










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkActa Biológica Colombiana
versão impressa ISSN 0120-548X
Acta biol.Colomb. vol.21 no.1 Bogotá jan./avr. 2016
https://doi.org/10.15446/abc.v21n1.49251
Doi: http://dx.doi.org/10.15446/abc.v21n1.49251.
IDENTIFICACIÓN DE QTLs ASOCIADOS A CARACTERES DE ARQUITECTURA VEGETAL EN YUCA (Manihot esculenta)
Identification of QTLs Associated to Plant Architecture in Cassava (Manihot esculenta)
Ruben Eduardo MORA MORENO1, Johana Carolina SOTO1, Camilo LÓPEZ1.
1 Departamento de Biologia. Universidad Nacional de Colombia, Bogotá, Colombia.
For correspondence. celopezc@unal.edu.co
Received: 18th February 2015, Returned for revision: 19th July 2015, Accepted: 8th September 2015.
Associate Editor: Leonardo Galindo.
Citation / Citar este artículo como: Mora Moreno RE, Soto JC, López C. Identificación de QTLs asociados a caracteres de arquitectura vegetal en yuca (Manihotesculenta). Acta biol. Colomb. 2016;21(1):99-109. doi: http://dx.doi.org/10.15446/abc.v21n1.49251.
RESUMEN
La yuca (Manihot esculenta) es el cuarto cultivo en importancia a nivel mundial como fuente de calorías para la población humana después del arroz, el azúcar y el maíz, posicionándose por esta razón como un cultivo primordial para la seguridad alimentaria. Su arquitectura ha sido considerada como un factor clave que subyace a la fisiología del rendimiento, relacionando características morfológicas con productividad. En este trabajo se evaluaron diferentes características de arquitectura vegetal en yuca. Los caracteres fueron evaluados en una población F1 compuesta por 133 hermanos completos (familia K) sembrados en dos lugares biogeográficamente diferentes: La Vega (Cundinamarca) y Arauca (Arauca) en Colombia. Las características evaluadas relacionadas con la arquitectura vegetal fueron altura de la planta (AT), número de brotes (NB), longitud entrenudos (LE), número de raíces (NR), peso de raíces (PR), pigmentación del peciolo (PP), área de la hoja (AH) y tipo de hoja (TH). A partir de los datos obtenidos y empleando un mapa genético de alta densidad basado en SNPs (Single Nucleotide Polymorphisms) se llevó a cabo un análisis de QTLs (Quantitative Trait Loci). Se lograron identificar tres QTLs para La Vega asociados con los caracteres altura total, número de brotes y área de la hoja. Para Arauca se detectaron tres QTLs asociados con altura total, longitud de entrenudos y número de brotes. Los QTLs se distribuyeron en cuatro grupos de ligamiento y explicaron entre 18,93 y 41,92 % de la variación genética.
Palabras clave: Arquitectura vegetal, grupo de ligamiento, mapeo genético, marcador molecular, QTLs, yuca.
ABSTRACT
Cassava (Manihot esculenta) is the fourth most important crop worldwide as a source of calories for the human population after rice, sugar and corn and therefore it is considered as a staple crop. Cassava's architecture has been considered as a key factor underlying the physiology of yield, relating morphological traits with productivity. In this work different characteristics of plant architecture were evaluated in a cassava F1 population composed by 133 complete siblings (family K) planted in two biogeographically different zones: La Vega (Cundinamarca) and Arauca (Arauca) in Colombia. The characteristics evaluated related to the vegetal architecture were plant height (AT), number of shoots (NB), internodes length (LE), number of roots (NR), root weight (PR), petiole pigmentation (PP), leaf area (AH) and leaf type (TH). From the data obtained and using a SNP- (Single Nucleotide Polymorphism) high-density genetic map a QTLs analysis (Quantitative Trait Loci) was carried out. It was possible to identify three QTLs for La Vega associated with characters plant height, internodes length and leaf area. From the Arauca's dataset, three QTLs were detected associated with plant height, number of shoots and internodes length. The QTLs were distributed into four linkage groups and explained between 18.93 and 41.92 % of genetic variation.
Keywords: Cassava, genetic mapping, linkage group, molecular marker, vegetal architecture, QTL.
INTRODUCCIÓN
La yuca (Manihot esculenta, Crantz), representa la cuarta fuente de calorías después del arroz, el azúcar y el maíz para cerca de un billón de personas en el mundo. La yuca es principalmente un cultivo de los trópicos, el cual se cultiva en terrenos marginales, con largos períodos de sequía y en suelos que pueden ser infértiles y ácidos (Mederos, 2006). El almidón producido por las raíces almacenadores se destina principalmente a cuatro mercados según sus usos: i) como raíz fresca y procesada para alimentación humana, ii) como materia prima en la industria de alimentos con el fin de producir harina, iii) en la producción de alimentos balanceados para animales y iv) como un producto intermediario en otro tipo de industrias no relacionadas con la alimentación (Suárez y Mederos, 2011). La yuca es principalmente cultivada por pequeños agricultores, los cuales son en la mayoría de los casos de bajos ingresos pero que a pesar de su producción a pequeña escala se produce de manera relativamente eficiente como para suplir las demandas de consumo de los productores. Según la FAO, si la yuca logrará desarrollar su potencial para entrar como materia prima en una gran variedad de productos procesados existiría una mayor demanda del producto y esto traería como consecuencia no solo una transformación agrícola sino que apoyaría al crecimiento económico de las áreas marginales donde la yuca es cultivada (FAO, 2013).
La yuca se caracteriza por ser un arbusto perenne, que puede alcanzar tamaños muy variables, entre uno y cinco metros, y que idealmente precisa de un fotoperiodo corto. La yuca presenta flores masculinas separadas de las femeninas (planta monoica), de ramificación simpoidal (Cock, 1989). Los nudos y entrenudos alternados se pueden observar relativamente bien a lo largo de todo el tallo, en donde sobresalen un tipo de protuberancias que marcan la presencia anterior de las hojas (Ospina y Ceballos, 2012). La longitud del entrenudo es otra característica del tallo y este carácter está condicionado en gran medida por el medio en el que se desarrolla el cultivo (Suarez y Mederos, 2011). El grosor del tallo es importante y se ha asociado directamente con alto rendimiento (Suarez y Mederos, 2011). Las hojas son simples con una lámina foliar y un peciolo. La lámina foliar se caracteriza por ser palmeada y puede presentar un numero variable de lobulos cuyo numero cambia cuando se comparan diferentes variedades y por lo general es impar, oscilando entre tres y nueve (Ospina y Ceballos, 2012). Los pecíolos son largos y delgados, cuyo color es variable y puede ser rojo o rojo verdoso en unas variedades mientras que en otras puede variar entre el verde rojizo y el verde (Ospina y Ceballos, 2012). Como el número de lóbulos y el color del peciolo, también hay una amplia variación en el tamaño de la hoja, la cual en algunos casos es una característica propia de cada variedad aunque se ve afectada en gran medida por factores ambientales. Las primeras hojas, aquellas que aparecen en los primeros cuatro meses, son más grandes que las se producen después del cuarto mes (Suarez y Mederos, 2011). Mientras que las raíces de yuca tienen una amplia gama de usos, las hojas no tienen la misma importancia, aunque en algunos países de África las hojas son utilizadas para el consumo humano aprovechando el gran valor nutricional que poseen, en donde sobresale el alto nivel de proteína (entre el 18 al 22 %) (Gil y Buitrago, 1990).
La principal característica de la yuca son sus raíces gracias a su alto contenido de almidón, lo que le da su gran valor económico (Ospina y Ceballos, 2012). Las características morfológicas, como las mencionadas anteriormente, permiten tipificar las especies dentro del género Manihot, pero al ser afectadas por el ambiente puede llevar a una sobre estimación en el número de especies (Suarez y Mederos, 2011). El efecto que ejerce el ambiente sobre la morfología y arquitectura vegetal en cada variedad de yuca puede ser bastante drástico. Puede suceder por ejemplo que la arquitectura típica que presente una variedad en un ambiente determinado sea bastante diferente cuando es plantada en otras condiciones edafoclimaticas (Suarez y Mederos, 2011).
En plantas, un gran porcentaje de las características de importancia económica son controladas por la acción de varios genes, por lo cual son denominadas poligénicas, cuantitativas o de herencia compleja (Ferreira y Grattapaglia, 1998). Las diferentes características morfológicas relacionadas con la arquitectura vegetal son consideradas de tipo cuantitativo. En este tipo de rasgos, si bien el ambiente juega un papel muy importante en la expresión del carácter, éste está determinado también por la acción de varios genes (Ferreira y Grattapaglia, 1998).
Los loci responsables de la expresión de las características cuantitativas son definidos como QTLs (del inglés Quantitative Trait Loci) (Ferreira y Grattapaglia, 1998) y su identificación a través del uso de mapas genéticos se constituye en una herramienta importante dentro de los programas de mejoramiento de plantas (Cerón y Sahagún, 2007; López, 2011).
El primer mapa genético de yuca fue obtenido a finales de los 90s el cual fue construido principalmente empleando marcadores de tipo RFLPs (Fregene et al., 1997). Actualmente existen varios mapas genéticos de yuca basados en una amplia gama de marcadores moleculares como microsatélites (Mba et al., 2001), ESTs (Expressed Sequences Tags) (Lopez et al., 2007) y SNPs (Single Nucleotide Polymorphisms) (Rabbi et al., 2014; Soto et al., 2015).
Los diferentes mapas genéticos se han empleado en la identificación de QTLs asociados con resistencia a enfermedades (Jorge et al., 2001; Akano et al., 2002) con caracteres agronómicos y morfológicos importantes así como caracteres relacionados con rendimiento (Okogbenin y Fregene, 2003; Okogbenin y Fregene, 2006; Okogbenin et al., 2007).
Recientemente se ha desarrollado un mapa genético basado en 2141 marcadores de tipo SNPs empleando la estrategia de GBS (Genotyping By Seqencing) en la misma población utilizada para la construcción del primer mapa genético de yuca (Soto et al., 2015).
Con el objetivo de realizar un análisis de caracteres morfológicos relacionados con la arquitectura vegetal, no evaluados previamente y que puedan tener algún tipo de relación con el rendimiento de la planta, en este trabajo se evaluaron ocho caracteres de arquitectura vegetal en la población biparental empleada en la construcción del mapa de yuca. A partir de esta información y del mapa genético disponible se pudieron identificar varios QTLs asociados a diferentes características de arquitectura vegetal.
MATERIALES Y MÉTODOS
Mapa genético y condiciones geográficas
El mapa genético empleado en este estudio comprende 2141 marcadores moleculares tipo SNP provenientes de la progenie de 133 individuos de una familia de hermanos completos (Soto et al., 2015). Cada uno de estos individuos así como los parentales fueron propagados en campo en Arauca (Universidad Nacional de Colombia, sede Orinoquía) (Latitud 7°' 22.32'' N, Longitud 70°44'42.50'' W) y en La Vega (Cundinamarca) (Latitud 0,5°,00'44.188'' N, Longitud 74°21'31.005'' N). La Vega presenta una temperatura media de 22 °C, humedad relativa media de 83 % y precipitación anual de 1900 mm (IDEAM, 2002). La temperatura media anual de Arauca es de 28 °C, presenta una humedad relativa media de 79 % y precipitación anual de 1763 mm (PBOT Municipio de Arauca, 2000).
Población de mapeo
La población estuvo formada por 133 plantas F1, hermanos completos, provenientes de un cruce intraespecífico entre la variedad TMS 30572 (parental femenino), un cultivar elite desarrollado en el IITA (International Institute of Tropical Agriculture) en Nigeria, y la variedad CM 2177-2 (parental masculino), un exitoso cultivar desarrollado por el CIAT (Centro Internacional de Agricultura Tropical) en Colombia. Una de las características principales de este cruce es que presenta una segregación para varios rasgos importantes (Fregene et al., 1997).
Fenotipificación
Tres estacas de cada individuo de la población de mapeo se sembraron en los dos ambientes: Arauca (Arauca) y La Vega (Cundinamarca). Las estacas de cada individuo fueron suministradas por la Unidad de Mejoramiento Genético de Yuca del CIAT y fueron sembradas a una distancia aproximada de 1 m de distancia. Las plantas estuvieron sometidas a las condiciones ambientales naturales. Los siguientes carácteres fueron evaluados a cada planta de cada uno de los individuos: Altura de la planta (AT), definida como la longitud total del tallo o la parte aérea de la planta. Longitud de entrenudos (LE): distancia entre cada uno de los brotes, se toma la medida en el segundo tercio del tallo central. Número de brotes (NB), es el número total de brotes que se encuentran sobre el tallo principal. Peso de raíces (PR), es el peso de todas las raíces almacenadoras en la planta. (NR), es el número de raíces presentes en la planta. Área de la hoja (AH), medido como el promedio del área de las hojas 5, 6 y 7. Pigmentación del peciolo (PP) medido en una escala de 0 a 3 según la coloración. Tipo de hoja (TH) entre oblonga o lanceolada. Los datos para AT, LE, NB se tomaron a lo largo del tiempo, en las semanas 4, 12 y 14 post- siembra (sps). Los datos para PR, NR fueron tomados a la semana 14 post-siembra y los datos para PP, AH y TH fueron obtenidos en la semana 19 post-siembra.
Mapeo y análisis de QTLs
Se utilizó la estadística descriptiva para los datos cuantitativos (medidas de tendencia central, de variación y de desviación) y se construyeron histogramas de frecuencia empleando el software Microsoft Excel.
Se realizó análisis de QTL empleando el mapeo por intervalo compuesto (MIC), el cual requiere un mapa genético y la determinación de un LOD (del inglés logarithm base 10 of odds). El método MIC detecta un intervalo flanqueado por dos marcadores dentro del cual se halla un QTL. En este análisis los genotipos de los marcadores son permutados 1000 veces para establecer el umbral o LOD a partir del cual un marcador sería responsable de la variabilidad del rasgo. Posteriormente se adicionan loci cercanos, generalmente de 20 a 40 cM que generan efectos pequeños sobre el rasgo.
Se empleó el software R versión 3.1.1 y la librería "qtl", para el análisis de cada uno de estos rasgos fenotípicos y su posible asociación con marcadores moleculares presentes en el mapa genético.
RESULTADOS
Evaluación fenológica y análisis descriptivo
A partir de 133 individuos (códigos) de la familia K, se logró obtener semilla y germinación exitosa para 100 y 105 códigos en La Vega y Arauca, respectivamente. Los individuos fueron fenotipificados para los caracteres morfológicos altura de la planta (AT), longitud entrenudos (LE), número de brotes (NB), número de raíces (NR), peso de raíces (PR), área de la hoja (AH), pigmentación del peciolo (PP) y tipo de hoja (TH). Los caracteres fueron evaluados en diferentes tiempos según características propias de cada rasgo. Así, para AT, LE, y NB se evaluaron a las 4, 12 y 14 semanas post siembra (sps), los valores para los caracteres NR y PR se tomaron a la 14 sps y para AH, PP y TH los datos fueron obtenidos en la 19 sps. No en todos los casos se evaluó el mismo número de individuos en cada semana debido a que se presentó mortalidad en algunas plantas a lo largo del experimento.
En la mayoría de los casos se obtuvieron valores para tres plantas por cada código. A partir de los datos obtenidos (aproximadamente 860 en La Vega y 760 en Arauca) se realizó un análisis descriptivo. Para cada carácter evaluado se observa que existen rangos de variación muy amplios, seguramente debido a la alta segregación que presenta esta población, al igual que desviaciones estándar elevadas (Tabla 1). Así por ejemplo para PR se obtuvieron valores que variaron entre 10 y 800, mientras que para NB los rangos estuvieron entre 10 y 83 en La Vega y 18 a 275 en Arauca (Tabla 1). El rasgo morfológico con menor variabilidad fue NR mostrando que el ambiente puede tener bajo efecto sobre este rasgo (Tabla 1).
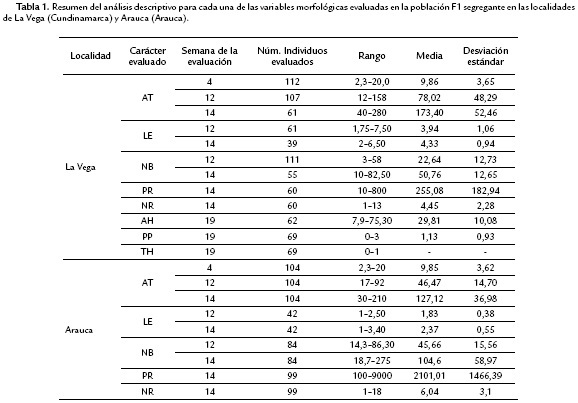
Cuando se compara el comportamiento de los rasgos morfológicos en los dos ambientes (La Vega y Arauca) se observa que es bastante diferente, lo cual es previsible considerando que el componente ambiental juega un papel muy importante en este tipo de rasgos cuantitativos. Por ejemplo, en La Vega las plantas tienden a tener una longitud de entrenudo mayor con un menor número de brotes por planta, mientras que en Arauca las plantas tienen un mayor número de brotes con una longitud de entrenudo menor.
Se observa también una alta diferencia en los valores obtenidos para rasgos que tienen que ver con el rendimiento (NR y PR) siendo en Arauca donde se obtuvieron los mayores valores (Tabla 1). Cabe destacar que si bien La Vega y Arauca son dos regiones donde el cultivo de yuca se adapta satisfactoriamente, las condiciones ambientales son bastante diferentes (ver materiales y métodos).
Los valores de distribución de frecuencia de características como AT, LE, NB, NR, PP, AH y TH muestran una tendencia central aunque no necesariamente sean normales, mientras que PR no presenta una tendencia definida. Se pudo determinar que para la mayoría de los caracteres evaluados, los dos parentales en Arauca muestran fenotipos claramente diferenciables. Sin embargo, en La Vega desafortunadamente no se pudo contar con semilla para el parental materno, por lo cual no se pudo establecer esta diferenciación (Fig. 1). Sin embargo, la ausencia de información para este parental no representa una dificultad en la posterior identificación de QTLs ya que el software empleado (R) hace una estimación sobre los individuos de la familia segregante a partir de los marcadores identificados en el ensamblaje del mapa de ligamiento.
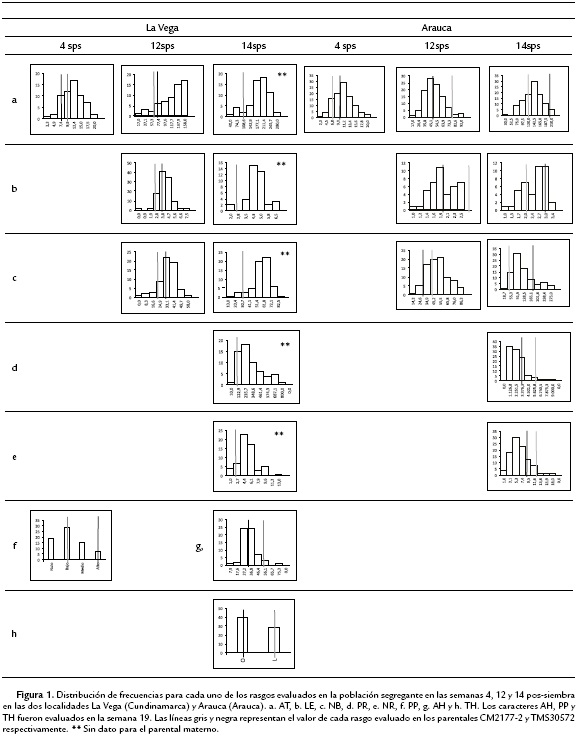
Identificación de QTLs
Con el objetivo de determinar qué regiones genómicas pueden estar regulando de manera parcial los diferentes rasgos morfológicos evaluados, se procedió a realizar un análisis de QTLs. Para ello se aprovechó la disponibilidad de un mapa genético que cuenta con 2141 marcadores de tipo SNPs obtenidos mediante la aproximación GBS (Soto et al., 2015). Para determinar una asociación entre los marcadores y los rasgos fenotípicos evaluados se empleó como umbral un valor de probabilidad mínimo de 0,05. Se consideraron QTLs significativos aquellos que tuvieran valores de LOD por encima del establecido por las permutaciones.
Un total de seis QTLs fueron identificados (qLV_1, qLV_2, qLV_3, qAr_1, qAr_2 y qAr_3) los cuales se encuentran distribuidos en cuatro grupos de ligamiento (LG3, LG5, LG13 y LG15). El porcentaje de la variación explicada por el factor genético (R2) para los diferentes rasgos varió entre 18,93 % y 41,92 % donde el mayor y menor valor de R2 corresponde a los rasgos NB en la segunda evaluación en La Vega y a AT en la segunda evaluación en Arauca, respectivamente. En general los intervalos en que se encuentran ubicados los QTLs varían entre 0,48 cM y 7,91 cM. Para el caso de los datos obtenidos de individuos crecidos en La Vega se identificaron tres QTLs (qLV_1, qLV_2 y qLV_3) distribuidos en dos grupos de ligamiento (LG5 y LG15) (Tabla 2a). A partir de los datos obtenidos en la localidad de Arauca se identificaron igualmente tres QTLs (qAr_1, qAr_2 y qAr_3) distribuidos en dos grupos de ligamiento diferentes (LG 3 y LG13) (Tabla 2b).
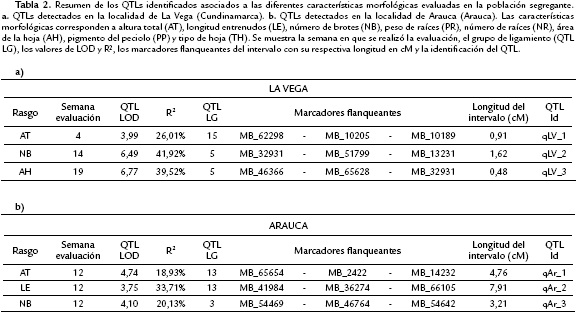
De las características que fueron evaluadas a lo largo del tiempo (AT, LE y NB), fueron identificados dos QTLs (qLV_1 y qLV_2) para La Vega (Tabla 2a) y tres QTLs (qAr_1, qAr_2 y qAr_3) para los datos obtenidos en Arauca (Tabla 2b). En cuanto a la relación de QTLs asociados al mismo rasgo en diferentes ambientes, no se encontró coincidencia entre los QTLs identificados en La Vega y en Arauca. Sin embargo, se observaron dos QTLs (qLV_2 y qLV_3) que se encuentran adyacentes y tienen un marcador en común (MB_32931) los cuales están involucrados en la variación fenotípica de los rasgos NB y AH. Igualmente dos QTLs (qAr_1 y qAr_2) localizados en el grupo de ligamiento 13 están explicando las características de AT y NB (Tabla 2).
Altura de la planta (AT)
Para la característica AT en La Vega se asoció un solo QTL (qLV_1) que explica la variación del fenotipo en el inicio del desarrollo (semana cuatro) en un 26,01 %. Este QTL tiene un intervalo que cubre una región de 0.91 cM. En Arauca igualmente se asoció un QTL (qAr_1) para la segunda semana de evaluación en el grupo de ligamiento 13 el cual cubre una región correspondiente a 4,76 cM. Este QTL explica el 18,93 % de la variación de esta característica.
Longitud entrenudos (LE)
Para LE en La Vega no se detectó ningún QTL. En Arauca se identificó un QTL (qAr_2) que se localizó en el grupo de ligamiento 13 en un intervalo de 7,91 cM. Sobresale este QTL ya que tiene uno de los valores de varianza explicada más alta (33,1 %). Sin embargo no se observó relación alguna entre QTLs que estuvieran presentes entre ambientes ni entre semanas de evaluación.
Número de brotes (NB)
El análisis de QTLs a partir de los datos de NB permitió identificar dos QTLs. El primero (qLA_2) fue detectado a partir de los datos obtenidos en La Vega en la semana 14, el cual se encuentra ubicado en el grupo de ligamiento 5 y cuyo valor de R2 fue el más alto encontrado en este estudio (41,92 %) y el cual abarca una longitud de 1,6 cM. En Arauca en la semana 12 se identificó un QTL (qAr_3) que explica el 20,13 % de la variación, se ubica en el grupo de ligamiento 3 en una región de 3,21 cM de longitud.
Peso de raíces (PR) y número de raíces (NR)
No se logró asociar ningún QTL con los datos obtenidos para estos caracteres en ninguna localidad.
Tamañ o de la hoja (AH)
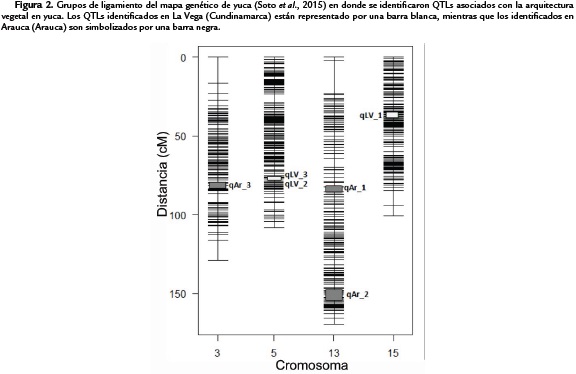
AH solo se evaluó en La Vega y se asoció con un QTL (qLV_3) que explica aproximadamente el 39,52 % de la variación fenotípica de este rasgo. Este QTL se ubica en el grupo de ligamiento 5 y es el que presenta el intervalo más pequeno (0,48 cM) de los QTLs identificados en este estudio. Es interesante resaltar que las regiones comprendidas por los marcadores flanqueantes de los QTLs para los caracteres NB y AH en La Vega (qLV_2 y qLV_3) son adyacentes en el LG5 y son los que tienen valores de varianza explicada más altas (41,92 y 39,52 % respectivamente) con una longitud total de 2,1cM para los dos QTLs (Fig. 2).
Pigmentación del peciolo (PP) y tipo de hoja (TH)
PP y TH fueron evaluados únicamente en La Vega y no se relacionó con ningún QTL.
DISCUSIÓN
La arquitectura de la planta ha sido identificada como un factor importante que subyace a las bases fisiológicas y abren una oportunidad para incrementar el rendimiento de los cultivos. Reportes de rasgos morfológicos han relacionado la arquitectura con el rendimiento de la planta (CIAT, 1975; Lian and Cock, 1979a; Lian and Cock, 1979b; Cock, 1985; Cock and El-Sharkawy, 1988; Iglesias et al., 1994; Hunt et al., 1997; Lenis et al., 2006), la cual es definida por la naturaleza y la disposición relativa de cada una de sus partes. La arquitectura vegetal es el resultado de la expresión equilibrada entre procesos de crecimiento endógenos y las restricciones exógenas ejercidas por el medio ambiente (Barthélémy y Caraglio, 2007).
Se ha encontrado relación entre rasgos morfológicos o de arquitectura vegetal con rasgos de rendimiento. Por ejemplo, el tamano de la planta en papa tiene una correlación positiva significativa con el rendimiento (Maity y Chatteriee, 1977), así como con el número de tubérculos por planta (Alcivar et al., 2007). Igualmente en soya se han relacionado QTLs involucrados en rendimiento de semilla con la altura de la planta (Alcivar et al., 2007). Por otra parte, Perez et al. (2010) mencionan la relación entre el tamano y forma de la hoja en especies de interés agronómico con la capacidad fotosintética repercutiendo no solo en el rendimiento sino también en la respuesta a estrés, resistencia a patógenos y calidad.
Estudios realizados en otros cultivos han identificado y correlacionado igualmente importantes QTLs (y en consecuencia factores genéticos) con rendimiento. En arroz (Oryzae sativa L.) por ejemplo, se han asociado QTLs con el rendimiento, los cuales también estarían explicando la variación de varios rasgos fenotípicos (Saikumar et al., 2014). También en arroz se identificaron QTL mayores asociados con rendimiento del grano que varían su efecto (18,7 % a 31,2 %) de acuerdo a las condiciones ambientales (estrés hídrico). Algunos de ellos co-localizan con otros rasgos como altura de la planta y tamano de panícula (Saikumar et al., 2014). En yuca, se ha encontrado relación entre rasgos morfológicos y rasgos de productividad (Okogbenin y Fregene, 2003).
De esta manera identificar regiones genómicas y posteriormente los genes subyacentes implicados en la arquitectura vegetal permitirían establecer estrategias de mejoramiento genético para lograr un mayor rendimiento en las plantas.
Desde esta perspectiva, este estudio presenta una primera aproximación en la identificación de regiones genómicas asociadas a la arquitectura vegetal en yuca. A partir de la fenotipificación de ocho caracteres morfológicos en la población F1 del cruce entre las variedades elite TMS 30572 (parental femenino) y CM 2177-2 (parental masculino) en dos ambientes diferentes se lograron identificar seis posibles QTLs asociados a los rasgos evaluados. Para ello se tomó como punto de partida el nuevo mapa de ligamiento producido por Soto et al. (2015).
Un mapa de ligamiento necesita < 10-20 cM de separación entre marcadores en cada uno de los grupos para la identificación y localización de QTLs (Sun et al.,2009). El mapa de ligamiento utilizado en este estudio presenta una distancia promedio entre marcadores de 2,5 cM, lo cual es adecuado para localizar QTLs y coincide en cuanto a número de grupos de ligamiento con el número de cromosomas (18) reportados para yuca (Soto et al., 2015).
Paterson et al. (1991) indica que solo QTLs con efectos suficientemente altos pueden ser identificados en un cruce particular, mientras que aquellos con efectos menores pueden pasar desapercibidos. Este mismo estudio indica también que algunos QTLs pueden aparecer con efectos pequeños ya que dependen de la interacción con otros loci. Estos bajos valores pueden también indicar que los rasgos asociados a estos QTLs pueden no segregarse en la población experimental, lo que dificulta su detección (Shrimpton, 1988; Paterson et al., 1991). Teniendo esto en cuenta, los seis QTLs identificados en este estudio muestran valores de R2 por encima de 18 % hasta 41,92 %. De estos, tres se identificaron en La Vega (qLV_1, qLV_2 y qLV_3) (Tabla 2a) para los rasgos AT, NB y AH con valores de LOD por encima de 3.0. Los tres restantes se identificaron en Arauca (qAr_1, qAr_2 y qAr_3) (Tabla 2b) para AT, LE y NB igualmente con valores de LOD por encima de 3,0. Estos QTLs pueden ser considerados como mayores o primarios. En general se lograron detectar solo QTLs con efectos importantes, mientras que los QTLs que pudiesen tener efectos menores o secundarios no se identificaron, seguramente debido a la astringencia en los parámetros asignados en el programa de análisis empleado. Sin embargo, esto no significa necesariamente que tengan menor importancia en la expresión del rasgo.
En La Vega se identificaron tres QTLs asociados a tres de los ocho rasgos evaluados y en Arauca se identificaron igualmente tres QTL asociados a tres rasgos de cinco evaluados en esta localidad. Para las características evaluadas a lo largo del tiempo (AT, LE y NB) en ningún caso se encontró un efecto constante de un mismo QTLs sobre un mismo rasgo a lo largo del tiempo. Es posible que el control en la expresión de estos rasgos se ejerza por diferentes QTLs y/o genes durante los diferentes estadios de desarrollo de la planta. Los QTLs qLV_2 y qLV_3 que afectan dos rasgos diferentes, NB y AH, fueron ubicados adyacentemente lo que sugiere que esta región cromosómica de aproximadamente 2,1cM (Fig. 2) puede poseer varios genes fuertemente ligados que afectan estos dos caracteres. Adicionalmente estos QTLs presentan los valores de R2 más altos (41,92 % y 39,52 %) así como los mejores valores de LOD (6,49 y 6,77 respectivamente) lo cual le da mayor robustez estadística al umbral elegido. Se puede afirmar por esto que el factor ambiental puede ejercer un efecto menor sobre el componente genético para estos dos caracteres tal como ha sido descrito anteriormente (Okogbening y Fregene, 2007). Los QTLs que son estables en un amplio espectro de entornos son los predilectos para el mejoramiento genético ya que en ellos el factor genético juega un papel mayor, y es éste el que es susceptible de ser modificado. En consecuencia, estas dos características son promisorias por tener alto potencial para ser tenidas en cuenta en programas de mejoramiento.
En yuca son varios los estudios que han reportado QTLs asociados con caracteres morfológicos. Boonchanawiwat et al. (2011) identificaron siete QTLs en cuatro grupos de ligamiento para altura de la planta y cinco QTLs en cinco grupos de ligamiento para la altura del primer brote en dos ambientes diferentes. Sin embargo otros estudios han reportado más de un centenar de QTLs, no solamente asociados con arquitectura sino también con productividad, contenido de carotenos y cantidad de proteína (Okogbenin y Fregene, 2003; Okogbenin y Fregene, 2006; Okogbenin y Fregene, 2007). En general estos reportes muestran que se han encontrado gran cantidad de QTLs, no solo primarios con valores de R2 altos sino una gran mayoría de QTLs con efectos menores. En el presente estudio se logró identificar únicamente seis QTLs partiendo de la base de un mapa significativamente mas denso en marcadores y con un paquete informático que ha venido siendo mejorado. A futuro será interesante poder unificar la información generada en este estudio con los previos para tener no solo un mapa integrado sino un repertorio completo de QTLs de arquitectura y productividad en yuca, que incluye QTLs mayores y menores así como QTLs comunes identificados en diferentes estudios.
Comparando los QTLs identificados para cada rasgo en cada uno de los ambientes se observó una discrepancia entre los QTLs encontrados entre las dos localidades en las cuales se realizó la fenotipificación. Por ejemplo, AT se asoció con un QTL en La Vega (qLV_1) y con uno diferente en Arauca, incluso a tiempos diferentes (qAr_1). Una situación similar se presentó para NB (Tabla 2a y 2b). Este resultado era esperado ya que esta característica está condicionada en gran medida por el medio ambiente (Suarez y Mederos, 2011). Dentro de los factores ambientales que pueden estar determinando la expresión de estos rasgos se encuentran por ejemplo el tipo y calidad del suelo, así como las condiciones climáticas que son diferentes en las dos localidades. Como se mencionó anteriormente La Vega y Arauca se encuentran a altitudes diferentes y poseen temperaturas promedios distintas. Se ha reportado que los genotipos o variedades que se desarrollan exitosamente en un medio, no se comportan necesariamente de igual forma en lugares con entornos diferentes (Suarez y Mederos, 2011). Estas condiciones edafoclimaticas contrastantes pueden explicar en gran parte los diferentes QTLs identificados en las dos regiones.
En otras especies de interés agronómico se han determinado características similares a PP y TH en donde el control genético es el responsable de la variación del rasgo y es independiente del medio ambiente. Tal es el caso por ejemplo de estudios en leguminosas (L. Japonicus) donde se pudieron identificar cuatro QTLs que coinciden en dos tiempos diferentes de evaluación para las características largo y ancho del lóbulo medio de la hoja (Choi et al., 2004). En tomate (Solanum lycopersicum L.) también se han identificado QTLs relacionados con el tamano y forma de la hoja donde se observa un bajo nivel de coincidencias entre QTLs que expliquen el mismo rasgo, indicando que el control genético para estos rasgos es predominantemente independiente (Frary et al., 2003; Frary et al., 2004). En este estudio no se obtuvo ninguna relación de los rasgos PP y TH evaluados en La Vega con ningún QTL que permitiera confirmar que estos rasgos son exclusivamente gobernados por el componente genético.
A futuro, contar con una población segregante conformada por más individuos recombinantes y disponer de un mayor número de marcadores moleculares permitiría mejorar la resolución del mapeo y en consecuencia se podría obtener una mejor identificación de los QTLs. En este trabajo si bien se identificaron varios QTLs, algunos de ellos se encuentran en intervalos bastante grandes (7,91 cM) lo que hace difícil la identificación de los genes candidatos implicados en el control de las características evaluadas. Al reducir este intervalo, incrementado la población y la frecuencia de recombinantes, así como aumentando el número de marcadores, se podría identificar más fácilmente los posibles genes responsables de los QTLs identificados.
La posibilidad de identificar QTLs en diferentes ambientes, como se presenta en este trabajo, permitiría identificar regiones genómicas que pueden ejercer un efecto, así éste sea pequeno, independiente de las condiciones ambientales. Por lo tanto, la combinación de varios QTLs que actúen en una amplia gama de entornos en un solo genotipo, permitiría alcanzar rendimientos aceptables incluso en condiciones ambientales muy diferentes (Paterson et al., 1991). Este estudio sugiere que QTLs relacionados con la arquitectura de la planta pueden ser empleados a través de programas de mejoramiento para incrementar la productividad de la yuca.
CONCLUSIONES
La generación de un mapa genético de yuca de alta densidad y la fenotipificación de diferentes características relacionadas con la arquitectura vegetal evaluadas en dos regiones geográficas diferentes permitió la identificación de seis QTLs. Se detectaron tres QTLs en La Vega asociados con altura total, número de brotes y área de la hoja. En Arauca los QTLs identificados estuvieron asociados con altura total, longuitud de entrenudos y número de brotes. La posterior identificación de los genes presentes en estas regiones genómicas contribuirá a desarrollar programas de mejoramiento genéticos encaminados a modificar la arquitectura vegetal con miras a incrementar el rendimiento de la yuca.
AGRADECIMIENTOS
Especial agradecimiento a la Sede Orinoquia de la Universidad Nacional de Colombia y a Lisímaco López, propietario de la Finca Los Bugambiles en La Vega, por la disponibilidad del terreno y mantenimiento de las plantas de yuca. Agradecimientos a Luis Augusto Becerra y Fernando Calle del CIAT por la disponibilidad del material vegetal. Los autores agradecen a la DIB de la Universidad Nacional de Colombia por apoyo a Ruben Mora a través de la convocatoria "Programa nacional de semilleros de investigación, creación e innovación 2013-2015) y a COLCIENCIAS por la beca de doctorado convocatoria 528, de la cual Johana Soto es beneficiaria.
REFERENCIAS
Akano A, Dixon A, Mba C, Barrera E, Fregene M. Genetic mapping of a dominant gene conferring resistance to cassava mosaic disease. Theor Appl Genet. 2002;105:521-525. Doi:10.1007/S00122-002-0891-7. [ Links ]
Alcaldía Municipal de Arauca. Plan Básico de Ordenamiento Territorial del Municipio de Arauca, Componente General. Arauca, Colombia: Gaceta Municipal; 2000.23 p. [ Links ]
Alcivar A, Jacobson J, Rainho J, Meksem K, Lightfoot D. Genetic analysis of soybean plant height, hypocotyl and internode lengths. Agric Food Environ Sci.2007;1(1):1-16. [ Links ]
Barthélémy D, Caraglio Y. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Ann Bot. 2007;99(3):375-407. Doi:10.1093/aob/mcl260. [ Links ]
Boonchanawiwat A, Sraphet S, Boonseng O, Lightfoot D, Triwitayacorn T. QTL underlying plant and first branch heigh in cassava (Manihot esculenta Crantz). Field Crops Res. 2011;121(3):343-349. Doi:10.1016/j. fcr.2010.12.022. [ Links ]
Cerón J, Sahagún J. Estimating QTL biometrics parameters in F2 populations: a new approach. Agrociencia. 2007;41:57-73. [ Links ]
Choi Kk, Munch Jh, Kim Dj, Zhu H, Baek Jm, et al. Estimating genome conservation between crop and model legume species. Proc Natl Acad Sci USA. 2004;101(43):15289-15294. Doi:10.1073/pnas.0402251101. [ Links ]
CIAT (Centro Internacional de Agricultura tropical). Cassava production systems. Cali, Colombia: Annual report; 1975. 260 p. [ Links ]
Cock JH. Cassava: New Potential for a Neglected Crop. Boulder, Colorado: Westview Press. 1985; 191 p. [ Links ]
Cock JH, EL-Sharkawy MA. Physiological characteristics for cassava selection. Exp Agric. 1988;24:443-448. [ Links ]
Cock JH. La yuca, nuevo potencial para un cultivo tradicional. Cali, Colombia: Centro Internacional de Agricultura Tropical; 1989. 240 p. [ Links ]
FAO. Panorama de la seguridad alimentaria y nutricional de América Latina y el Caribe. Santiago: Oficina Regional para América Latina y el Caribe de FAO; 2013. p. 48. [ Links ]
Ferreira M, Grattapaglia D. Introducción al uso de marcadores moleculares en el análisis genético. Brasilia: Embrapa-Cenargen; 1998. p. 1221. [ Links ]
Frary A, Doganlar S, Daunay M, Tanksley S. QTL analysis of morphological traits in eggplant and implications for conservation of gene function during evolution of solanaceous species. Theor Appl Genet 2003; 107(2):359-370. Doi:10.1007/s00122-003-1257-5. [ Links ]
Frary A, Fritz A, Tanksley D. A comparative study of the genetic bases of natural variation in tomato leaf, sepal, and petal morphology. Theor Appl Genet. 2004;109:523-533. Doi:10.1007/s00122-004-1669-x. [ Links ]
Fregene M, Angel F, Gomez R, Rodriguez F, Chavarriaga P, et al. A molecular genetic map of cassava (Manihot esculenta Crantz). Theor Appl Genet. 1997;95:431-441. [ Links ]
Gil J, Buitrago A. La yuca en la alimentación animal. La yuca en el tercer milenio. Publicación CIAT, Cali, Colombia. 1990;85:26-26. [ Links ]
Hunt A, Wholey W, Cock J. Growth physiology of cassava (Manihot esculenta Crantz). Field Crops Abstracts. 1997;30:77-91. [ Links ]
Ideam (Instituto de Hidrología, Meteorología y Estudios Ambientales). Sistemadeinformación ambiental,registros estacionales. Estación Sabaneta (Cundinamarca). 2002. [ Links ]
Iglesias C, Calle F, Hershey C, Jaramillo G, Mesa E. Sensitivity of cassava (Manihot esculenta Crantz) clones to environmental changes. Field Crops Res. 1994;36:213-220. Doi: 10.1016/0378-4290(94)90113-9. [ Links ]
Jorge V, Fregene M, Velez C, Duque M, Tohme J, Verdier V. QTL analysis of field resistance to Xanthomonas axonopodis pv. manihotis in cassava. Theor Appl Genet, 2001;102:564-571. Doi:10.1007/s001220051683. [ Links ]
Lenis JI, Calle F, Jaramillo G, Perez JC, Ceballos H, Cock J. Leaf retention in cassava productivity. Field Crops Res. 2006;95(2-3):126-134. Doi:10.1016/j.fcr.2005.02.007. [ Links ]
Lian TS, Cock JH. Branching habit as a yield determinant in cassava. Field Crops Res. 1979a;2:281-289. Doi:10.1016/0378-4290(79)90029-7. [ Links ]
Lian TS, Cock JH. Cassava plant forms and their associated and their associated morphophysiological characters. MARDI Res Bull. 1979b;2:55-69. [ Links ]
Lopez C, Quesada L, Bohorquez A, Duque M, Vargas J, Tohme J, et al. Mapping EST-derived SSRs and ESTs involved in resistance to bacterial blight in Manihot esculenta. Genome. 2007;50:1078-1088. Doi:10.1139/G07-087. [ Links ]
Lopez C. Descifrando las bases moleculares de la resistencia cuantitativa. Acta biol. Colomb. 2011;2:3-14. [ Links ]
Maity S, Chatteriee B. Growth attributes of potato and theirinter-relationship with yield. Potato Res. 1977;20:337-341. Doi:10.1007/BF02362245. [ Links ]
Mba R.E.C., Stephenson P, Edwards K, Melzer S, Nkumbira J, Gullberg U, et al. Simple sequence repeat (SSR) markers survey of the cassava (Manihot esculenta Crantz) genome: towards an SSR-based molecular genetic map of cassava. Theor App Genet. 2001;102:21-31. Doi:10.1007/s001220051614. [ Links ]
Mederos V. Embriogénesis somática en yuca (Manihot esculenta Crantz). (Tesis de Doctorado). Ciego de Ávila, Cuba: Universidad de Ciego de Ávila, Centro de Bioplantas; 2006. 5 p. [ Links ]
Okogbenin E, Fregene M. Genetic mapping of QTLs affecting productivity and plant architecture in a full-sib cross from non-inbred parents in Cassava (Manihotesculenta Crantz). Theor Appl Genet. 2003;107:1452-1462. Doi:10.1007/s00122-003-1383-0. [ Links ]
Okogbenin E, Marin J, Fregene M. An SSR-based molecular genetic map of cassava. Euphytica. 2006;147:433-440. Doi:10.1007/s10681-005-9042-y. [ Links ]
Okogbenin E, Marin J, Fregene M. QTL analysis for early yield in a pseudo F2 population of cassava. Afr J Biotechnol. 2007;7(2):131-138. [ Links ]
Ospina B, Ceballos H. Cassava in the third millennium: Modern production, processing, use, and marketing systems. In: Centro Internacional de Agricultura Tropical (CIAT). Latin American and Caribbean Consortium to support Cassava Research and Development (CLAYUCA). Cali, Colombia: Technical Centre for Agricultural and Rural Cooperation (CTA); 2012. 574 p. [ Links ]
Perez J, Bruna D, Micol J. QTL analysis of leaf architecture. J Plant Res. 2010;123:15-23. Doi:10.1007/s10265-009-0267-z. [ Links ]
Paterson A, Damon S, Hewitt JD, Zamir D, Rabinowitch H, Lincoln E, et al. Mendelian factors underlying quantitative traits in tomato: comparison across species, generations, and environments. Genetics. 1991;127:181-197. [ Links ]
Rabbi I, Hamblin M, Kumar L, Gedil M, Ikpan A, Jannink J, et al. High-resolution mapping of resistance to cassava mosaic geminiviruses in cassava using genotyping-by-sequencing and its implication for breeding. Virus Res. 2014;186:87-96. Doi:10.1016/j.virusres.2013.12.028. [ Links ]
Saikumar S, Kalmeshwer P, Saiharini A, Moohan C, Vineesha O, Padmavathi G, et al. Major QTL for enhancing rice grain yield under lowland reproductive drought stress identified using an O. sativa/O glaberrima introgression line. Field Crops Res. 2014;163:119-131. Doi:10.1016/j. fcr.2014.03.011. [ Links ]
Shrimpton A, Robertson A. The isolation of polygenic factors controlling bristle score in Drosophila melanogaster. II. Distribution of third chromosome bristle effects within chromosome sections. Genetics. 1988;118(3):445-459. [ Links ]
Soto J, Ortiz JF, Perlaza-Jiménez L, Vásquez AX, Lopez-Lavalle LA, Mathew B, et al. A genetic map of cassava (Manihot esculenta Crantz) with integrated physical mapping of immunity-related genes. BMC Genomics. 2015;16:190. Doi: 10.1186/s12864-015-1397-4. [ Links ]
Suarez L, Mederos V. Apuntes sobre el cultivo de yuca (Manihot esculenta Crantz) tendencias actuales. Cultivos Tropicales. 2011;32(3):27-35. [ Links ]
Sun W, Zhang Y, Le W, Zhang H. Construction of a genetic linkage map and QTL analysis for some leaf traits in pear (Pyrus L.). Front Agric China. 2009;3:67-74. Doi:10.1007/s11703-009-0013-2. [ Links ]
