










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. vol.20 no.3 Bogotá set./dez. 2015
https://doi.org/10.15446/abc.v20n2.43776
Doi:http://dx.doi.org/10.15446/abc.v20n2.43776.
Artículo de investigación/Original research paper
SEED GERMINATION AND PLANT DEVELOPMENT IN Escobedia grandiflora (OROBANCHACEAE): EVIDENCE OF OBLIGATE HEMIPARASITISM?
Germinación de semillas y desarrollo de plantas en Escobedia grandiflora (Orobanchaceae): ¿Evidencia de hemiparasitismo obligado?
Edison CARDONA-MEDINA1,2, Sandra B. MURIEL RUIZ1.
1 Facultad de Ciencias Agrarias, Politécnico Colombiano Jaime Isaza Cadavid, Carrera 48 n.° 7- 151, Medellín, Colombia.
2 Mestrado en Recursos Genéticos Vegetais, Universidade Federal de Santa Catarina.
For correspondence: sbmuriel@elpoli.edu.co.
Received: 29th May 2014, Returned for revision: 26th March 2015, Accepted: 8th April 2015.
Associate Editor: Hernan Mauricio Romero.
Citation / Citar este artículo como: Cardona-Medina E, Muriel Ruiz SB. Seed germination and plant development in Escobedia grandiflora (Orobanchaceae): evidence of obligate hemiparasitism?. Acta biol. Colomb. 2015;20(3):133-140. doi: http://dx.doi.org/10.15446/abc.v20n2.43776.
ABSTRACT
Root parasitic plants can be facultative or obligate. Facultative parasites are able to complete their life cycle and their seeds can germinate without a host. Escobedia grandiflora is a poorly studied species in spite of its ancestral importance as dye of foods and medicinal use. The present study evaluates the states of seed, seedlings and mature plants, under presence and absence of possible hosts, for inferring the type of parasitism exhibited by E. grandiflora. Seeds were evaluated using two conditions each of light (12 and 0 hours) and temperature (20 °C and 25 °C); percentage germination, and germination speed were determined. The seeds did not require a host to germinate, as is typical of facultative parasitic plants. Percentage of germination varied between 66 % and 85.3 % and was not affected by light or temperature although germination speed was greater at 25 °C. Larger seeds had a higher percentage of germination and produced larger seedlings. The seedlings planted without a host did not survive, while those planted with Paspalum notatum had a 45 % survival rate, demonstrating that this is a critical stage of development, even with a host. Escobedia grandiflora plants sowed with grasses began the reproductive stage at the 28th week, and those planted with Pennisetum purpureum showed better performance, expressed in more haustoria, higher dry matter of total plant, rhizome and aerial stems. Plants sowed alone lived for more than six months, but they did not produce flowers or fruits. According to the behavior of seedlings and plants, E. grandiflora is an obligate parasite.
Keywords: Escobedia, obligate hemiparasite, parasitic plant, root parasitism, seed germination.
RESUMEN
Las plantas parásitas de raíces pueden ser facultativas u obligadas, las primeras pueden completar su ciclo de vida y sus semillas pueden germinar sin un hospedero. Escobedia grandiflora es una especie poco estudiada, a pesar de su importancia ancestral como colorante de alimentos y uso medicinal. Este estudio evaluó los estados de semilla, plántula y planta adulta, en presencia y ausencia de posibles hospederos para inferir sobre su tipo de parasitismo. En las semillas se evaluaron dos condiciones de luz (12 y 0 horas) y temperatura (20 °C y 25 °C), el porcentaje y velocidad de germinación. Las semillas no requirieron la presencia del hospedero para germinar. El porcentaje de germinación osciló entre 66 y 85,3 % y no fue afectado por la luz o la temperatura, aunque la velocidad de germinación fue mayor a 25 °C. Las semillas con mayor tamaño presentaron mayor porcentaje de germinación y produjeron plántulas más grandes. Las plántulas sembradas sin hospedero no sobrevivieron, mientras que las sembradas con Paspalum notatum, tuvieron una sobrevivencia del 45 %, evidenciando que este estado es crítico, aún con hospedero. Las plantas de Escobedia grandiflora sembradas con pastos, iniciaron la etapa reproductiva en la semana 28, y aquellas sembradas con Pennisetum purpureum presentaron más haustorios, y mayor materia seca en la planta total, rizoma y tallos aéreos. Las plantas sembradas solas vivieron más de seis meses, pero ellas no desarrollaron flores y ni frutos. Según el comportamiento de las plántulas y las plantas, E. grandiflora es parásita obligada.
Palabras clave: Escobedia, hemiparasíta obligada, planta parasítica, Parasitismo de raíz, germinación de semillas.
INTRODUCTION
Root parasitic plants are classified according to their degree of host dependence. They are considered either holoparasites or hemiparasites. Holoparasites are almost completely lacking in chlorophyll while hemiparasites have chlorophyll, are photosynthetically active, and derive some resources from their host through haustoria, the organ of parasitism (Yoder, 2001; Watling and Press, 2001; Nickrent, 2002; Bennett and Mathews, 2006). According to the degree of dependence upon host plant(s) for completing the life cycle, root hemiparasitic plants can be facultative or obligate. Facultative hemiparasites can survive in autotrophic form, independent of the host, for at least part or all of their life cycle. They can reproduce without a connection to a host, and when available, parasitize opportunistically on neighboring plants (Fay et al., 2010; Westwood et al., 2010). In contrast, obligate parasites must parasitize a host in order to complete their life cycle (Westwood et al., 2010).
One of the characteristics differentiating obligate and facultative plants has to do with seed germination behavior. In order to germinate, seeds of obligate parasitic plants require the presence of the host plant whose roots secrete substances that are perceived by the seed of the parasite and thus stimulate its germination (Bouwmeester et al., 2007; Westwood et al., 2010; Cardoso et al., 2011). For example, in species of Orobanche L. (Plakhine et al., 2012), Phelipanche Pomel (Matusova et al., 2004) and Striga Lour. (Matusova et al., 2004), a very low germination rate (<10 %) has been found in the absence of hosts in all cases. The system of chemical recognition assures that germination begins only when the root of the potential host is available in the immediate vicinity (Mohamed et al., 2001; Joel et al., 2011). The formation of haustoria also occurs in answer to chemical and tactile stimuli provided by the host roots (Westwood et al., 2010). The seeds of other species such as Rhamphicarpa fistulosa (Hochst.) Benth do not require the exudates of the host root to stimulate germination and are able to complete their life cycle without a host, albeit with reduced fitness evidenced by the production of only a few seeds relative to those grown with a host (Ouedraogo et al., 1999). Nevertheless, seeds of these species require light, and because they are very small and have little food reserve for the embryo, they must be very close to the surface of the ground in order to quickly begin photosynthesis (Ouedraogo et al., 1999).
Orobanchaceae is a cosmopolitan family consisting of approximately 90 genera and over 2060 species approximately (McNeal et al., 2013); 21 genera and around 300-330 species are native to the Neotropics, especially the Andean Region and higher altitudes in Brazil (Souza, 2012). This family is morphologically diverse and includes the greatest number of root parasite species in the Plant Kingdom, but only one non-parasitic genus Lindenbergia Lehm., which is phylogenetically supported as sister to the remainder of the family (Young et al., 1999; Phoenix and Press, 2005; Wolfe et al., 2005). Many species of this family are components of natural vegetation communities in different parts of the world, appreciated for their colorful flowers (Bouwmeester et al., 2007) and some, such as Euphrasia officinalis (L.) and Escobedia grandiflora (L.f) Kuntze, for their medicinal and food uses (Pennell, 1931; Trovato et al., 1999; Bussman et al., 2010). Nevertheless, many species are more well-known for their damaging effects when parasitizing economically important agricultural plants (Parker, 2009; Bouwmeester et al., 2007). In particular, species of the genus Alectra Thunb., Striga and Orobanche are considered noxious weeds of agriculture causing low productivity in a wide spectrum of economically important plants, especially in Africa and Asia (Wolfe et al., 2005; Van Ast and Bastiaans, 2006; Morawetz et al., 2010).
The degree of host specificity of species of Orobanchaceae is variable. There are records of simultaneous parasitism of various hosts, even of distant families (Yeo, 1964; Phoenix and Press, 2005; Parker, 2009; Qasem, 2009). For example, species of Euphrasia parasitize both monocot (Poaceae) and eudicot (Fabaceae) plants (Yeo, 1964). In contrast, some cases ofhigh host specificity have also been documented (Ren et al., 2010). Studies carried out to evaluate the possibility of establishing some parasitic species of Euphrasia (Yeo, 1961; 1964) under culture conditions found that the plants can grow when planted as seeds or seedlings in pots in which a host has previously been established. Yet, as a result of auto-parasitism, they were also able to achieve the flowering stage when planted very close together (Yeo, 1961; Yeo, 1964).
Escobedia Ruiz and Pav. is an American hemiparasitic genus (Bennett and Mathews, 2006; Carranza and Medina, 2008). Two phylogenetic analyses of the family placed this genus in the same clade of obligate parasitic species such as Striga, and as sister to the clade of Melasma P.J. Bergius and Alectra(Bennett and Mathews, 2006; Morawetz et al., 2010). In contrast to other genera of the Orobanchaceae family, there are no studies on the parasitic behavior of Escobedia species, nor records that relate it to the loss of economic crops.
Escobedia grandiflora is found from Central America (Mexico, Costa Rica, Panama) to South America (Colombia, Venezuela, Ecuador, Peru, Brazil, Paraguay and Argentina) and is widely distributed in Colombia, from 0 to 3500 m (Tropicos.org, 2013; review of JAUM, MEDEL, HUA and TOLI herbariums). This species has been important to ancestral communities, especially in the north of South America because of the use of its orange-colored roots as a natural food coloring (Pennell, 1931), and until a few decades ago, for medicinal purposes (Uribe, 2011). Even today, E. grandiflora is sold in local markets in Colombia, and it is considered a native species that can generate economic value, about which there is little information. It is thought that its use has been mainly extractive since no record of its cultivation has been found. This species, abundant in the past (Uribe, 2011), is now found sporadically in small patches especially along roads and in abandoned pastures.
Escobedia seeds are small and linear, similar to those of Melasma and Alectra (Falçao, 1980), and translucent in color except at the center (Souza and Giulietti, 2009). Over 300 seeds are formed in each capsule (Falçao, 1980). There have been no studies related to the germination capacity of E. grandiflora seeds, nor the effect of different hosts on the germination and development of the plants, except for a reference indicating a low percentage of viability of the seed (Castro et al., 2012).
The proposed objective of this study was to evaluate E. grandiflora seed germination, seedling, and plant development, with or without hosts of the Poaceae family, in order to make inferences about the degree of parasitism for this species.
MATERIALS AND METHODS
Study site
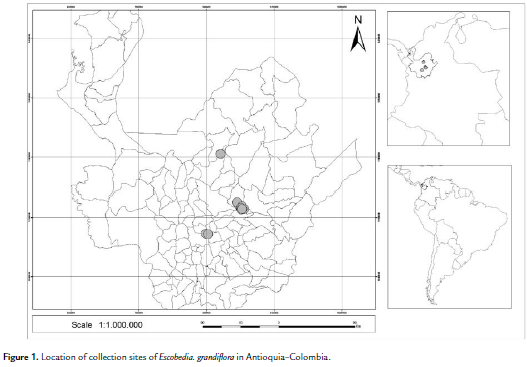
Escobedia grandiflora seeds were collected from capsules of individuals found in four municipalities in Antioquia (Colombia): Yarumal (07°09' N, 75°22' W), Gomez Plata (06°38' N, 75°11' W), Yolombo (06°34' N, 75°08' W) and Copacabana (06°18' N, 75°30' W) (Fig. 1).
The seeds were extracted manually and then selected with the aid of a stereoscope, in order to reject deformed ones.
The plants were collected from the same natural population located in the municipality of Copacabana. The monitoring of the plants was carried out at the Politecnico Colombiano Jaime Isaza Cadavid laboratories in the municipality of Bello, Antioquia.
Seed germination
All of the seeds were submerged in distilled water for 24 hours and were then divided into four germination treatments each with four repetitions. Variables in treatments were light and temperature. Two temperatures were evaluated: 20 °C and 25 °C, corresponding to values frequent on sites with wild populations of the species. The light treatments were 12 hours light/12 hours of darkness, and 0 hours light/24 hours of darkness.
Fifty seeds were set on moistened absorbent paper placed in plastic trays, and then covered with a thin, porous white veiling to allow for aeration and avoid a rapid loss of humidity. The seeds were watered periodically with distilled water and observed every 24 hours, for 31 days. They were considered to have germinated when the radicle became visible.
The variables evaluated were percent of germination and vigor, the latter measurement through speed of seed germination and seedling length (Gupta, 1993). Variables were determined in the following manner:
Percent of germination: %G=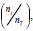 where n is the number of seeds germinated and nT is the total number of seeds. Germination speed was determined according to Raizada and Raghubanshi (2010):
where n is the number of seeds germinated and nT is the total number of seeds. Germination speed was determined according to Raizada and Raghubanshi (2010):
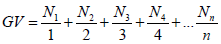
where Nn = number of germinated seeds on days 1, 2, 3, n etc.
Variance analysis was used to analyze the data after verifying fulfillment of the assumptions of homoscedasticity and distribution of residuals. The data on percentage of germination was transformed using the arcsine function. The Tukey Test was used as an a posteriori test.
One hundred and twenty seeds were used to evaluate the length of the E. grandiflora seedlings. Seeds had previously been hydrated in water for 24 hours at a temperature of 25 °C and with 12 hours of light. When the radicle emerged of the seed, it was then planted in a germinator containing a substrate taken from the four localities and mixed with soil rich in organic matter in a proportion of 2:1. During a 40-day period, five seedlings were measured every seven days. Different plants were chosen for each measurement because the stress produced by their extraction could also cause a delay in growth. The information was analyzed by means of multiple regression analysis using SAS System (SAS Institute Inc., Cary, North Carolina).
Seedling development
Seeds from two grasses previously planted in a germination tray were used. Eight days after their germination, a recently germinated E. grandiflora seedling was planted. The seedlings were divided into three treatments: E. grandiflora alone, E. grandiflora with Andropogon bicornis L., and E. grandiflora with Paspalum notatum Alain ex Flüggé. The plants were observed for 16 weeks, and every four weeks the variables of percentage, survival and length of the E. grandiflora seedlings were measured.
Plant development
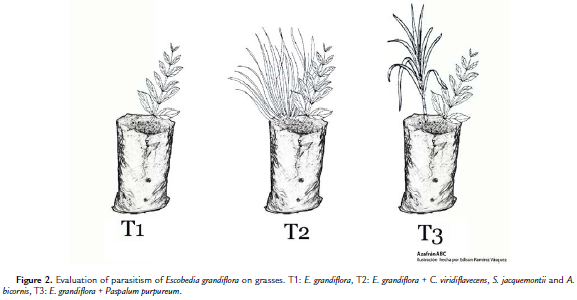
Sixty small plants were collected from a natural population of E. grandiflora, selected according to the following characteristics: height between 4.0 and 9.5 cm, a minimum of two leaves and root length ofbetween 3.0 and 9.0 cm. The plants were then taken to the laboratory, washed with water and planted in nursery bags (43 cm long and 18 cm wide) using a substrate consisting of sand, organic material and soil from areas where the plant was found. The plants were divided into three treatments of 20 plants each consisting of E. grandiflora alone (T1); E. grandiflora in association with the narrow-leafed grasses Calamagrostis viridiflavecens (Poir.) Steud., Sporobolus jacquemontii Kunth, and A. bicornis (T2); and E. grandiflora with Pennisetum purpureum (T3) (Fig. 2). T2 included three grasses because, although the initial collection was identified as a single species, three different species were later recognized. Thus, the three were included in all T2 repetitions. The species on the collection site were naturally associated with E. grandiflora. All bags received an application of diluted inorganic fertilizer (BP-150, Safer Agrobiológicos, Registro ICA 6870) every two weeks in order to guarantee the input of nutrients from the grasses.
The E. grandiflora plants were observed for 28 weeks, and measured every two weeks for variables of stem length, number of leaves and width-length of leaves in the middle third of the plant. On the last two dates, the distance between internodes, number of internodes, number of stems and thickness of stems were measured. At the end of this period, the plants were extracted from the bags in order to measure final root length and determine presence and number of haustoria. A repeated analysis measurement was used to analyze the data: the SAS System (SAS Institute Inc., Cary, North Carolina) and MIXED procedure. Comparison of measurements was carried out using the Tukey test. The Wilcoxon rank test was used to compare the variables measured on the last two dates.
RESULTS
Seed germination
Germination of the E. grandiflora seeds ranged between 66 % and 85.3 % (Table 1). The root emerged from the narrowest area of the seed and, later, the plumule with a pair of pubescent cotyledons although three cotyledons were observed in a few seeds. In cases where the plumule emerged first, the seedlings did not survive. The two conditions of temperature and luminosity had no effect on seed germination percentage .
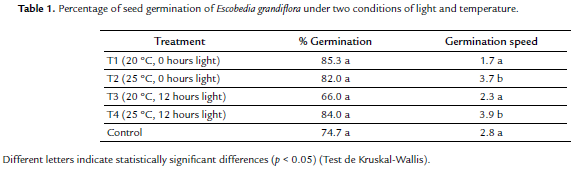
In contrast, germination speed was affected by temperature (F = 18.09; p = 0.003), being greater in seeds submitted to a temperature of 25 °C. The two conditions of light evaluated had no differential effect on rate of germination. The size of the seed determined the size of the seedling (R-adjusted = 63.42 %, p = 0.001). Thus, the longest and heaviest seeds showed the largest percentage of germination and produced larger seedlings.
During the germination tests, some seeds were affected by the Fusarium sp. fungus, which were discarded. The genus Fusarium have been reported on seeds of Striga and Alectra (Abbasher and Sauerborn, 1992; Marley et al., 2005).
Seedling development
During the first four weeks, the survival of E. grandiflora seedlings from sexual seeds was 100 %. At eight weeks, E. grandiflora without host had a survival percentage of 7.5 %, E. grandiflora with P. notatum of 95%, and E. grandiflora with A. bicornis of 60 %. At the end of the test, only E. grandiflora seedlings with P. notatum survived at 45 %. In general, the growth rate of the seedlings was slow: they grew an average of 1 mm and developed two leaves every four weeks. At 16 weeks, the roots began to exhibit the orange color typical of the species.
Plant development
In contrast to survival of seedlings, at four weeks the percentage of survival of small plants was 93.3 %. All of the plants, independent of the treatment, showed a slow growth rate and aerial stems formed from buds of the rhizome. The formation of branches of the aerial stem from axillary buds was an occasional event that could possibly be induced by the breaking of the main stem through wind action, or by damage or weakening of the main stem. In many apparently healthy plants, independent of treatment, the apical bud of the main stem would usually die and the aerial stem would then wither completely. Later, another stem would form from the new buds on the rhizome. This behavior led to a great variation in the data from all of the treatments as seen in the high values in standard deviation. Nevertheless, clear differences between treatments began to be noted by the 16th week. Between weeks 22 and 26, T3 (E. grandiflora+P. purpureum) presented a greater number of leaves, a greater length of aerial stems and larger leaves (Table 2). However in the 28th week, T2 (E. grandiflora+narrow-leafed grasses) caught up with T3 regarding number of leaves, length of aerial stem, size ofleaves and length of internodes, both being superior to T1 (E. grandiflora alone). The more significant event was that T2 and T3 initiated formation of reproductive structures, but T1 did not (Table 2). There were a greater number of internodes in T3 than in the other two treatments, but no differences among treatments as far as the variables of internode distance, number of aerial stems and stem thickness (Table 2).
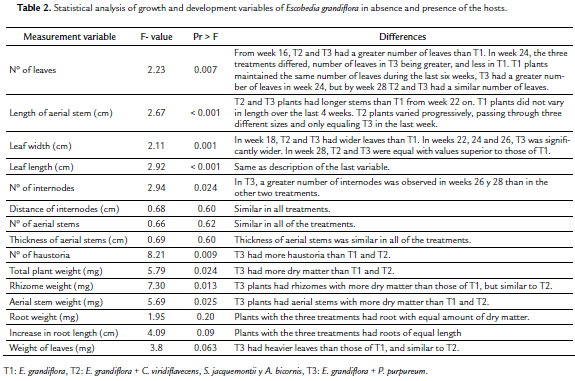
At the end of the experiment T3 E. grandiflora plants showed a greater number ofhaustoria, greater accumulation of total dry matter of the plant, rhizome dry matter, and of aerial stems than in T1 and T2 plants (Table 2). Root and leaf weight, as well as increased root length, was similar in the three treatments (Table 2). In T2 haustoria were observed in the three narrow-leafed grasses and, in some replicates simultaneous parasitism was also observed of two or three species. In general, the most parasitized hosts were S. jacquemontii and A. bicornis. In T1, haustoria sometimes formed without the presence of a host, and there were haustoria in T2 that were not parasitizing roots, but in any case auto-parasitism was not observed.
DISCUSSION
Escobedia grandiflora seeds do not require the presence of the host in order to germinate, which is typical of facultative parasitic plants (Těšitel et al., 2011). Although no studies on Escobedia seed germination are known, studies on seeds of related obligately parasitic genera, Alectra and Aeginetia L., recorded a lower percentage of germination than that of those found here (French and Sherman, 1976; Bennett and Mathews, 2006). So it can be argued that E. grandiflora seeds have a high percentage of germination. The 25 °C temperature at which the seeds had a greater germination speed is similar to the average temperature of locations where the plant is most abundant in Colombia. The two conditions of light evaluated were not decisive in germination.
Although the largest and heaviest seeds produced more vigorous seedlings, survival after four weeks was not successful without the host. Seedlings even with the host, were few in number and showed slow growth afterwards. It is possible that, as in other hemiparasitic species of the family, the state of the seedling is critical (Těšitel et al., 2011) although it initially seems to depend on the food reserve of the seeds, and on their own photosynthetic activity. Germinated seeds need to quickly parasitize a host in order to obtain nutrition. The food reserve within the seed is scarce given its morphology and small size. Although each capsule produces 300 seeds or more (Falçao, 1980) that are easily disbursed to new sites by wind, a low survival rate and a slow growth rate of the seedlings could imply serious consequences for the population and be a vulnerability factor for the species. This may explain the scarce disbursed natural populations that are presently observed in the Colombian Andean region, although each population is locally abundant.
Propagation of E. grandiflora from small plants between 4 and 9 cm in height produced an initially high rate of survival, 93 %, even without the presence of the host. This study found that the species was able to live without a host for over 28 weeks, and that only after week 18 is a clear disadvantage observed in plants alone with respect to plants with a host. The differences between treatments in the accumulation of dry matter from the rhizome, number of haustoria and dry matter from the aerial stems suggest that the plant initially accumulates an important food reserve in the rhizome. This permits it to survive for a time until parasitism is successfully established. Additionally, after the plant has been established it is able to form new functional roots even without a host. This is suggested by the similarity in root length and the accumulation of dry matter of this organ. The plants without host did not produce flowers or fruits/seeds, and, conversely the tendency in their development indicates that they will die after 28 weeks, while those formed with hosts will begin the reproductive phase.
Of the species considered in this study, the best hosts for E. grandiflora were P. purpureum, S. jacquemontii and A. bicornis. The first of these permitted rapid growth and there was a strong attachment of haustoria in all three. In order to evaluate the reproductive phase, the application of this study over longer periods of time is recommended. Hemiparasitic plants have an ambiguous relation with their host since they take nutrients from it but also compete with it for light thus affecting photosynthetic activity (Těšitel et al., 2011). In this experiment, the leaves of the host were pruned to favor the photosynthetic activity of the E. grandiflora. So it is possible that the results obtained will be modified if the host is allowed to develop completely.
The obligate hemiparasitic nature of E. grandiflora is confirmed, an aspect unknown until now in tropical American countries, where it is used for coloring foods (Pennell, 1931; Ulloa, 2006). Biological knowledge of the species is also increased in key aspects such as seed germination, seedling development and the recording of four hosts under natural conditions. Studies must be continued in order to evaluate the feasibility of cultivating this parasitic species of ethnobotanical importance in Central and South America.
CONCLUSION
Escobedia grandiflora demonstrates attributes of facultative hemiparasitic plants such as a high rate of seed germination in the absence of a host, and prolonged survival of plants with rhizomes in the absence of a host. Nevertheless, two important factors support its habit as an obligate hemiparasite. First, the seedling cannot survive alone and presents a low survival rate even with a host. Second, the reproductive stage only begins in plants with a host. This paper is the first revision of parasitism type presented by this species and by Genus Escobedia, which may be important to understand why E. grandiflora has been disappearing from the Colombian Andes.
ACKNOWLEDGMENTS
To the Direction of Research and the Faculty of Agricultural Sciences at the Politécnico Colombiano Jaime Isaza Cadavid for partially financing this study, to D Castañeda and JC Valdez who advised the geo-referencing of the sites, to JS Medina y C Reyes who helped locate the plants in the field, to EP González, V Alvarez, M Serna and LD Velez for their valuable suggestions regarding development and writing, to scientific illustrator E Ramírez, to the persons on site who received us and supplied information regarding the plants, to J Morawetz who resolved our initial concerns regarding the species and encouraged us to continue with the research, to D Giraldo who identified the grasses and to JM Velez of the Medel Herbarium.
REFERENCES
Abbasher AA, Sauerborn J. Fusarium nygamai, a potential bioherbicide for Striga hermonthica control in sorghum. Biol Control. 1992;2(4):291-296. Doi:10.1016/1049-9644(92)90021-5. [ Links ]
Bennett Jr, Mathews S. Phylogeny of the parasitic plant family Orobanchaceae inferred from phytochrome A. Am J Bot. 2006;93(7):1039-1051. Doi: 10.3732/AJB.93.7.1039. [ Links ]
Bouwmeester HJ, Roux C, Lopez-Raez JA, Becard G. Rhizosphere communication of plants, parasitic plants and AM fungi. Trends Plant Sci. 2007;12(5):224-230. Doi:10.1016/J.TPLANTS.2007.03.009. [ Links ]
Bussmann RW, Glenn A, Sharon D. Antibacterial activity of medicinal plants of Northern Peru-can traditional applications provide leads for modern science. Indian J Traditional Knowledge. 2010;9(4):742-753. [ Links ]
Cardoso C, Ruyter-SPIRA C, Bouwmeester HJ. Strigolactones and root infestation by plant-parasitic Striga, Orobanche and Phelipanche spp. Plant Sci. 2011;180(3):414-420. Doi:10.1016/J.PLANTSCI.2010.1 1.007. [ Links ]
Carranza E, Medina C. Una especie nueva de Escobedia (Orobanchaceae) del Estado de Michoacán, México. Acta Bot Mex. 2008;85:31-37. [ Links ]
Castro D, Arredondo J, Jaramillo D. Propagación in vitro del azafrán de raíz (Escobedia grandiflora L.f.) Kuntze. Revista Universidad Católica de Oriente. 2012;33:23-29. [ Links ]
Falçao C. Morfologia das sementes de 35 generos de Scrophulariaceae do Brasil - sua aplicação a sistemática desta família. Rodriguesia. 1980;53:33-107. [ Links ]
Fay MF, Bennett JR, Dixon KW, Christenhusz MJ. Parasites, their relationships and the disintegration of Scrophulariaceae sensu lato. Curtis's Bot Mag. 2010;26(4):286-313. Doi: 10.1111/j.1467-8748.2009.01668.x. [ Links ]
French RC, Sherman LJ. Factors affecting dormancy, germination, and seedling development of Aeginetia indica L.(Orobanchaceae). Am J Bot. 1976;63(5):558-570. [ Links ]
Gupta PC. Seed vigour testing. In: Agarwal PK, editors. Handbook of seed testing. New Delhi; 1993. p. 242-249. [ Links ]
Joel DM, Chaudhuri SK, Plakhine D, Ziadna H, Steffens JC. Dehydrocostus lactone is exuded from sunflower roots and stimulates germination of the root parasite Orobanche cumana. Phytochemistry. 2011;72(7):624-634. Doi:10.1016/j.phytochem.201 1.01.037. [ Links ]
Matusova R, Van Mourik T, Bouwmeester HJ. Changes in the sensitivity of parasitic weed seeds to germination stimulants. Seed Sci Res. 2004; 14(4): 335-344. Doi:http://dx.doi.org/10.1079/SSR2004187. [ Links ]
Marley PS, Kroschel J, Elzien A. Host specificity of Fusarium oxysporum Schlect (isolate PSM 197), a potential mycoherbicide for controlling Striga spp. in West Africa. Weed Res. 2005; 45(6):407-412. Doi: 10.1111/j.1365-3180.2005.00482.x. [ Links ]
Mcneal JR, Bennett JR, Wolfe AD, Mathews S. Phylogeny and origins of holoparasitism in Orobanchaceae. Am J Bot. 2013;100(5):971-983. Doi: 10.3732/ajb.1200448. [ Links ]
Mohamed KI, Musselman LJ, Riches CR. The genus Striga (Scrophulariaceae) in Africa. Ann Mo Bot Gard. 2001;88(1):60-103. Doi: 10.2307/2666132. [ Links ]
Morawetz JJ, Randle CP, Wolfe AD. Phylogenetic relationships within the tropical clade of Orobanchaceae. Taxon. 2010;100(2):416-426. [ Links ]
Nickrent DL. Parasitic plants of the world. In: J. A. López-Sáez, P. Catalán and L. Sáez, editors. Parasitic plants of the Iberian peninsula and Balearic Islands. Madrid: Mundi-Prensa; 2002. p. 7-27. [ Links ]
Parker C. Observations on the current status of Orobanche and Striga problems worldwide. Pest Manag Sci. 2009;65(5):453-459. Doi: 10.1002/ps.1713. [ Links ]
Pennell FW. Escobedia: A Neotropical Genus of the Scrophulariaceae. Proc Acad Nat Sci Philadelphia. 1931; 411-426. [ Links ]
Plakhine D, Tadmor Y, Ziadne H, Joel DM. Maternal tissue is involved in stimulant reception by seeds of the parasitic plant Orobanche. Ann Bot. 2012;109(5):979-986. Doi: 10.1093/aob/mcs016. [ Links ]
Phoenix GK, Press M. Linking physiological traits to impacts on community structure and function: the role of root hemiparasitic Orobanchaceae (ex-Scrophulariaceae). J Ecol. 2005;93(1):67-78. Doi: 10.1111/j.1365-2745.2004.00950.x. [ Links ]
Ouédraogo O, Neumann U, Raynal-Roques A, Sallé G, Tuquet C, Dembélé B. New insights concerning the ecology and the biology of Rhamphicarpa fistulosa (Scrophulariaceae). Weed Research. 1999;39(2)159-169. Doi: 10.1046/j.1365-3180.1999.00133.x. [ Links ]
Qasem J. Parasitic weeds of the Orobanchaceae family and their natural hosts in Jordan. Weed Biol Manag. 2009;9(2):112-122. Doi: 10.1111/j.1445-6664.2009.00328.x. [ Links ]
Ren YQ, Guan KY, Li AR, Hu XJ, Zhang L. Host dependence and preference of the root hemiparasite, Pedicularis cephalantha Franch.(Orobanchaceae). Folia Geobot. 2010;45(4):443-455. Doi 10.1007/s12224-010-9081-6. [ Links ]
Souza VC, Giulietti AM. Levantamento das espécies de Scrophulariaceae sensu lato nativas do Brasil. Pesquisa, Botânica. 2009;60:7-288. [ Links ]
Souza VC. Neotropical Orobanchaceae. In: Milliken, W., Klitgård, B. & Baracat, A. (2009 onwards), Neotropikey-Interactive key and information resources for flowering plants ofthe Neotropics. [Cited: 1 feb 2014]. Available at: http://www.kew.org/science/tropamerica/neotropikey/families/Orobanchaceae.htm. [ Links ]
Těšitel J, Lepš J., Vráblová M, Cameron DD. The role of heterotrophic carbon acquisition by the hemiparasitic plant Rhinanthus alectorolophus in seedling establishment in natural communities: A physiological perspective. New Phytol. 2011;192(1): 188-199. Doi: 10.1111/j.1469-8137.2011.03777.x. [ Links ]
Tropicos.org. [Internet]. Missouri: Missouri Botanical Garden; [Cited 1 feb 2014]. Available at: http://.www.tropicos.org/. [ Links ]
Trovato A, Monforte MT, Forestieri AM, Pizzimenti F. In vitro anti-mycotic activity of some medicinal plants containing flavonoids. Boll Chim Farm. 1999;139(5): 225-227. [ Links ]
Ulloa C. Aromas y sabores andinos. In: Moraes, M., B. Ollgard, LP Kvist, F. Borchsenius and H. Balslev, editores. Botánica económica de los Andes Centrales. La Paz: Universidad Mayor de San Andrés.; 2006. 319 p. [ Links ]
Uribe JA. Flora Sonsonesa. Colección Bicentenario de Antioquia, memorias y horizontes. Imprenta departamental de Antioquia: 2011. p. 194. [ Links ]
Van Ast A, Bastiaans L. The role of infection time in the differential response of sorghum cultivars to Striga hermonthica infection. Weed Res. 2006;46(3): 264-274. Doi: 10.1111/j.1365-3180.2006.00507.x. [ Links ]
Watling JR, Press MC. Impacts of infection by parasitic angiosperms on host photosynthesis. Plant Biol. 2001; 3(3): 244-250. Doi: 10.1055/s-2001-15195. [ Links ]
Westwood JH, Yoder JI, Timko MP, Depamphilis CW. The evolution of parasitism in plants. Trends Plant Sci. 2010; 15(4): 227-235. Doi:10.1016/J.TPLANTS.2010.01.004. [ Links ]
Wolfe AD, Randle CP, Liu L, Steiner KE. Phylogeny and biogeography of Orobanchaceae. Folia Geobot. 2005;40(2-3):115-134. [ Links ]
Yeo PF. Germination, seedlings, and the formation of haustoria in Euphrasia. Watsonia. 1961; 5(1):1-22. [ Links ]
Yeo PF. The growth of Euphrasia in cultivation. Watsonia 6(1): 1-24. [ Links ]
Yoder JI. 2001 Host-plant recognition by parasitic Scrophulariaceae. Plant Biol. 2001;4(4):359-365 Doi:10.1016/S1369-5266(00)00185-0. [ Links ]
Young ND, Steiner KE, Depamphilis CW. The evolution of parasitism in Scrophulariaceae/Orobanchaceae: plastid gene sequences refute an evolutionary transition series. Ann Mo Bot Gard. 1999; 876-893. [ Links ]
