soluciones para el ambiente y desarrollo. Manejo agronómico de frutales. Imprenta Managua, Nicaragua; 2009. p. 27. [ Links ]
Charles M, Laurie J. Central limit teorema. In: Charles Laurie J, editor. Introduction to probability. American Mathematical Society. United States. 1997. p. 325. [ Links ]
Corporación Autónoma Regional del Centro de Antioquia. Identificación y valoración de los sistemas productivos en el área de manejo especial del sistema de páramos y bosques Altoandinos del Noroccidente Medio Antioqueño con el fin de establecer mecanismos de compensación Medellín. 2009. p. 1-18. [ Links ]
Cuesta F, Peralvo M, Valarezo N. Los Bosques montanos de los Andes tropicales. Una evaluación regional de su estado de conservación y de su vulnerabilidad a efectos del cambio climático. Serie investigación y sistematización. Número 5. Programa regional Ecobona- Intercooperación. Imprenta Mariscal. Quito, Ecuador; 2009. p. 41. [ Links ]
Díaz A, Bonilla M, Vargas O. Competencia entre pastos exóticos y plantas nativas: una estrategia para la restauración del bosque altoandino. Acta biol Colomb. 2004;9(2):103. [ Links ]
Erskine P, Lamb D, Bristow M. Tree species diversity and ecosystem function: can tropical multi-species plantations generate greater productivity?. For Ecol Manage. 2006;233(2-3):205-210. Doi: http://dx.doi.org/10.1016/j.foreco.2006.05.013. [ Links ]
Franco A, Nobel P. Effect of nurse plants on the microhabitat and growth of cacti. J Ecol. 1989;77:870-886. Doi: http://dx.doi.org/10.2307/2260991. [ Links ]
Ferreira R, Santos P. Direct Sowing: An Alternative to the Restoration of Ecosystems of Tropical Forests. In: Sudarshana P, Nageswara-Rao M, Soneji J, editors. Tropical Forests. Brazil: Editorial In Tech. Croacia; 2012. p. 333-348. [ Links ]
Galindo R, Betancur J, Cadena J. Estructura y composición florística de cuatro bosques Andinos del Santuario de Flora y Fauna Guanentá, Alto Río Fonce, Cordillera Oriental Colombiana. Caldasia. 2003;25(2):313-335. [ Links ]
Gentry A. Patterns of diversity and floristic composition in Neotropical Montane Forest. In: Churchill S, editor. Biodiversity and conservation of Neotropical Montane Forest. The New York Botanical Garden, NY. U.S.A. 1995. p. 103- 126. [ Links ]
Gómez P, Vargas O. Grupos funcionales de especies promisoras para la restauración ecológica con base en sus rasgos de historia de vida en la Reserva natural Ibanasca (Ibagué, Tolima, Colombia). In: Vargas O, Reyes SP, editors. La restauración ecológica en la práctica: Memorias del I congreso Colombiano de restauración ecológica. Universidad Nacional de Colombia. Bogotá D.C.; 2011. p. 239-247. [ Links ]
Gómez-Aparicio L, Zamora R, Gómez JM, Hódar JA, Castro J, Baraza E. Applying plant facilitation to forest restauration: A meta-analysis or the use shrubs as nurse plants. Ecol Appl. 2004;14(4):1128-1138. Doi: http://dx.doi.org/10.1890/03-5084. [ Links ]
González-Espinosa M, Ramírez-Marcial N, Camacho-Cruz A, Holtz S, Rey-Benayas J, Parra-Vásquez M. Restauración de bosques en territorios indígenas de Chiapas: Modelos ecológicos y estrategias de acción. Bol Soc Bot Méx. 2007;80:11-23. [ Links ]
Guerrero S, Paz E, Parrado A. Efecto de la intervención antrópica en la distribución de las semillas y plántulas del roble (Quercus humboldtii Bonpl., Fagaceae) en la cordillera oriental colombiana. Colomb For. 2010;13(1):163-180. Doi: http://dx.doi.org/10.14483/udistrital.jour.colomb.for.2010.1.a07. [ Links ]
Howell E, Harrington J, Glass S. Working with people. In: Introduction to Restoration Ecology. Island Press. Washington; 2012. p. 359-372. [ Links ]
Hurlbert, S. Pseudoreplication and the design of ecological field experiments. Ecol Monogr. 1984;54:187-211. Doi: http://dx.doi.org/10.2307/1942661. [ Links ]
Instituto Agustín Codazzi. Zonificación de los conflictos de uso de las tierras en Colombia. En: Coberturas y uso actual de las tierras de Colombia. Bogotá, D.C; 2002. p.84. [ Links ]
Jesus RM, Rolim SG. Fitossociologia da Mata Atlântica de Tabuleiro. Boletim Técnico da Sociedade de Investigações Florestais. 2005;19:1-149. [ Links ]
Lázaro J, Echeverry A. Caracterización de la diversidad vegetal de diez bosques Altoandinos en la jurisdicción de CORANTIOQUIA. Corporación autónoma regional del centro de Antioquia. Medellín, Colombia. 1998. 25 p. [ Links ]
Lázaro J, Vanegas G. 2003. Flora de los Páramos y Bosques Altoandinos del Noroccidente Medio de Antioquia. CORANTIOQUIA. Primera edición. Medellín. 180 p. [ Links ]
Lesica P, Allendorf F. Ecological genetics and the restoration of plant communities: Mix or match? Restor Ecol. 1999;7:42-50. [ Links ]
Martínez M. Holcus lanatus. Revista Plan Agropecuario. 2008;125:45-55. [ Links ]
Meli P. Restauración ecológica de bosques tropicales. Veinte años de investigación académica. Interciencia. 2003;28(10):581-589. [ Links ]
Morgan JW, Lunt ID. Effects of time-since-fire on the tussock dynamics of a dominant grass (Themeda triandra) in a temperate Australian grassland. Biol Conserv. 1999;88:379-386. [ Links ]
Mostacedo B, Fredericksen T. Manual de Métodos Básicos de Muestreo y análisis en Ecología vegetal. Editorial el Pais, Santa Cruz de la Sierra, Bolivia. 2000. p. 82. [ Links ]
Oksanen L. Logic of experiments in ecology: is pseudoreplication a pseudoissue? Oikos. 2001;94:27-38. [ Links ]
Omeja P, Chapman C, Obua J, Lwanga J, Jacob A, Wanyama F, Mugenyi R. Intensive tree planting facilitates tropical forest biodiversity and biomass accumulation in Kibale National Park, Uganda. For Ecol Manage. 2011;261:703-709. Doi: 10.1016/j.foreco.2010.11.029. [ Links ]
Osunkoya O, Ash J, Hopkins M, Graham W. Factors affecting survival of tree sedlings in North Queensland rainforest. Oecologia. 1992;91:569-578. [ Links ]
Padilla F, Pugnaire F. The role of nurse plants in the restoration of degraded environments. Front Ecol Environ. 2006;4 (4):196-202. Doi: 10.1890/1540-9295(2006)004[0196:TRONPI]2.0.CO;2. [ Links ]
Quinn G, Keough M. Experimental desing and data analysis for Biologists. United Kingdom at the University Press, Cambridge. 2002. p. 393-399. [ Links ]
Ramírez J, Zapata C, León J. Descomposición de hojarasca en bosques montanos naturales de Quercus humboldtii Bompland y reforestados (Pinus patula Schltdl. & Cham. y Cupressus lusitanica Mill.) de la región de Piedras Blancas, Antioquia (Colombia). In: Díez MC, León JD, editors. Primer Simposio Forestal DCF 2003. Universidad Nacional de Colombia. Medellín; 2003. p. 29-35. [ Links ]
Ramírez-Marcial N. Survival and growth of tree seedlings in anthropogenically disturbed Mexican montane rain forests. J Veg Sci. 2003;14:881-890. [ Links ]
Ramírez-Marcial N, Camacho-Cruz A, Gonzáles-Espinosa M, López-Barrera F. Establishment, survival and growth of tree seedlings under successional montane oak forest in Chiapas, Mexico. In: Kappelle M, editors. Ecology and conservation of Neotropical montane oak forests. Ecol Stud. 2006;185:177-189. [ Links ]
Ren H, Yang L, Liu N. Nurse plant theory and its application in ecological restoration in lower subtropics of China. Prog Nat Sci. 2008;18:137-142. [ Links ]
Robles C. Caracterización de la diversidad y uso de la flora silvestre en el municipio de Donmatías-Antioquia. Corporación autónoma regional del centro de Antioquia. Medellín, Colombia; 2006. 76 p. [ Links ]
Society for Ecological Restoration (SER). Science and Policy Working group. The SER International Primer on Ecological Restoration. Washington D.C.; 2004:1-15. [ Links ]
Shono K, Cadaweng E, Durst P. Application of assisted natural regeneration to restore degraded tropical forestlands. Restor Ecol. 2007;15 (4):620-626. Doi: 10.1111/j.1526-100X.2007.00274.x. [ Links ]
Sigmaplot. Exact graps and data analysis. Versión12.0. Systat Sofware Inc. San Jose, USA. 2014. [ Links ]
Slocum M, Aide T, Zimmerman J, Navarro L. Natural regeneration of subtropical montane forest after clearing fern thickets in the Dominican Republic. J Trop Ecol. 2004;20: 483-486. [ Links ]
Solange E, Petry V. Frugivory by birds in Myrsine coriacea (Myrsinacea) inhabiting fragments of mixed Araucaria Forest in the Aparados da Serra Natural Park, RS, Brazil. Rev Bras Ornit. 2009;17(2):113-120. [ Links ]
SPSS Inc. Released. SPSS for Windows, Versión 16.0. Chicago, SPSS Inc. 2007. [ Links ]
Tejedor N, Álvarez E, Arango S, Araujo A, Blundo C, Boza T, et al. Evaluación del estado de conservación de los bosques montanos en los Andes tropicales. Ecosistemas. 2012;21(1-2):148-166. [ Links ]
Vargas W. Guía ilustrada de las plantas de las montañas del Quindío y los Andes Centrales. Manizales: Universidad de Caldas, Centro Editorial; 2002. p. 253- 256. [ Links ]
Vázquez C, Batis A, Alcocer S, Gual M, Sánchez C. Árboles y arbustos nativos potencialmente valiosos para la restauración ecológica y la reforestación, Proyecto J-084. Instituto de Ecología, Universidad Autónoma de México. 1999. 15 p. [ Links ]
Vargas O. Restauración Ecológica: Biodiversidad y Conservación. Acta biol Colomb. 2011;16(2):221-246. [ Links ]
Vargas J, Gómez P. Grupos funcionales de especies promisoras para la restauración ecológica con base en sus rasgos de historia de vida en la Reserva natural Ibanasca (Ibagué, Tolima, Colombia). In: la restauración ecológica en la práctica: Memorias del I congreso Colombiano de Restauración Ecológica. Universidad Nacional de Colombia. Bogotá; 2011. p. 239-247. [ Links ]
Webster GL. The panorama of neotropical cloud forest. En: Churchill SP, Balslev H, Forero E, Luteyn J. Biodiversity and conservation of neotropical montane forest. New York Botanical Garden; 1995. p. 53-57. [ Links ]
Whisenant S. Repairing damaged wildlands: a process- orientated, landscape- scale approach. Cambridge, New York. 1999. p.128- 167. [ Links ]
Zar JH. Bioestatistical analisys. Cuarta edición. Prentice Hall, apper saddle river. New Jersey USA. 1999. p.127. [ Links ]