










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.3 Bogotá Sept./Dec. 2015
https://doi.org/10.15446/Abc.V20n3.42545
Doi: http://dx.doi.org/10.15446/Abc.V20n3.42545.
Artículo de investigación/Original research paper
COMPOSICIÓN VEGETAL, PREFERENCIAS ALIMENTICIAS Y ABUNDANCIA DE BIBLIDINAE (LEPIDOPTERA: NYMPHALIDAE) EN UN FRAGMENTO DE BOSQUE SECO TROPICAL EN EL DEPARTAMENTO DEL ATLÁNTICO, COLOMBIA
Plant Composition, Feeding Preferences and Abundance of Biblidinae (Lepidoptera: Nymphalidae) in a Tropical Dry Forest Fragment in the Department of Atlántico, Colombia
María Angélica VARGAS- ZAPATA1,2, Carlos José BOOM- URUETA1,2, Leidys Isabel SEÑA-RAMOS2, Alba Lucia ECHEVERRY-IGLESIAS2, Neis José MARTÍNEZ HERNÁNDEZ3.
1 Estudiantes de Maestría en Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Km 7-Vía Puerto Colombia, Barranquilla, Colombia.
2 Semillero Investigación de Artrópodos e Insectos (NEOPTERA) del Caribe colombiano, Grupo Biodiversidad del Caribe colombiano. Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Km 7-Vía Puerto Colombia, Barranquilla, Colombia.
3 Grupo de Investigación Biodiversidad del Caribe colombiano, Programa de Biología, Facultad de Ciencias Básicas, Universidad del Atlántico. Km 7-Vía Puerto Colombia, Barranquilla, Colombia.
For correspondence. neyjosemartinez@gmail.com.
Received: 12th March 2014, Returned for revision: 10th May 2014, Accepted: 9th January 2015.
Associate Editor: Víctor López Martínez.
Citation/ citar este artículo como: Vargas-Zapata MA, Boom-Urueta CJ, Seña-Ramos LI, Echeverry-Iglesias AL, Martínez Hernández NJ. Composición vegetal, preferencias alimenticias y abundancia de Biblidinae (Lepidoptera: Nymphalidae) en un fragmento de bosque seco tropical en el departamento del Atlántico, Colombia. Acta biol. Colomb. 2015;20(3):79-92. doi: http://dx.doi.org/10.15446/Abc.V20n3.42545.
RESUMEN
Se analizó la variación espacio-temporal de la abundancia de las mariposas de la subfamilia Biblidinae (Lepidoptera: Nymphalidae) en un fragmento de Bs-T en la Reserva Campesina La Montaña (RCM), Atlántico, Colombia; desde enero hasta agosto de 2011. Se marcaron cuatro puntos dentro del área de estudio, donde se ubicaron trampas Van Someren-Rydon cebadas con calamar en descomposición, fruta fermentada y con una mezcla de los anteriores cebos. Adicionalmente, se realizó una caracterización de la vegetación por punto, para lo cual se tomaron datos de diámetro a la altura del pecho (DAP), altura y tamaño de la copa para todas aquellas plantas leñosas con DAP mayor o igual a 2,5 cm. Se capturaron 76 individuos agrupados en seis especies y cuatro géneros; destacándose Hamadryas februa (Hübner) como la más dominante con 32 individuos. El mes de marzo presentó la mayor riqueza y abundancia (6 especies y 25 individuos) durantes las primeras lluvias en la zona. El punto 3 presentó los valores más altos de diversidad y abundancia de Biblidinae (5 especies y 37 individuos) y la mayor densidad de árboles (D= 0,28 individuos/m2). Se demuestra que la estructura de este grupo de mariposas presenta un patrón temporal y espacial en esta reserva. El análisis de componentes principales demostró que el área basal total (ABT) y la Densidad (D) de plantas leñosas, pueden considerarse como un factor determinante en la distribución y abundancia de las especies de la subfamilia Biblidinae en la RCM.
Palabras clave: abundancia, Biblidinae, densidad, Hamadryas februa, Van Someren-Rydon.
ABSTRACT
The abundance and spatio-temporal variation of butterflies of the Biblidinae subfamily (Lepidoptera: Nymphalidae) in a fragment of Tropical dry forest at the Reserva Campesina La Montaña (RCM), Atlántico, Colombia; from January until August 2011, was analyzed. Within the study area four points were marked; Van Someren-Rydon traps were placed.in each point using rotten squid, fermented fruit and a mixture of both as bait. In addition, a characterization of the vegetation by point was performed, taking data of the diameter to the breast height (DBH), height and size of treetop of all woody plants with a DBH greater than or equal to 2.5 cm. In total 76 butterfly individuals grouped in six species and four genera were captured; being Hamadryas februa (Hübner) the most dominant with 32 individuals. The greatest richness and abundance was recorded in March (6 species and 25 individuals), during the first rains in the study area. The point 3 presented the highest values of diversity and abundance (five species and 37 individuals) of Biblidinae and the higher density of trees (D= 0,28 individuals/m2). The structure of this butterflies group presents a spatio-temporal pattern in this reserve. The principal components analysis showed that the total basal area (ABT) and the density (D) of woody plants, can be considered a determining factor in the distribution and abundance of species of the subfamily Biblidinae at the RCM.
Keywords: abundance, Biblidinae, density, Hamadryas februa, Van Someren - Rydon.
INTRODUCCIÓN
El Bosque Seco Tropical (Bs-T) es uno de los ecosistemas más amenazados en Colombia. Los remanentes con el mayor grado de perturbación se localizan en el Caribe y los valles interandinos (Cauca y Magdalena), donde los procesos de intervención como la expansión de las actividades agropecuarias y el aumento de la densidad poblacional han originado el reemplazo de la cobertura vegetal, dejando remanentes aislados que pueden perder su viabilidad para mantener sus componentes y procesos biológicos (Álvarez et al., 1998). Debido a esta problemática, se hace necesario determinar el estado de conservación de estas áreas, así como los efectos que causa el deterioro de estos bosques en las poblaciones naturales, esto puede lograrse a través de la identificación de especies que reflejen el grado de conservación de estos ecosistemas, tales como las mariposas (Papilionoidea), las cuales son sensibles a los procesos de perturbación (Andrade-C, 1998; Brown Jr y Freitas, 2002). Dentro de este grupo de mariposas, las larvas de especies de Biblidinae se alimentan de plantas de Euphorbiaceae, Moraceae, Sapindaceae, Tiliaceae, Ulmaceae y Urticaceae, las cuales se encuentran en los Bs-T (Díaz, 2006). Los adultos de estas mariposas presentan hábitos alimenticios frugívoros y algunas especies liban sobre el suelo húmedo en busca de sales (Erazo y González, 2008). Estas mariposas son frecuentes en las áreas boscosas húmedas y en los bosques secos, estimándose para el Neotropico más de 270 especies (Erazo y González, 2008).
Sin embargo, las comunidades de biblidínos y las condiciones en que habitan han sido poco estudiadas en los Bs-T en el Caribe colombiano. Algunas caracterizaciones ecológicas e inventarios realizados son los trabajos de Campos-Salazar et al. (2011), quienes registraron ocho especies de esta subfamilia en áreas circundantes a ciénagas en el departamento de Córdoba; Vargas et al. (2011) reportaron en un fragmento de Bs-T en la Reserva Natural Las Delicias (Magdalena, Colombia) las subfamilias Biblidinae y Nymphalinae con diez especies cada una. Para los fragmentos de Bs-T en el departamento del Atlántico, Montero et al. (2009) encontraron 13 especies y Callicore pitheas (Latreille) fue la más abundante; mientras que Prince et al. (2011) reportaron nueve especies de biblidínos en dos fragmentos de Bs-T en Corrales de San Luis en este mismo departamento, donde Hamadryas februa (Hübner) fue la especie más común.
Sin embargo, no existe información sobre la variación espacio-temporal de la abundancia de las mariposas de la subfamilia Biblidinae en los fragmentos de Bs-T en el departamento del Atlántico y sus relaciones con los parámetros de la vegetación, como la densidad y área basal de plantas leñosas. Adicional a lo anterior, en los fragmentos de bosque seco del Caribe colombiano no se ha documentado sobre la preferencia por un tipo de recurso en este grupo de mariposas, razón por la cual se hace necesario evaluar varios tipos de cebos para este grupo de mariposas. Con el propósito de aportar a esta discusión, se evaluó la variación espacio-temporal de la abundancia de las especies de esta subfamilia en la Reserva Campesina la Montaña (RCM).
MATERIALES Y MÉTODOS
Área de estudio
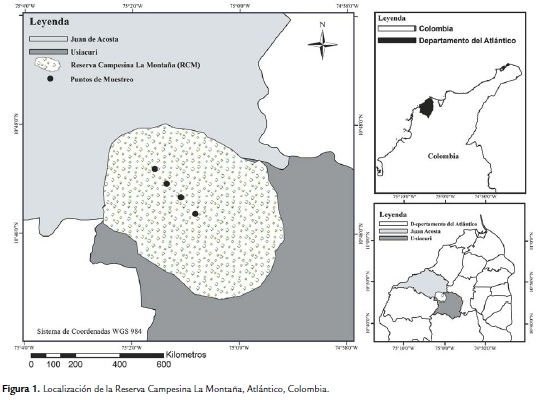
La Reserva Campesina La Montaña (RCM) está ubicada en el municipio de Juan de Acosta, Atlántico, Colombia (Fig. 1) a 10°46'02,6''N y 75°02'34"O; a una altitud entre los 150 y los 260 m.s.n.m. Esta Reserva presenta un fragmento de bosque seco con una extensión de 47 Ha y está rodeado de cultivos permanentes de maíz y yuca, así como de áreas ganaderas de tipo extensiva. La formación vegetal hace parte del Bosque Seco Tropical (Bs-T), según la clasificación de Holdridge (1978) y se encuentra dentro del distrito de los Montes de María y Piojó, localizado en la región biogeográfica del cinturón árido pericaribeño (Martínez et al., 2010). El suelo es de tipo arcilloso-arenoso, con un relieve ondulado que hace parte del sistema de serranías con pendiente entre 12 y 25 %, con grado de erosión moderado (Panza et al., 2001) y el área presenta elevaciones hasta los 238 m. Los meses de mayor precipitación histórica corresponden a agosto y septiembre, con rangos que van desde 100 mm/mes hasta 580 mm/mes. Enero, febrero y marzo corresponden a los meses con menor precipitación, con valores entre 0 mm/mes hasta 30,0 mm/mes. La temperatura media promedio es de 27,6°C, con pocas variaciones a lo largo del año (Gutiérrez y Borjas, 2006).
Dentro del bosque se pueden apreciar los estratos herbáceo, arbustivo y arbóreo. El bosque presenta una altura entre 20 y 25 m de altura y un dosel continuo hacia el borde de los arroyos, lugar donde se pueden encontrar los árboles con mayor altura y en el sotobosque se pueden presentar lianas y arbustos. La vegetación característica del área presenta pocos musgos y helechos, así como aráceas de los géneros Philodendron y Anthurium, característicos de los arroyos en bosques secos. Son abundantes el jobo (Spondias mombin L.), ceiba blanca (Hura crepitans L), indio en cuero (Bursera simaruba L), majagua (Pseudobombax septenatum Jacq.), camajón (Sterculia apetala Jacq.), carito (Enterolobium cyclocarpumJacq.), carreto (Aspidospermapolineuron Müll. Arg), macondo (Cavanillesia platanifolia Bonpl), balso (Ochroma pyramidale Cav. ex Lam). En el estrato arbustivo las especies más dominantes son Capparis baducca (L.), C. odoratissima (Jacq), y C. indica (L). (Martínez et al., 2010).
Muestreo de biblidínos
Los muestreos se realizaron desde enero hasta agosto de 2011, llevándose a cabo dos faenas de campo, para un total de 16 muestreos. Lo anterior, con el fin de tener más información sobre la variación entre muestreos realizados entre la época seca y cuando aparecen las primeras lluvias (abril) y las precipitaciones más fuertes (junio hasta agosto) y sobre los datos de recaptura. En el interior del fragmento localizado en la RCM se delimitó un transecto lineal de 600 m, donde se marcaron cuatro puntos distanciados 150 m entre cada uno. El primero se estableció en el extremo sur del bosque y el cuarto en el extremo norte, siendo estos dos los más cercanos a los bordes; el segundo y tercer punto se ubicaron en el centro del bosque. En cada punto se marcó una parcela con un área de 30 m de largo por 30 de ancho, en la cual se ubicaron en posición triangular y equidistante tres trampas Van Someren-Rydon (VSR) a una altura promedio de 8 m, para un total de 12 por transecto. Por punto, una de las tres VSR fue cebada con 115 g de calamar en descomposición, otra con 115 g de fruta fermentada (papaya, mango y plátano maduro) y la tercera con una mezcla de 115 g de los anteriores cebos. Las trampas se instalaban el día anterior del muestreo, se cebaban el día del muestreo a las 06:00 h y permanecieron en campo 12 horas (hasta las 18:00 h), con cuatro revisiones para cada una (09:00, 12:00, 15:00 y 18:00 h). Las mariposas de la subfamilia Biblidinae, capturadas en cada trampa se sacrificaron siguiendo las recomendaciones descritas por Camero y Calderón (2007), para su posterior identificación en el laboratorio (Permiso 00000934, Corporación Regional Autónoma del Atlántico-CRA). Aquellas mariposas de fácil identificación fueron marcadas teniendo en cuenta las especificaciones descritas por Kingsolver (1999) y Prieto et al., (2005) y de esta manera evitar contar nuevamente los individuos registrados una vez que fueran recapturados. Los ejemplares recolectados se identificaron a través de las claves taxonómicas de Chacón y Montero (2007) y la pagina ilustrada de Butterflies of América (Warren et al., 2012). Los especímenes se encuentran depositados en la Colección Entomológica del Museo de la Universidad del Atlántico (UARC- Números de acceso 382-387).
En cada parcela se midieron las variables ambientales como temperatura ambiente (°C) y humedad relativa (%) cada 30 min con un termohigrómetro (Extech Model RHT 10.USA), tomándose tres mediciones para obtener promedios y contrarrestar las posibles variaciones en cada parcela. La precipitación se midió mediante un pluviómetro (Hellman- Alemania) ubicado en el borde del fragmento.
Con respecto a la vegetación, se realizó una caracterización mediante perfiles vegetales; para ello se levantaron cuatro parcelas, cada una ubicada en un punto de muestreo de mariposas, siguiendo el protocolo propuesto por Villarreal et al. (2006). Cada una constó de un área de 100 m2, distribuida en un transecto lineal de 2 m por 50 m. Se tomaron datos de diámetro a la altura del pecho (DAP), altura y tamaño de la copa para todas aquellas plantas leñosas con DAP mayor o igual a 2,5 cm. Con el fin de tener información que permitiera ilustrar el estrato herbáceo, dentro del transecto fueron censadas totas aquellas plantas con DAP menor a 2,5 cm que tuvieran altura superior a 100 cm (Vázquez y Givnish, 1998).
Se determinó la densidad de las plantas leñosas con DAP superior a los 2,5 cm, de acuerdo al concepto de densidad propuesto por Mostacedo y Fredericksen (2000). La densidad (D) es el número de individuos (N) en un área (A) determinada, que en este caso se tomó al número de individuos de plantas leñosas con DAP superior 2,5 cm como una clase de plantas. La ecuación que se utilizó fue:
D = N/A.
Mientras que el área basal en metros cuadrados se calculó utilizando la fórmula propuesta por Newton (2007):
AB = 0,00008754(DAP)2
El AB de todos los individuos que cumplieron con las condiciones de censo fueron sumados obteniendo el AB total (ABT) para cada parcela.
Análisis de datos
Se determinó el número de especies e individuos capturados en los cebos de fruta fermentada, calamar en descomposición y mixto, así como la variación en el número de especies y la abundancia de mariposas Biblidinae por muestreo, punto y cebo. La similitud entre muestreos fue comparada mediante análisis de agrupamiento UPGMA (Unweighted PairGroup Average), utilizando como medida de similitud el índice de Bray-Curtis, que considera el valor cuantitativo del dato y es asimétrico por no incluir los pares de ausencias en la similitud (Zuur et al., 2007). Para este análisis, los datos de abundancia fueron transformados a logn (x+1) con el fin de contrarrestarle peso a las especies dominantes, pero sin disminuir su importancia. Estos análisis fueron realizados con la ayuda del programa PRIMER-E 6.0 (Clarke y Gorley, 2006). También se determinó la variación espacial en la composición y abundancia de acuerdo al punto de muestro teniendo en cuenta el número de especies y abundancia de éstas.
Con el fin de determinar si se presentaron diferencias en la abundancia entre los puntos y cebos, se aplicó la técnica de ANOSIM (análisis de similaridad) con diseño de una vía. Por otra parte, al considerar a las especies que aportaron más del 10 % a la abundancia total, se determinó qué especies tipifican o caracterizan los puntos de muestreo y los cebos mediante una rutina SIMPER (PRIMER 6.0) (Clarke y Gorley, 2006). Con el fin de conocer qué variables de la vegetación explican mejor la abundancia de Biblidinae en cada punto, se realizó un Análisis de Componentes Principales (ACP). Los indicadores que se utilizaron para este análisis fueron área basal total (ABT), densidad total de árboles (D), puntos de muestreos y la abundancia de las especies de Biblidinae.
RESULTADOS
Variación temporal de la abundancia de mariposas Biblidinae por puntos y muestreo
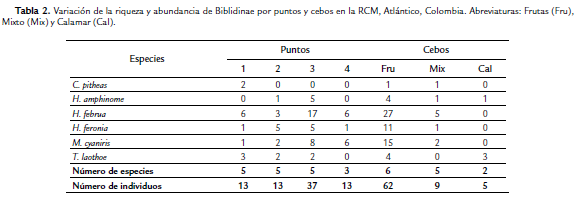
Se capturaron 76 individuos que corresponden a seis especies de cuatro géneros: Callicore pitheas (Latreille), Hamadryas amphinome (Linnaeus), H. februa (Hübner), H. feronia (Linnaeus), Myscelia cyaniris (Doubleday) y Temenis laothoe (Cramer); de las cuales la que presentó la mayor abundancia fue H. februa con 32 individuos y la de menor fue C. pitheas con sólo dos individuos (Tabla 1). Con respecto a los puntos de muestreo, en el 3 se presentó el valor más alto de individuos (37), mientras que el punto 4 presentó la menor riqueza con sólo tres especies. La especie H. februa fue la más abundante en el punto 3, mientras que en el 2 fue H. feronia con cinco individuos (Tabla 2).
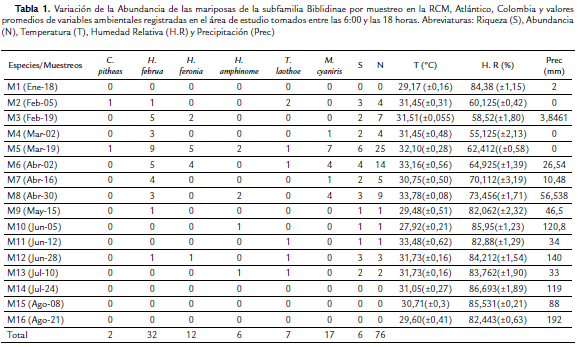
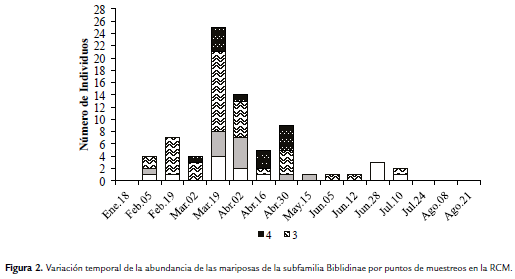
De acuerdo a la variación temporal (meses secos y con lluvias) de la abundancia por puntos, el mayor número de capturas se registró en el punto 3 durante el quinto muestreo (M5), que corresponde a la época seca en el mes de Marzo. Este punto se caracterizó por ser el más abundante entre el M3 y M6 (Tabla 1, Fig. 2). En los muestreos M9 (May-15), M10 (Jun-5), M11 (Jun-12) y M12 (Jun-28), sólo se capturaron individuos en un punto y únicamente en los muestreos M5 y M6 se capturaron individuos en todos los puntos. En el punto 4, se registraron capturas sólo entre los muestreos M4 (Mar-2) hasta el M8 (Abr-30), presentando el mayor número de capturas en el quinto y octavo muestreo (Tabla 1, Fig. 2).
De acuerdo a los muestreos, el que presentó la mayor riqueza y abundancia fue el quinto (Mar-19) con seis especies y 25 individuos; con valores de temperatura (T), Humedad Relativa (H.R) y Precipitación (Prec.) de 32,10 °C, 62,412 % y 0,0 mm respectivamente (Tabla 1); mientras que los muestreos nueve (May-15), diez (Jun-5) y once (Jun-12), sólo se capturó un individuo en cada uno de ellos, pertenecientes a H. februa, H. amphinome y T. laothoe respectivamente. Durante el primer muestreo (T = 29,17 °C, H.R = 84,38 % y Prec. = 2,0 mm) y los tres últimos muestreos (Jul-24, T = 31,05 °C, H.R = 86,693 % y 119,0 mm; Ago-08, T = 30,71 °C, H.R = 85,531 % y 88,0 mm; Ago-21, T = 29,60 °C, H.R = 82,443 % y 192,0 mm) no se registraron capturas de las especies de Biblidinae (Tabla 1).
Frecuencia de captura de mariposas Biblidinae por cebo
El cebo donde se capturó la mayor riqueza (6 especies) y abundancia (62 individuos) fue el de frutas fermentadas (Tabla 2), mientras que sólo dos especies se capturaron en calamar (H. amphinome y T. laothoe) y H. februa en el mixto. La especie H. februa se caracterizó por ser la más abundante en fruta fermentada, con 27 individuos (Tabla 2).
Análisis de agrupamiento y similitud (ANOSIM) y rutina SIMPER
Con respecto al agrupamiento ponderado, teniendo en cuenta la matriz de similaridad generada por el análisis de Bray-Curtis se formaron tres grupos, sin tener en cuenta las ramificaciones de los muestreos M1, M14, M15 y M16, donde no se registraron capturas (Tabla 1, Fig. 3). El primer grupo está formado por los muestreos M10 (Jun-05), M11 (Jun-12) y M13 (Jul-10) con una similaridad inferior al 30 %. El segundo grupo está formado por los muestreos M2 (Feb-5), M3 (Feb-19), M9 (May-15) y M12 (Jun-28), este último es junto con M3 los que presentaron la mayor similaridad con un valor de 73,44 % (Fig. 3). El tercer grupo lo conforman dos subgrupos, el primero (Similitud > 65 %) representado por M6 (Abr-02), M7 (Abr-16) y M8 (Abr-30); mientras que el segundo subgrupo está conformado por M5 (Mar-19) y M6 (Abr-02) con una similaridad del 79,43 % (Fig. 3).
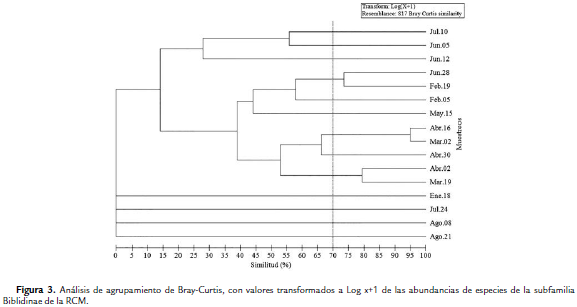
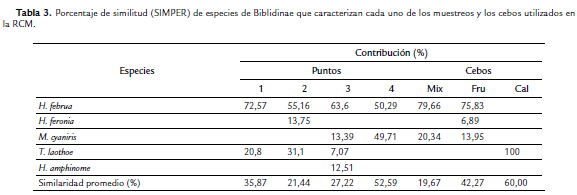
Por otro lado, con el análisis de similitud ANOSIM se determinó que existen diferencias significativas en la estructura de la comunidad entre los puntos estudiados (R = 0,118, p = 0,001): los puntos más disimiles fueron el 1 con el 3 (Estadístico R = 0,183, p = 0,001) y el 3 con el 4 (Estadístico R = 0,197, p = 0,001). En lo que respecta a la rutina SIMPER, H. februa contribuyó a la abundancia en todos los puntos con un mayor porcentaje en el punto 1 (72,57 %) (Tabla 3). En cuanto a los cebos, se presentaron diferencias significativas entre éstos (estadístico R = 0,372, p = 0,001) (Prueba ANOSIM). La mayor similaridad se observó entre mixto-fruta (estadístico R = 0,10, p = 0,2222), mientras que la menor se presentó entre calamar-fruta (estadístico R = 0,575, p = 0,001). El análisis SIMPER muestra que H. februa presentó la mayor contribución en el cebo mixto y el de fruta (79,66 % y 75,83 % respectivamente), mientras que T. laothoe fue la única especie que caracterizó el cebo de calamar en descomposición con una contribución del 100 % (Tabla 3), debido a que sólo se capturaron dos especies en este tipo de cebo.
Caracterización de la vegetación
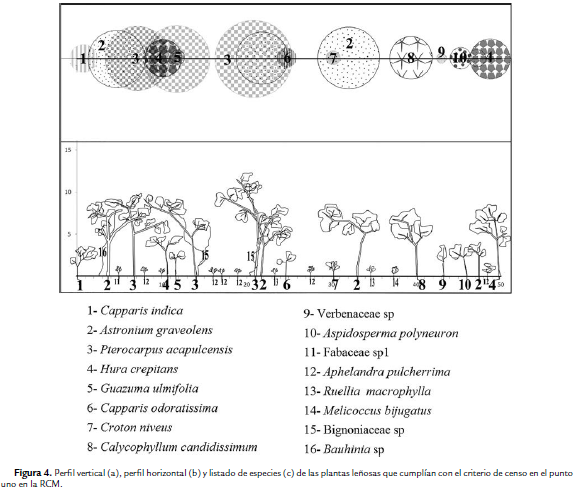
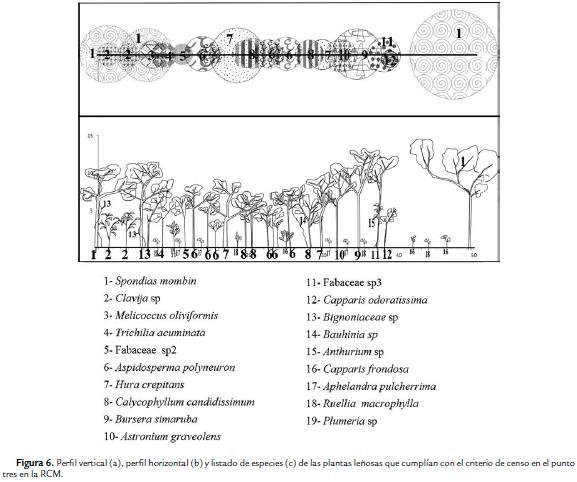
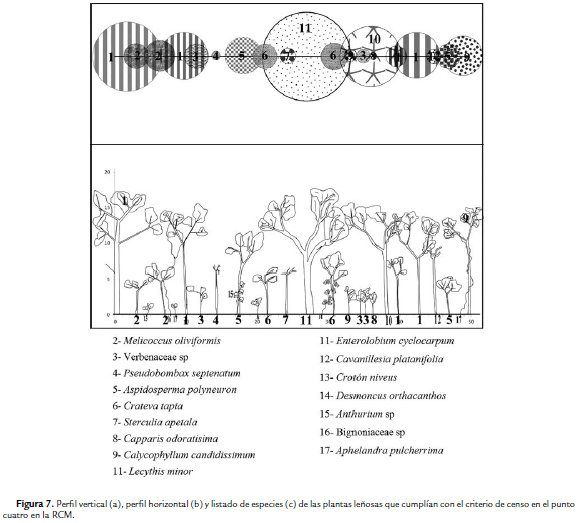
Para el área de estudio se registró un total de 19 familias y 34 especies de plantas vasculares con DAP superior a 2,5cm. La familia con la mayor riqueza fue Fabaceae con cinco especies. En el punto 1 se encontraron 16 especies y C. indica fue la más abundante (Fig. 4). De acuerdo a los parámetros de vegetación tomados en este punto, la densidad total (D) fue la más baja entre los cuatro puntos (D = 0,19 individuos/m2) y el área basal total (ABT) fue de 0,37 m2/ha. En el punto 2 se registró como especie más abundante Crateva tapia (L.), seguida de Calycophyllum candidissimum (Vahl) (Fig. 5). En este punto, el número de plantas leñosas fue de 24 individuos (D= 0,24 individuos/ m2) y un ABT de 1,11 m2/ha. En el Punto 3, S. mombin fue el árbol más abundante (Fig. 6) y la D fue de 0,28 individuos/ m2, con una ABT de 1,06 m2/ha; mientras que en el punto 4, H. crepitans fue la más abundante (Fig. 7); mientras que la D fue de 0,23 individuos/m2 y el ABT de 1,70 m2/ha.

Relación entre la abundancia de las mariposas Biblidinae con los parámetros de la vegetación (densidad total y área basal total) y las variables ambientales
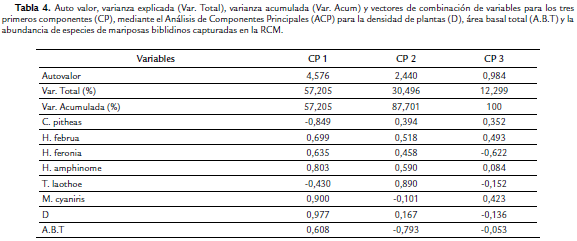
Teniendo en cuenta el análisis de componentes principales (ACP), las dos variables de la vegetación medidas en el área de estudio (D y ABT) están explicando el 87,7 % de la variación de los datos (componente 1 y 2). El componente 1 presenta la variacion más alta (57,2 %) y por lo tanto es donde están las variables con mayor efecto sobre el comportamiento de los datos (Tabla 4). Las variables que mas aportan al componente 1 de manera directa es D, seguido de la abundancia de M. cyaniris, H. amphinome, H. februa y H. feronia junto con ABT; mientras que la abundancia de T. laothoe fue una de las variables que más contribuyó al componente 2 (Tabla 4). Es importante resaltar que tanto C.pitheas como T. laothoe, presentó correlaciones negativas con D y ABT. De acuerdo a los valores de precipitación por muestreo, se observó que la mayor abundancia se registró durante los meses secos en la zona, con una baja captura de individuos cuando empiezan las lluvias fuertes. Este mismo comportamiento, se presentó para H. februa, que fue la especie más abundante en la RCM (Tabla 1).
DISCUSIÓN
El número de especies reportadas en este estudio corresponde al 3,33 % de las 180 especies de la subfamilia Biblidinae registradas para Colombia (Andrade-C, 2002) y al 46 % de las 13 registradas por Montero et al. (2009) para el departamento del Atlántico; encontrándose las mariposas más comunes de la subfamilia. Los valores de abundancia de H. februa muestran que esta especie es dominante en la reserva y que C. pitheas es una especie rara en el área.
Montero et al. (2009) reportan valores mayores de abundancia de C. pitheas en un bosque de galería en Guaibaná (Municipio de Repelón) y en un bosque ripario en San Juan de Tocagua (Municipio de Piojó) en el departamento del Atlántico; mientras que Prince et al. (2011) solamente reportan dos individuos en Corrales de San Luis (Tubará, Atlántico). Por lo tanto, estas diferencias en la abundancia de C. pitheas se debe a que esta especie es considerada rara y sus requerimientos de hábitat se restringen a áreas poco alteradas, disponibilidad de recursos alimenticios, presencia de plantas hospederas o factores microclimáticos que puedan afectar la presencia y abundancia de esta especie en un determinado lugar (Brown Jr y Freitas, 2002).También puede estar relacionado a que los biblidínos se encuentren en áreas de borde donde haya mayor disponibilidad de plantas hospederas y facilidad de acceder a la oferta de alimentos (Campos-Salazar et al., 2011), sin embargo en esta investigación no se muestreo en esta zonas; lo cual puede estar explicando la baja abundancia de C. pitheas. Además, la categoría de especie rara puede estar relacionada a los pocos números de individuos en las poblaciones de este grupo de mariposas en la RCM.
Con respecto a los muestreos, la mayor abundancia de biblidínos obtenida en el punto 3 puede deberse a la mayor densidad de plantas leñosas observadas en este lugar. Además el perfil vegetal mostró mayor riqueza de árboles con respecto a los demás puntos, constituyéndose en un espacio que puede aportar mayor cantidad de recursos como frutos y refugio e incluso área para perchar ya que este género de mariposas tiene preferencia por arboles grandes para la búsqueda de pareja. La abundancia de H. februa en este punto puede explicarse por la presencia y abundancia del árbol S. mombin, el cual presenta corteza grisácea y líquenes adheridos que pueden ser utilizados por esta especie para camuflarse debido al patrón de coloración de sus alas (Jenkins, 1983). Es importante resaltar que S. mombin produce muchos frutos que pueden exudar fermentos, los cuales constituyen un recurso abundante, favoreciendo la presencia y abundancia de H. februa y las otras especies de Hamadryas; ya que muchas especies de este género se benefician de este tipo de árboles, ya sea en bosques o en grandes potreros arbolados.
La densidad de árboles leñosos en el punto 3 también explica por qué la abundancia tiende a ser mayor durante casi todos los muestreos. Durante el quinto, se observaron en este punto frutos de S. mombin y otros árboles, lo que indica una sincronización entre los adultos de este grupo de mariposas y el aumento de exudados fermentados de frutos (Monge-Nájera et al., 1998), disparándose así la abundancia de biblidinos con respecto a los otros muestreos. Lo anterior concuerda con lo propuesto por Bandini et al. (2012), quienes sostienen que la distribución y abundancia de mariposas no es aleatoria, siguiendo el patrón de distribución de los recursos alimenticios. La persistencia de biblidinos en el punto 3 obedece a la preferencia que tienen las mariposas frugívoras por hábitats con mayor disponibilidad de recursos como los frutos, condición que se da en mayores proporciones en áreas con mayor cobertura vegetal. En estos hábitats es común encontrar gran variedad de frutos que sirven a las mariposas que se alimentan de jugos de frutas en descomposición. Además, en el bosque seco la sombra producida por el dosel de árboles (especies siempreverdes) que no pierden parte de su follaje en las épocas más secas, permite reducir el impacto de las altas radiaciones en la parte baja del bosque (Berdugo-Lattke y Rangel-Ch, 2014) donde se encuentran los frutos; la cual puede deshidratar este recurso rápidamente y de esta forma las sustancias acuosas y volátiles se evaporan más rápido, convirtiéndose en un alimento poco atractivo para este grupo de insectos.
De acuerdo a la variación temporal de la abundancia de Biblidinae en la RCM, los resultados demuestran que hay una tendencia a aumentar con las primeras lluvias en el área (finales de marzo). Estas observaciones pueden explicarse porque al caer las primeras lluvias aumenta el follaje, lo que genera mayor disponibilidad de nichos y refugios, afectando la dinámica poblacional de estas mariposas, especialmente en los muestreos posteriores (Vargas et al., 2011). La mayor proporción de Biblidinae en el quinto muestreo puede ser el resultado de la emergencia de imagos sincronizada con la tendencia en el cambio fenológico de la vegetación después de la precipitación registrada para el tercer muestreo. Sin embargo, la reducción de la abundancia después del octavo muestreo puede relacionarse con los valores más altos de precipitación, lo cual disminuye la actividad de los adultos de algunas especies de Biblidinae; coincidiendo con lo reportado por Boom et al. (2013) para la abundancia de mariposas diurnas en la Reserva Campesina de Luriza en el departamento del Atlántico. Sin embargo, estos resultados difieren a los reportados por Vargas et al. (2011) en ecosistemas de bosque seco en la Reserva Natural Las Delicias, quienes evaluaron la riqueza y abundancia de Hesperioidea y Papilionoidea. En dicha reserva, los autores encontraron que la abundancia de mariposas aumentó a medida que las precipitaciones fueron mayores. DeVries y Walla (2001) también reportan que la riqueza y abundancia de la comunidad de mariposas fruteras en un bosque húmedo en Sucumbio, Ecuador, es mayor durante los periodos de lluvias. Aunque en esta investigación no se realizó durante todo el año, se puede esperar que la abundancia de los biblidinae sea diferente durante los cambios de precipitación en el resto del año.
Barlow et al. (2007) sostienen que las diferencias en la variación temporal de mariposas no están claras y que la relación entre la estacionalidad y los tipos de hábitats podría depender de muchos factores incluyendo la subfamilia evaluada y las características del hábitat estudiado. Además, los análisis realizados por las investigaciones citadas son evaluaciones que tienen en cuenta la abundancia total de mariposas y no representan un seguimiento a las especies de Biblidinae. Por lo tanto, este estudio en la RCM constituye el primer trabajo sobre la variación temporal entre épocas de bajas y altas lluvias de las especies de la subfamilia Biblidinae en bosques secos del Caribe colombiano.
Los valores mayores de similaridad registrados entre los muestreos M4 (Mar-02) y el M7 (Abr-16), obedecen a que sólo H. februa y M. cyaniris aportaron a la abundancia, con valores similares. De igual manera, la similaridad entre el M5 (Mar-19) y M6 (Abr 02) se debe a los aportes a la abundancia de las dos especies mencionadas anteriormente más el aporte de H. feronia, la cual presentó su mayor incremento durante estos dos muestreos. El aumento en la abundancia de H. feronia en el quinto y sexto muestreo puede deberse también a la presencia de frutos de S. mombin (Anacardiaceae) en el punto 3.
La contribución a la abundancia aportada por H. februa (de acuerdo a la Rutina SIMPER) muestra la dominancia de esta especie de Biblidinae en la RCM. Se encontró que esta mariposa presenta una alta proporción de individuos con respecto a otras especies, debido a que sus plantas hospederas pueden mantener su follaje todo el año, incluso durante el transcurso de la época seca, proporcionando así un recurso constante para las larvas, lo que refleja un aumento en la abundancia de los adultos (Young, 1974); coincidiendo con lo reportado por este mismo autor para Guanacaste (Costa Rica).
El ACP muestra que H. februa, H. amphinome, H. feronia y M. cyaniris se congregan en puntos donde la densidad de árboles es mayor, por ser sitios que principalmente pueden ofrecer condiciones ideales para su establecimiento, por tener mayor cobertura vegetal y poca radiación solar, las cuales inciden en el mantenimiento de unas condiciones microclimáticas con una temperatura y humedad ambiental óptimas para su actividad, así como la oferta de recursos y sustratos para camuflarse. Por el contrario, C. pitheas y T. laothoe se distribuyen en las áreas más abiertas, donde la densidad y área basal de plantas leñosas es menor. Esto concuerda principalmente con lo reportado por Chacón y Montero (2007) en cuanto a T. laothoe y otras especies del género Callicore, ya que describen que estas especies habitan bordes de bosques, claros, márgenes de quebradas y caminos bordeados de bosques primarios y secundarios. Las diferencias mostradas por el ANOSIM entre el punto 3 y los restantes, demuestra que la estructura vegetal puede considerarse como un factor determinante en la distribución y abundancia de las especies de la subfamilia Biblidinae en la RCM.
Durante la época seca en la RCM, empieza la producción y maduración de frutos de muchos árboles del bosque seco (por ejemplo H. crepitans y S. mombin), los cuales se convierten en una oferta alimenticia para este grupo mariposas; razón por la cual es posible algunos ejemplares de Biblidinae. Además, en ausencia de lluvias se presentó un incremento de otros parámetros como la temperatura; lo que permite a las mariposas calentar sus alas para realizar sus actividades como la búsqueda de alimento, pareja o refugio. Prince et al. (2011) también reportaron que la temperatura puede explicar la variación de mariposas en dos fragmentos de Bs-T en Corrales de San Luis, Atlántico. Por otro lado, con la llegada de las lluvias fuertes en la zona, se presentó un aumento de la humedad relativa. En este caso, a medida que aumentan sus valores, la actividad de las Biblidinae disminuye; reduciéndose así la probabilidad de que sean capturadas. La temperatura y la humedad relativa son parámetros que dependen tanto de las precipitaciones como de la intensidad de la luz, por lo cual, es posible que estos parámetros sean un determinante en la actividad de estas mariposas. La reducción durante las fuertes lluvias se ve reflejada en la abundancia total de biblidínos. Este comportamiento puede obedecer a que durante los días con valores altos de precipitación se presentó una reducción de los valores de intensidad lumínica y temperatura debido a la nubosidad, por lo cual estas dos especies buscan refugios, disminuyen su actividad y no se logra su captura. Sin embargo, para realizar análisis más robustos y tener soportes estadísticos para ver cómo estas variables influyen en la variación de la abundancia de este grupo de mariposas, se hace necesario capturar un mayor número de individuos para próximos eventos de captura en un mayor lapso de tiempo.
La mayor frecuencia de captura de Biblidinae en el cebo de fruta fermentada muestra que las seis especies de este grupo mariposas en la RCM pueden capturarse utilizando solamente este cebo. Es importante anotar que aunque cinco especies se capturaron también en mixto, es posible que las mariposas llegaran a libar a éste por ser una mezcla de los dos cebos, buscando así azúcares, aminoácidos y sales o lo hacían por un cebo en particular. Por otro lado, la presencia de algunas especies de esta subfamilia en otros cebos como la carroña, se debe a la búsqueda de otros recursos (sales), los cuales no están disponibles en las frutas. La mayor contribución de H. februa en fruta fermentada y mixto muestran que utilizando estos cebos pueden capturarse un alto número de individuos de esta especie, según lo demuestra la rutina SIMPER. Esto indica que esta mariposa es atraída por los exudados y olores volátiles que se originan durante el proceso de fermentación de las frutas, los cuales aportan suficientes nutrientes para los altos requerimientos de energía de esta especie, especialmente para las actividades de vuelo con el fin de conseguir pareja.
CONCLUSIONES
La composición y abundancia de mariposas de la subfamilia Biblidinae varía espacialmente dentro del bosque. Los resultados sugieren que la estructura de la vegetación se considera un factor determinante en distribución y abundancia de biblidínos en la RCM. También se observó que las especies de Biblidinae capturadas presentan un patrón temporal entre los meses de muestreo que puede estar demarcado por los niveles de precipitación en la RCM (periodos de lluvia y sequía); los cuales inciden en la variación de factores abióticos (temperatura, humedad relativa) en este fragmento. La especie H. februa es la dominante en la RCM, presentando mayores valores de abundancia en los puntos con mayor densidad de plantas leñosas, mientras que C. pitheas es poco abundante en el bosque, congregándose donde hay menor densidad de plantas y más cercano al borde del fragmento. Por otro lado, el cebo de fruta fermentada es el más efectivo para capturar las mariposas Biblidinae, capturándose todas las especies registradas para la RCM y los valores más altos de abundancia.
AGRADECIMIENTOS
A la comunidad campesina de La Montaña, especialmente a Santiago Conrado "Chago" y Rafael Conrado "El Mono", por la atención en su vivienda y compañía durante los meses de muestreo. A IDEA WILD por el apoyo con la financiación de los equipos para tomar los datos de las variables ambientales Al Semillero de Investigación en Artrópodos e Insectos NEOPTERA (Grupo Biodiversidad del Caribe Colombiano), por su apoyo y colaboración en las jornadas de campo, en especial a Edgar Ortiz por su colaboración en el muestreo de la vegetación. A William Colina, por su colaboración en el transporte al área de estudio y a Yeison Gutiérrez Rojas, por la facilitación del mapa del área de estudio; así como los evaluadores anónimos que directa o indirectamente contribuyeron con sus comentarios al enriquecimiento de este documento. A la Facultad de Ciencias Básicas de la Universidad del Atlántico, por facilitar el Laboratorio de Zoología y el Museo UARC por facilitar el espacio para identificar y confirmar los especímenes capturados y poder culminar esta investigación.
REFERENCIAS
Álvarez M, Escobar F, Gast F, Mendoza H, Repinzzo A, Villarreal H. Bosque Seco Tropical. In: Chávez M, Arango N, editors. Informe Nacional sobre el Estado de la Biodiversidad, Colombia. Tomo I. Diversidad Biológica. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt; 1998. p. 56-71. [ Links ]
Andrade-C MG. Utilización de las mariposas como bioindicadoras del tipo de hábitat y su biodiversidad en Colombia. Rev Acad Colomb Cienc. 1998; 22(84):407-421. [ Links ]
Andrade-C GM. Biodiversidad de la mariposas (Lepidoptera: Rhopalocera) de Colombia. S.E.A. 2002;2:153-172. [ Links ]
Bandini D, Batista R, Prado Pi, Brown Jr Ks, Freitas Avl. The importance of small scales to the fruit-feeding butterfly assemblages in a fragmented landscape. Biodivers Conserv. 2012;21(3):811-827. Doi:10.1007/s10531-011-0222-x. [ Links ]
Barlow J, Araujo I, Overal Wl, Gardner Ta, Da Silva F, Lake Ir, et al. Diversity and composition of fruit-feeding butterflies in tropical Eucalyptus plantations. Biodivers Conserv. 2007;17(5):1089-1104. Doi: 10.1007/s10531-007-92240-0. [ Links ]
Berdugo-Lattke ML, Rangel-Ch JO. Composición florística del bosque tropical seco del santuario "Los Besotes" y fenología de especies arbóreas dominantes (Valledupar, Cesar, Colombia). Colombia Forestal, 2015;18(1):87-103. Doi: http://dx.doi.org/10.14483/udistrital.jour.colomb.for.2015.1.a05. [ Links ]
Boom C, Seña L, Vargas MA, Martínez N. Mariposas Hesperioidea y Papilionoidea (Insecta: Lepidoptera) en un fragmento de bosque seco tropical, Atlántico, Colombia. Bol Cient Mus Hist Nat. 2013;17(1):149-167. [ Links ]
Brown Jr KS, Freitas AVL. Butterfly communities of urban forest fragments in Campinas, São Paulo, Brazil: structure, instability, environmental correlates, and conservation. J Insect Conserv. 2002;6:217-231. [ Links ]
Butterflies of America. [Internet]. Illustrated Lists of American Butterflies. [18-Febrero- 2015;10- Enero 2015]. Disponible en http://www.butterfliesofamerica.com/. [ Links ]
Camero E, Calderón A. Comunidad de Mariposas (Lepidoptera: Rhopalocera) en un gradiente altitudinal del cañón del Río Combeima-Tolima, Colombia. Acta biol Colomb. 2007;12(2):95-110. [ Links ]
Campos-Salazar L, Gómez J, Andrade-c MG. Mariposas (Lepidoptera: Hesperioidea - Papilionoidea) de las áreas circundantes a las Ciénagas del Departamento de Córdoba, Colombia. Rev Acad Colomb Cienc.2011;35(134):45-60. [ Links ]
Chacón I, Montero J. Mariposas de Costa Rica. Costa Rica: Editorial INBio; 2007. p. 156-163. [ Links ]
Clarke KR, Gorley RN. PRIMER v6. User manual/ tutorial. Plymouth: PRIMER-E Ltd; 2006. p. 7-70. [ Links ]
Devries PJ, Walla TR. Species diversity and community structure in Neotropical fruit-feeding butterflies. Biol J Linn Soc Lond. 2001;74:1-15. [ Links ]
Erazo M, González L. Mariposas. En: Rodríguez J, Rueda J, Gutiérrez T, editores. Guía ilustrada de la Fauna del Santuario de Vida Silvestre Los Besotes, Valledupar, Cesar, Colombia. Serie de guías tropicales de campo N° 7 Conservación Internacional. Bogotá: Editorial Panamericana, Formas e Impresos; 2008. p. 28-116. [ Links ]
Gutiérrez L, Borjas R. Sistema de Areas Protegidas del Departamento del Atlántico. Colombia: Universidad del Atlántico, Corporación Autónoma Regional del Atlántico; 2006. p.16. [ Links ]
Holdridge L. Ecología Basada en Zonas de Vida. No. 34. San José: Instituto Interamericano de Ciencias Agrícolas; 1978. p. 13-26. [ Links ]
Jenkins DW. Neotropical Nymphalidae I. Revisión de Hamadryas. Bull Allyn Mus. 1983;81:1-146. [ Links ]
Kingsolver JG. Experimental analyses of wing size, flight, and survival in the western white butterfly. Evolution. 1999;53(5):1479-1490. [ Links ]
Martínez NJ, Cañas LM, Rangel JL, Barraza JM, Montes JM, Blanco OR. Coleópteros coprófagos (Scarabaeidae: Scarabaeinae) en un fragmento de bosque seco tropical en el departamento del Atlántico, Colombia. Boletín Museo de Entomología de la Universidad del Valle. 2010;11(1):21-30. [ Links ]
Libros de la Colección Ecológica del Banco de Occidente [Internet]. Colombia: I/M editores; [2006; Enero 2014]. Bosque Seco Tropical Colombia. Disponible en www.imeditores.com/banocc/seco/indice.htm. [ Links ]
Monge-Nájera J, Hernández F, González M, Soley J, Araya J, Zolla S. Spatial distribution, territoriality and sound production by tropical cryptic butterflies (Hamadryas, Lepidoptera: Nymphalidae): implications for the "industrial melanism" debate. Revista de Biología Tropical. 1998;46(2):297-330. [ Links ]
Montero F, Moreno M, Gutiérrez LC. Mariposas (Lepidoptera: Hesperioidea y Papilionoidea) asociadas a fragmentos de bosque seco tropical en el departamento del Atlántico, Colombia. Bol Cient Mus Hist Nat. 2009;13(2):157-173. [ Links ]
Mostacedo B, Fredericksen TS. Manual de métodos básicos de muestreo y análisis en ecología vegetal. Proyecto de Manejo Forestal Sostenible (BOLFOR). Santa Cruz de la Sierra, Bolivia: Editorial El País.; 2000. p. 31-32. [ Links ]
Newton AC. Forest Ecology and Conservation: A Handbook of Techniques. Oxford: Oxford University Press; 2007. p. 107-111. [ Links ]
Panza E, Torres P, De La Roda I, Berdugo A, Torres P, De La Rosa, et al. Esquema de Ordenamiento territorial (Ley 388 de 1997) del municipio de Juan de acosta, Atlántico. Juan de Acosta: Alcaldía municipal de Juan de Acosta 2001-2003.; 2001. 224 p. [ Links ]
Prieto C, Takegami C, Rivera JM. Estructura poblacional de Morpho sulkowskyi Kollar, 1850 (Lepidoptera: Nymphalidae) en un sector de la cordillera occidental, departamento del Cauca (Colombia). Entomotrópica. 2005;20(1):15-22. [ Links ]
PRIMER-E. [Internet]. Clarke KR, Warwick RM PRIMER-ELtd. Reino unido. [06- Junio-2015; Enero 2014]. Disponible en http://www.primer-e.com/. [ Links ]
Prince S, Vargas MA, Salazar JA, Martínez NJ. Mariposas Papilionoidea y Hesperioidea (Insecta: Lepidoptera) en dos fragmentos de bosque seco tropical en Corrales de San Luis, Atlántico, Colombia. Bol SEA. 2011;48:243-252. [ Links ]
Vargas MA, Martínez N, Gutiérrez L, Prince S. Riqueza y abundancia de Hesperioidea y Papilionoidea (Lepidóptera) en la Reserva Natural Las Delicias, Santa Marta, Magdalena, Colombia. Acta Biol Colomb. 2011;16(1):43-60. [ Links ]
Vázquez JA, Givnish JT. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. J Ecol. 1998;86(6):999-1020. [ Links ]
Villarreal H, Alvarez M, Córdoba S, Escobar F, Fagua G, Gast F. Manual de Métodos para el Desarrollo de Inventarios de Biodiversidad. Programa de Inventarios de Biodiversidad. Bogotá: Instituto de Investigaciones de Recursos Biológicos Alexander Von Humboldt; 2006. p. 81-4. [ Links ]
Young AM. On the biology of Hamadryas februa (Lepidoptera: Nymphalidae) in Guanacaste, Costa Rica. J Appl Entomol. 1974;76:380-393. [ Links ]
Zuur A, Ieno EN, Smith GM. Analysing Ecological Data. En: Gail M, Krickeberg K, Samet J, Tsiatis A, Wong W, editores. Statistics for Biology and Health. New York: Springer; 2007. p. 163-179. [ Links ]
