










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.21 no.2 Bogotá May./Aug. 2016
https://doi.org/10.15446/abc.v21n2.48883
Doi: http://dx.doi.org/10.15446/abc.v21n2.48883
COMPARATIVE GROWTH AND BIOCHEMICAL COMPOSITION OF FOUR STRAINS OF Nostoc AND Anabaena (CYANOBACTERIA, NOSTOCALES) IN RELATION TO SODIUM NITRATE
Comparación del crecimiento y Composición Bioquímica de cuatro cepas de Nostoc y Anabaena (Cyanobacteria, Nostocales) en relación con el nitrato de sodio
Néstor ROSALES LOAIZA1, Patricia VERA1, Cateryna AIELLO-MAZZARRI2, Ever MORALES1.
1 Laboratorio de Microorganismos Fotosintéticos, Departamento de Biología, Facultad Experimental de Ciencias, Universidad del Zulia. Maracaibo, Venezuela.
2 Laboratorio de Fermentaciones Industriales, Departamento de Ingeniería Bioquímica, Facultad de Ingeniería, Universidad del Zulia. Maracaibo, Venezuela.
For correspondence. nestoralgae@gmail.com
Received: 4th February 2015, Returned for revision: 23th July 2015, Accepted: 26th October 2015. Associate Editor: Rafael Riosmena Rodríguez.
Citation / Citar este artículo como: Rosales Loaiza N, Vera P, Aiello-Mazzarri C, Morales E. Comparative growth and biochemical composition of four strains of Nostoc and Anabaena (Cyanobacteria, Nostocales) in relation to sodium nitrate. Acta biol. Colomb. 2016;21(2):347-354. doi: http://dx.doi.org/10.15446/abc.v21n2.48883
ABSTRACT
Nitrogen concentration is an essential parameter in cyanobacterial cultures to produce enriched biomass with biotechnological purposes. Growth and biochemical composition of Nostoc LAUN0015, Nostoc UAM206, Anabaena sp.1 and Anabaena sp.2 were compared at 0, 4.25, 8.5 and 17 mM NaNO3. Cultures under laboratory conditions were maintained for 30 days at a volume of 500 mL. Anabaena sp.1 yielded the highest value of dry mass of 0.26 ± 2.49 mg mL-1 at 8.5 mM NaNO3. For chlorophyll, phycocyanin and phycoerythrin, maximum values were achieved at 17 mM NaNO3 with 18.09 ± 1.74, 102.90 ± 6.73 and 53.47 ± 2.40 μg mL-1, respectively. Nostoc LAUN0015 produced its maximum value of protein 644.86 ± 19.77 μg mL-1, and 890 mg mL-1 of carbohydrates in the absence of nitrogen. This comparative study shows that the most efficient strain for the production of protein, carbohydrates and lipids in diazotrophic conditions corresponded to Nostoc LAUN0015. However, Anabaena sp.1 and Anabaena sp.2 required high nitrogen concentrations to achieve higher values of metabolites, comparing with Nostoc strains. Nitrogen dependence for the production of pigments and high protein production in strains of Anabaena and in diazotrophic conditions for Nostoc was demonstrated. Nostoc can be cultured under nitrogen deficiency and Anabaena in sufficiency, for biomass production enriched with proteins and carbohydrates.
Keywords: Anabaena, biochemical composition, biomass, culture, nitrate, Nostoc.
RESUMEN
La concentración de nitrógeno constituye un parámetro esencial en cultivos de cianobacterias para la producción de biomasa enriquecida con fines biotecnológicos. Se comparó el crecimiento y composición bioquímica de las cepas Nostoc LAUN0015, Nostoc UAM206, Anabaena sp.1 y Anabaena sp.2 a 0, 4,25; 8,5 y 17 mM NaNO3. Los cultivos en condiciones de laboratorio fueron mantenidos durante 30 días a un volumen de 500 mL. En masa seca, Anabaena sp.1 obtuvo el mayor valor, con 2,49 ± 0,26 mg mL-1 a 8,5 mM NaNO3. Para clorofila, ficocianina y ficoeritrina, los máximos se alcanzaron a 17 mM NaNO3 en Anabaena sp.1, con 18,09 ± 1,74; 102,90 ± 6,73 y 53,47 ± 2,40 μg mL-1, respectivamente. Nostoc LAUN0015 produjo su máximo valor de proteínas de 644,86 ±19,77μg mL-1, y alrededor de 890 μg mL-1 de carbohidratos en ausencia de nitrógeno. El estudio comparativo indica que la cepa más eficiente para la producción de proteínas, carbohidratos y lípidos, en condiciones diazotróficas, correspondió a Nostoc LAUN0015. En cambio, las cepas de Anabaena sp.1 y sp.2 requieren de elevadas concentraciones de nitrógeno para alcanzar los mayores valores de metabolitos, respecto a las cepas de Nostoc. Se demuestra la dependencia de nitrógeno para la producción de los pigmentos y la alta producción proteica en las cepas de Anabaena y en condiciones diazotróficas para Nostoc. Esta última puede ser cultivada bajo una deficiencia de nitrógeno y Anabaena con suficiencia para la producción masiva de biomasa enriquecida con proteínas y carbohidratos.
Palabras clave: Anabaena, biomasa, composición bioquímica, cultivo, nitrato, Nostoc.
INTRODUCTION
Cyanobacteria or blue-green algae are photosynthetic microorganisms that can be used to produce high-value compounds (Vincent, 2009). These include high protein content; capacity to synthesize all amino acids (and provide the essential ones to humans and animals); presence of carbohydrates composed of starch, glucose, sugars and non-digestible polysaccharides (agar, carrageenan and alginate); lipids in the form of glycerol and fatty acids of the ω-3 and ω-6 families; and a valuable content of many essentials vitamins, minerals and antioxidant substances (Harun et al., 2010). With this biochemical composition is not a surprise that this microorganism can be used as a food source for animal and humans (Gantar and Svirčev, 2008; Cunningham and Joshi, 2010).
Dried microalgal biomasses typically contain 46-63 % protein, 8-17 % carbohydrates, and 4-22 % lipids, as well as a wide range of vitamins and other biologically active substances such as bioactive peptides and pigments (Gantar and Svirčev, 2008).
Nostoc, an edible blue-green alga, is a cyanobacterium that has been grown and cultivated for medicinal uses for centuries (Gantar and Svirčev, 2008). Recent studies have indicated that Nostoc contains cryptophycin, a compound that inhibits cancer cell growth, as well as anti-viral compounds (Cunningham and Joshi, 2010; Sharma et al., 2011).
Filamentous cyanobacteria Nostoc, Spirulina, Arthrospira, Anabaena, Aphanizomenon, Rivularia, and many others are particularly attractive for the production of high quality biomass, because they represent a source of protein and a variety of chemicals and pharmaceuticals (Gantar and Svirčev, 2008).
Despite the great interest in culturing microalgae and cyanobacteria, it is estimated that only 10 % of existing species have been studied in order to know their physiology and their potential as producers of biocompounds, especially in relation to tropical strains (Rasmussen and Morrissey, 2007). Therefore, the aim of this work was to evaluate the gross biochemical characteristics of two stains of Nostoc, and two strains of Anabaena, cultivated in different nitrogen concentrations in order to prove their potential use as food or supplement, especially as a protein source.
MATERIALS AND METHODS
Filamentous heterocystous cyanobacteria studied were: (1) Nostoc LAUN 0015, isolated form a humid environment in Bogota, Colombia; (2) Nostoc UAM206, isolated from an inundated rice field in Valencia, Spain; (3) Anabaena sp.1, from activated sludge of a treatment plant; and (4) Anabaena sp.2 from an oil pit in Venezuela.
Cultures by triplicate were maintained in 1 L flasks with 500 mL culture medium composed of sterilized tab water enriched BG-11 culture medium (Rippka et al., 1979). Flasks were inoculated to an absorbance of 0.08 at 750 nm, and incubated at 29 ± 2 °C under a 12h-light/12h-dark cycle with a light intensity of 156 mmol of photons s-1 m-2 and constant aeration of 4.95 ± 0.03 mL s-1.
Growth was evaluated at four different sodium nitrate concentrations by using 0, 4.25, 8.5 and 17 mM NaNO3, equivalent to 0, 25, 50 and 100 % of nitrate concentration present in BG-11 culture medium.
Growth was determined by turbidity (OD750 nm). Biomass was harvested by centrifugation to 10 x103 g for 10 min. Frozen biomass, stored to -20 ºC, were used for all the biochemical analyses, except for pigments and dry weight, for which fresh biomass samples were used. The protein content was determined by the modified Folin-Lowry method (Herbert et al., 1971). Pigments were extracted in methanol (99 %) at 4 °C overnight and measured by spectrophotometric methods (Strickland and Parsons, 1972; Marker and Jinks, 1982). Carbohydrates were measured by the phenol-sulfuric acid method (Kochert, 1978). Dry weight was determined using a Millipore© filtration system, with 0.45 µm fiberglass filter, by the method of Utting (1985).
Statistical analyses were performed with SPSS 15.0, using analysis of variance (ANOVA) and Sheffé's test to examine differences in cellular density and biochemical composition between different nutrient concentrations.
RESULTS
For biomass production there were different patterns for every strain of cianobacteria (Table 1). Nostoc strains produced higher biomass values to lower sodium nitrate concentrations, with maximum of 1.32 ± 0.12 and 1.56 ± 0.16 mg mL-1, at 0 mM NaNO3, for Nostoc LAUN0015 and Nostoc UAM206, respectively (p < 0.05). Anabaena strains enhanced biomass production increasing nitrogen concentration until 8.5 mM NaNO3. Anabaena sp.1 reached the highest biomass production of 2.49 ± 0.26 mg mL-1 (p < 0.05).
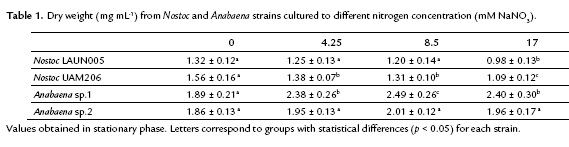
Chlorophyll a and carotenoids production from filamentous cyanobacteria to different nutrient concentrations are shown in Figure 1. Production of these pigments seems not to be influenced for nitrogen concentration, except for Nostoc LAUN0015. This strain showed a diminishing of pigment production with an increase of nitrogen. Maximal production for Nostoc LAUN0015 was obtained at 0 mM NaNO3 with 2.56 ± 0.28 and 0.84 ± 0.04 µg mL-1 for chlorophyll a and carotenoids, respectively (p < 0.05).
The remaining strains showed higher levels of pigments with increasing nitrogen concentration. Nostoc UAM206 obtained 2.37 ± 0.26 and 2.46 ± 0.26 µgmL-1 for chlorophyll a at 4.25 and 8.5 mM NaNO3, respectively; with no significant differences (p > 0.05). In contrast, carotenoids reached their peak between 8.5 and 17 mM NaNO3 with 1.21 ± 0.12 and 1.13 ± 0.11 µg mL-1, respectively (p > 0.05).
Anabaena strains obtained their higher values of liposoluble pigments to the highest nitrogen concentration. Anabaena sp.1 produced 1.74 ± 18.09 and 2.99 ± 0.15 µg mL-1 of chlorophyll a and carotenoids, respectively. Anabaena sp.2 achieved 15.04 ± 1.33 µg mL-1 of chlorophyll a and 2.44 ± 0.32 µg mL-1 of carotenoids. These values were obtained at 17 mM NaNO3, although there was no significant difference (p > 0.05) with respect to treatments 4.25 and 8.5 mM NaNO3.
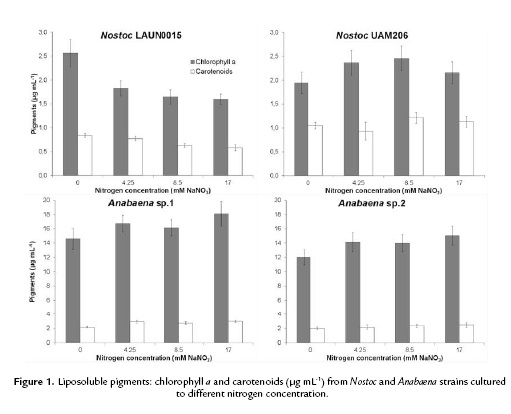
Figure 2 shows production of hydrosoluble pigments (phycocyanin and phycoerythrin). Nostoc LAUN0015 (Fig. 2) showed for phycoerythrin, the same trend observed in the liposoluble pigments, with the highest value of 23.68 ± 2.33 µg mL-1 in the absence of nitrogen (p < 0.05). Highest values of phycocyanin, 14.04 ± 0.83 and 13.97 ± 0.23 µg mL-1, were obtained at 0 and 4.25 mM NaNO3, respectively; with no significant differences (p > 0.05).
Nostoc UAM206 showed statistical homogeneity (p > 0.05) for phycocyanin, phycoerythrin production from 0 to 8.5 mM NaNO3. Maximum values of phycocyanin and phycoerythrin 11.55 ± 1.05 and 13.10 ± 1.04 µg mL-1 were obtained at 8.5 and 0 mM NaNO3.
Anabaena strains showed an increase in hydrosoluble pigments with nitrogen concentration. Anabaena sp.1 (Fig. 2) reached the highest values at 8 and 17 mM NaNO3 with 94.31 ± 4.82 and 102.90 ± 6.73 µg mL-1 and 49.83 ± 2.54 and 53, 47 ± 2.40 µg mL-1 for phycocyanin and phycoerythrin, with no significant difference (p > 0.05). Meanwhile, Anabaena sp.2 produced 85.46 ± 9.95 and 45.27 ± 3.49 µg mL-1 for phycocyanin and phycoerythrin to a nitrogen concentration of 8.5 mM NaNO3 (p < 0.05).
Nostoc LAUN0015 produced highest phycocyanin and phycoerythrin values at 0 and 4.25 mM NaNO3, compared to Nostoc UAM206. Anabaena strains produced higher content of these pigments, compared to Nostoc. Highest phycocyanin and phycoerythrin production in Anabaena sp.1, at 17 mM NaNO3, were 7.33 and 2.26 times higher than the highest productions achieved for Nostoc LAUN0015, growing with no nitrogen. The production order for phycobiliproteins was: Anabaena sp.1 > Anabaena sp.2 > Nostoc LAUN0015 > Nostoc UAM206.
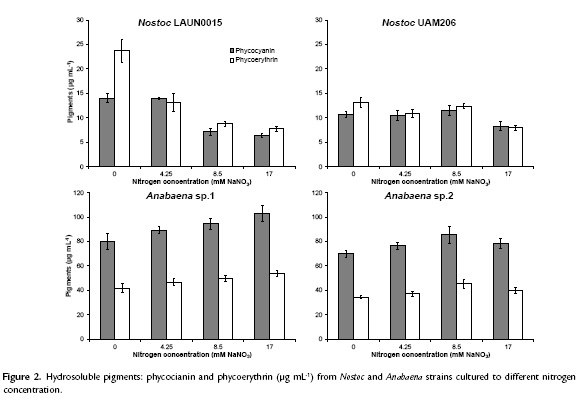
In general, protein production seemed to be enhanced at medium to high nitrogen concentrations (Fig. 3). On the other hand, Nostoc LAUN0015 achieved maximum value in the absence of nitrogen with 686 ± 19.77 mg mL-1(p < 0.05). Maximal values for protein concentration for Nostoc UAM206, Anabaena sp.1 y Anabaena sp.2 of 442.14 ± 17.09, 897.64 ± 46.94 and 758.13 ± 11.53 mg mL-1were achieved at 8.5 mM NaNO3 (p < 0.05).
Carbohydrate production for filamentous cyanobacteria under study at different nitrogen concentrations were summarized in Table 2. Nostoc and Anabaena strains presented two different patterns. Nostoc seems to accumulate carbohydrates under absence of nitrogen in the culture, while Anabaena accumulates carbohydrates in high nitrogen concentrations.
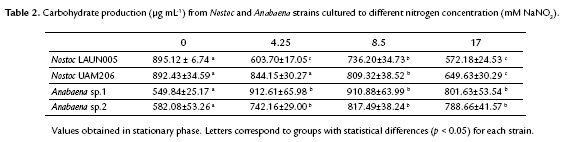
Carbohydrate maximum production for Nostoc LAUN0015 and Nostoc UAM206 were 895.12 ± 6.74 to 893.43 ± 34.59 mg mL-1 at 0 mM NaNO3, with no statistical differences (p > 0.05). For Anabaena maximum values were reached at 4.25 and 8.5 mM NaNO3 with 912.61 ± 65.98 and 817.49 ± 38.24 mg mL-1 for Anabaena sp.1 and Anabaena sp.2, respectively. These values did not differ significantly with respect to the treatments where nitrogen was added to the culture (p > 0.05), but did respect to the absence of nitrogen (p < 0.05).
DISCUSSION
In Synechococcus, Synechocystis, Oscillatoria agardhii and O. redekei, a positive correlation between nitrogen concentration and growth, using low concentrations, from 0 to 2.5 mM NaNO3 have been described (Foy, 1993; Hu et al., 2000). Increased biomass production from Nostoc strains at low nitrogen concentrations seems not to be related neither with increased cellular production nor intracellular accumulation of metabolites, but with the increase in exopolysaccharide production; which is very common in cyanobacteria strains under nutritional stress conditions (Otero and Vincenzini, 2004). Because there is no simple method to determine dry weight from free cells, with no capsular polysaccharide, it was included within the value of dry weight.
Nostoc and Anabaena strains were capable of diazotrophic growth, which means that these strains do not need a nitrogen source in the culture medium for growth (Whitton and Potts, 2012). Nitrogen-fixing cyanobacteria are widespread among filamentous heterocyst-forming genera, such as Anabaena, Nostoc, Rivularia, Stigonema and Scytonema, among others (Tsygankov, 2007). Its ability to grow at different nitrogen concentrations shows its physiological versatility to adapt to diverse environments, even when nitrogen is limiting or absent. Therefore, its growth at low nitrogen concentrations is supported by the nitrogen-fixing process (Loreto et al., 2003).
Similar studies has been verified that cyanobacteria grow better with higher levels of nitrogen (Jonte et al., 2003; Loreto et al., 2003; Rosales et al., 2006; Fuenmayor et al., 2009; Rosales Loaiza and Morales, 2013). However, it should be noted that this characteristic is not present in all cyanobacterial cultures. Growth differences, in relation to environmental conditions such as light, temperature, and pH, can be found even within strains of the same specie (Jonte et al., 2003; Vonshak and Torzillo, 2004).
Results from chlorophyll a provide important tool that helps to quantify the growth of a phototrophic organism. There is considerable evidence that supports the fact that the amount of chlorophyll is positively correlated with cell density or biomass (Serpa and Calderon, 2006). Results showed that Anabaena sp. 1 is an excellent source of chlorophyll a, with commercial interest, especially for its antioxidant properties (Lanfer-Marquez et al., 2005). The order of production of both chlorophyll a and total carotenoids was as follows: Anabaena sp.1 > Anabaena sp.2 > Nostoc LAUN0015 > Nostoc UAM206.
Also, it was demonstrated that pigment production is suitable at low and intermediate nitrogen concentrations. In Anabaena, highest value was found at 17 mM NaNO3, but this value was just 1.06 times higher than chlorophyll concentration obtained at 4.25 mM NaNO3; despite the fact that nitrogen concentration was increased three times. This finding has great importance, especially for industrial purposes, because the use of low nitrogen concentrations yields a very good chlorophyll a production, without causing great expenses.
The situation is more evident with Nostoc LAUN0015 which the best result for liposoluble pigments achieved in total absence of nitrogen. Chlorophyll a and carotenoids production was 1.6 and 1.5 times higher under complete absence of nitrogen compared to the highest concentration of 17 mM NaNO3, which verifies the high diazotrophic capacity for growth and pigment production in this strain.
Pigment content depends on the nitrogen source and concentration (Simeunović et al., 2013). In fact, for nitrogen non-fixing strains, the first biomolecule degraded in the process of cellular acclimation under absence of nitrogen are phycobiliproteins (Baier et al., 2001; Simeunović et al., 2013).
Synthesis of pigments, especially phycobiliproteins, is particularly susceptible to environmental influences. In general, the low phycocyanin production in nitrogen-limited cultures obeys to degradation processes in order to mobilize this chromoprotein, for most primary processes such as growth (Lewitus and Caron, 1990); but results showed that at least for Nostoc; nitrogen limited cultures actually produce more phycobiliproteins than non-limited cultures. This can be explain through nitrogen fixation.
Heterocystous cyanobacteria, such as Nostoc and Anabaena, are capable to fix the atmospheric nitrogen, to produce chlorophyll, carotenoids and phycobiliproteins in significant quantities, and that can be seen in the results, especially with Nostoc strains. Therefore, production of these microorganisms represents a metabolic strategy with great biotechnological interest, for being produce under diazotrophic conditions. This non-nitrogen culture conditions, it would produce savings in biomass production, since much of the cost of mineral nutrients (i.e. fertilizers) is in the transport of their mass (Stephens et al., 2012). Nutrient supply constitutes a primary limitation for mass production for food and fuel (Stephens et al., 2012; Acien et al., 2015).
Decrease in carotenoid content in nutrient-limited cultures, suggests that these strains do not accumulate carotenoids under nitrogen deficiency, such as Pseudanabaena, Oscillatoria, Chlorella and Dunaliella (Canto de Loura et al., 1987; Hu, 2004). Previous reports stated an increase in pigment and protein production with high nitrate concentration in Gleotrichia sp. (Pattnaik and Singh, 1978), Merismopedia tenuissima (Konopka and Schnur, 1981), Chroococcidiopsis sp. (Billi and Grilli, 1996), Anabaena sp. PCC 7120 (Loreto et al., 2003) and Synechococcus sp. (Rosales et al., 2006).
Nostoc LAUN0015 seems to improve pigment production in total absence or low nitrogen concentrations in the culture medium. This case seems that nitrogen fixation is a more effective than nitrogen assimilation from the surrounding environment.
Increase in biomass and protein production by increasing nitrogen concentration, as it seen in result of Nostoc and Anabaena, has been widely supported by various reports from Pseudanabaena (Leal et al., 2001), Anabaena (Loreto et al., 2003), Oscillatoria (Saha et al., 2003; Fuenmayor et al., 2009), Chaetoceros (Leonardos and Geider, 2004), Synechococcus (Rosales et al., 2006), Dunaliella (Rosales Loaiza et al., 2007) and Spirulina platensis (Colla et al., 2007).
It has been shown that several environmental factors including nutrients status, light, salinity, among others, not only affect photosynthesis and productivity of algal cells, but also influence the overall metabolic activity and cellular composition (Hu, 2004; Guschina and Harwood, 2009).
Microalgae and cyanobacteria are known to modulate the production of both exopolysaccharides and endopolysaccharides in response to various environmental factors, such as salinity stress, high irradiances, and nitrogen deficiency (Moreno et al., 1998; Hu, 2004). Meanwhile, capsular polysaccharide (PSC) production seems to be influenced by nitrogen absence in the culture medium. Numerous studies also show that lipid accumulation is one of the main responses of microalgae and cyanobacteria in nitrogen limited culture (Arias Peñaranda et al., 2013). Also, carbohydrate accumulation occurs frequently (Hu, 2004).
Nitrogen deficiency stimulates polysaccharide production, as it has been demonstrated in Merismopedia tenuisima (Konopka and Schnur, 1981), Synechococcus (Roux, 1996), Cyanothece sp. (De Philippis et al., 1998), Anabaena (De Philippis and Vincenzini, 1998; Singh and Das, 2011), Nostoc (Otero and Vincenzini, 2003; Otero and Vincenzini, 2004; Singh and Das, 2011) and Oscillatoria (Jindal, 2011).
CONCLUSIONS
This comparative study indicates that protein, carbohydrates and liposoluble pigments production increases under diazotrophic conditions for Nostoc LAUN0015. Instead, Anabaena sp.1 and Anabaena sp.2 require high nitrogen concentrations to reach the highest values of metabolites, including pigments and biomass. Growth and production followed the order: Anabaena sp.1 > Anabaena sp.2 > Nostoc LAUN0015 > Nostoc UAM206. These results showed that nitrogen concentration between 0 and 17 mM NaNO3 modulate the production of biomolecules in both strains of Nostoc and Anabaena.
REFERENCES
Acien F, Fernandez J, Molina-Grima E. Economics of microalgae biomass production. In: Pandey A, Lee D, Chisti Y, Soccol C, editors. Biofuels from alga. Oxford, UK: Elsevier; 2014. p. 313-325. [ Links ]
Arias Peñaranda M, Martínez Rondán A, Cañizares Villanueva R. Producción de biodiesel a partir de microalgas: Parámetros del cultivo que afectan la producción de lípidos. Acta biol Colomb. 2013;18(1):43-68. [ Links ]
Baier K, Nicklisch S, Grundner C, Reinecke J, Lockau W. Expression of two nblA-homologous genes is required for phycobilisome degradation in nitrogen-starved Synechocystis sp. PCC6803. FEMS Microbiol Lett. 2001;195(1):35-39. Doi:10.1111/j.1574-6968.2001.tb10494.x. [ Links ]
Billi D, Grilli M. Effects of nitrogen limitation and starvation on Chroococcidiopsis sp. (Chroococcales). New Phytol. 1996;133(4):563-571. Doi:10.1111/j.1469-8137.1996.tb01925.x. [ Links ]
Canto de Loura I, Dubacq J, Thomas J. The effect of nitrogen deficiency on pigments and lipids of cyanobacteria. Plant Physiol. 1987;83(4):838-843. Doi: 10.1104/pp.83.4.838. [ Links ]
Colla L, Oliveira C, Reichert C, Vieira Costa J. Production of biomass and nutraceutical compounds by Spirulina platensis under different temperature and nitrogen regimes. Biores Technol. 2007;98(7):1489-1493. Doi:10.1016/j.biortech.2005.09.030. [ Links ]
Cunningham S, Joshi L. Algal Biotechnology: An Emerging Resource with Diverse Application and Potential. In: Kole C, Michler C, Abbott A, Hall T, editors. Transgenic Crop Plants, Vol. 1: Principles and Development. Berlin and Heidelberg, Germany: Verlag-Springer; 2010. p. 343-357. Doi:10.1007/978-3-642-04812-8_9. [ Links ]
De Philippis R, Margheri M, Materassi R, Vincenzini M. Potential of unicellular cyanobacteria from saline environments as exopolysaccharide producers. Appl Environ Microbiol. 1998;64(3):1130-1132. [ Links ]
De Philippis R, Vincenzini M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol Rev. 1998;22(3):151-175. Doi: 10.1111/j.1574-6976.1998.tb00365.x. [ Links ]
Foy R. The phycocyanin to chlorophyll a ratio and cell components as indicators of nutrient limitation in two planktonic cyanobacteria subjected to low-light exposure. J Plank Res. 1993;15(11):1263-1276. Doi: 10.1093/plankt/15.11.1263 [ Links ]
Fuenmayor G, Jonte L, Rosales-Loaiza N, Morales E. Efecto de la salinidad y la concentración de nutrientes sobre el crecimiento y composición bioquímica de la cianobacteria autóctona Oscillatoria sp. MOF-06. Ciencia. 2009;17(1):50-57. [ Links ]
Gantar M, Svirčev Z. Microalgae and cyanobacteria: Food for thought. J Phycol. 2008;44(2):260-268. Doi:10.1111/j.1529-8817.2008.00469.x. [ Links ]
Guschina I, Harwood J. Algal lipids and effect of the environment on their biochemistry. In: Arts M, Brett M, Kainz K, editors. Lipids in aquatic ecosystems. Dordrecht, The Netherlands: Springer Academics; 2009. p. 1-24. Doi:10.1007/978-0-387-89366-2_1. [ Links ]
Harun R, Singh M, Forde G, Danquah M. Bioprocess engineering of microalgae to produce a variety of consumer products. Renew Sust Energ Rev. 2010;14(3):1037-1047. Doi:10.1016/j.rser.2009.11.004. [ Links ]
Herbert D, Phipps P, Strange P. Chemical analysis of microbial cells. In: Norris J, Ribbons D, editors. Methods in microbiology Vol 5B. London, UK: Academic Press; 1971. p. 209-344. [ Links ]
Hu Q, Westerhoff P, Vermaas W. Removal of nitrate from groundwater by cyanobacteria: Quantitative assessment of factors influencing nitrate uptake. Appl Environ Microbiol. 2000;66(1):133-139. Doi:10.1128/AEM.66.1.133-139.2000. [ Links ]
Hu Q. Environmental effects on cell composition. In: Richmond A, editor. Handbook of microalgal culture, biotechnology and applied phycology. Oxford, UK: Blackwell Publishers; 2004. p. 83-94. Doi:10.1002/9780470995280. [ Links ]
Kochert G. Carbohydrate determination by the phenol-sulfuric acid method. In: Hellebust J., Craigie J, editors. Handbook of phycological methods. Physiological and biochemical methods. Cambridge, UK: Cambridge University Press; 1978. p. 95-97. [ Links ]
Konopka A, Schnur M. Biochemical composition and photosynthetic carbon metabolism of nutrient limited cultures of Merismopedia tenuissima (Cyanophyceae). J Phycol. 1981;17(2):118-122. Doi:10.1111/j.1529-8817.1981.tb00829.x. [ Links ]
Jindal N, Singh D, Khattar J. Kinetics and physico-chemical characterization of exopolysaccharides produced by the cyanobacterium Oscillatoria formosa. World J Microbiol Biotechnol. 2011;27(9):2139-2146. Doi:10.1007/s11274-011-0678-6. [ Links ]
Jonte L, Rosales N, Briceño B, Morales E. La salinidad y la irradiancia modulan el crecimiento de la cianobacteria Synechocystis minuscula en cultivos discontinuos. Multiciencias. 2003;3(1):1-13. [ Links ]
Lanfer-Marquez U, Barros R, Sinnecker P. Antioxidant activity of chlorophylls and their derivatives. Food Res Int. 2005;38(8-9):885-891. Doi: 10.1016/j.foodres.2005.02.012. [ Links ]
Leal M, Ortiz N, Mora R, Morales E. Caracterización de la cianobacteria Pseudanabaena galeata en condiciones de laboratorio. Bol Centro Invest Biol. 2001;35:223-241. [ Links ]
Leonardos N, Geider R. Responses of elemental and biochemical composition of Chaetoceros muelleri to growth under varying light and nitrate: phosphate supply ratios and their influence on critical N: P. Limnol Oceanogr. 2004;49(6):2105-2114. Doi: 10.4319/lo.2004.49.6.2105. [ Links ]
Lewitus A, Caron D. Relative effects of nitrogen and phosphorus depletion and light intensity on the pigmentation, chemical composition and volume of Pyrenomonas salina (Cryptophyceae). Mar Ecol Progr Ser. 1990;61:171-181. [ Links ]
Loreto C, Rosales N, Bermúdez J, Morales E. Producción de pigmentos y proteínas de la cianobacteria Anabaena PCC 7120 en relación a la concentración de nitrógeno e irradiancia. Gayana Bot. 2003;60(2):83-90. [ Links ]
Marker A, Jinks S. The spectrophotometric analysis of chlorophyll a and pheopigments in acetone, ethanol and methanol. In: Rai H, Marker A, editors. The measurement of photosynthetic pigments in freshwaters and standardization of methods. Arch Hydrobiol Beih Ergebn Limnol. 1982;16:3-17. [ Links ]
Moreno J, Vargas M, Olivares H, Rivas J, Guerrero M. Exopolysaccharide production by the cyanobacterium Anabaena sp. ATCC 33047 in batch and continuous culture. J Biotechnol. 1998;60(3):175-182. Doi: 10.1016/S0168-1656(98)00003-0. [ Links ]
Otero A, Vincenzini M. Extracellular polysaccharide synthesis by Nostoc strains as affected by N source and light intensity. J Biotechnol. 2003;102(2):143-152. Doi:10.1016/S0168-1656(03)00022-1. [ Links ]
Otero A, Vicenzini M. Nostoc (Cyanophyceae) goes nude: extracellular polysaccharides serve as a sink for reducing power under unbalanced C/N metabolism. J Phycol. 2004;40(1):74-81. Doi:10.1046/j.1529-8817.2004.03067.x. [ Links ]
Pattnaik U, Singh P. Effect of nitrate nitrogen on the growth, heterocyst differentiation and nitrogen fixation in rice field blue-green alga Gleotrichia sp. Arch Hydrobiol. 1978;51:318-327. [ Links ]
Rasmussen R, Morrissey M. Marine biotechnology for production of food ingredients. Adv Food Nutr Res. 2007;52:237-292. Doi:10.1016/S1043-4526(06)52005-4. [ Links ]
Rippka R, Deruelles J, Waterbury J, Herdman M, Stanier R. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J Gen Microbiol. 1979;111:1-61. [ Links ]
Rosales N, Jonte L, Morales E. Crecimiento y composición bioquímica de Synechococcus sp. modulados por el nitrato de sodio. Bol Centro Invest Biol. 2006;40(2):120-132. [ Links ]
Rosales-Loaiza N, Avendaño D, Otero A, Morales E. Crecimiento, producción de pigmentos y proteínas de la microalga Dunaliella viridis (Chlorophyta) en cultivos semicontinuos. Bol Centro Invest Biol. 2007;42(3):323-334. [ Links ]
Rosales Loaiza N, Morales E. Synechococcus sp. En función de parámetros y sistemas de cultivo. Cultivo de una cianobacteria de ambientes tropicales y la verificación de su potencial biotecnológico. Barcelona, Spain: Editorial Académica Española; 2013. 109 p. [ Links ]
Roux J. Production of polysaccharide slime by microbial mats in the hypersaline environment of a Western Australian solar saltfield. Int J Salt Lakes Res. 1996;5(2):103-130. [ Links ]
Saha S, Uma L, Subramanian G. Nitrogen stress induced changes in the marine cyanobacterium Oscillatoria willei BDU 130511. FEMS Microbiol Ecol. 2003;45(3):263-272. Doi:10.1016/S0168-6496(03)00162-4. [ Links ]
Sharma N, Prakash Tiwari S, Tripathi K, Rai A. Sustainability and cyanobacteria (blue-green algae): facts and challenges. J Appl Phycol. 2011;23(6):1059-1081. Doi:10.1007/s10811-010-9626-3. [ Links ]
Serpa R, Calderón A. Efecto de diferentes fuentes de nitrógeno en el contenido de carotenoides y clorofila de cuatro cepas peruanas de Dunaliella salina TEOD. Ecol Apl. 2006;5(1,2):93-99. [ Links ]
Simeunović J, Bešlin K, Svirčev Z, Kovač D, Babić O. Impact of nitrogen and drought on phycobiliprotein content in terrestrial cyanobacterial strains. J Appl Phycol. 2013;25(2):597-607. Doi:10.1007/s10811-012-9894-1. [ Links ]
Singh S, Das S. Screening, production, optimization and characterization of cyanobacterial polysaccharide. World J Microbiol Biotechnol. 2011;27(9):1971-1980. [ Links ]
Stephens E, Wagner L, Ross I, Hankamer B. Microalgal production systems: Global impact of industry scale-up. In: Posten C, Walter C, editors. Microalgal biotechnology: Integration and economy. Boston, USA: De Gruyter; 2012. p. 267-306. [ Links ]
Strickland J, Parsons T. A practical handbook of seawater analysis. Fish Res Board Can Bull. 1972;167:310. [ Links ]
Tsygankov A. Nitrogen-fixing cyanobacteria: a review. Appl Biochem Microbiol. 2007;43(3):50-259. Doi:10.1134/S0003683807030040 [ Links ]
Utting S. Influence of nitrogen availability on the biochemical composition of three unicellular marine algae of commercial importance. Aquacul Engin. 1985;4(3):175-190. Doi: 10.1016/0144-8609(85)90012-3. [ Links ]
Vincent W. Cyanobacteria. In: Likens G, editor. Encyclopedia of inland waters. San Diego, USA: Elsevier; 2009. p. 226-232. [ Links ]
Vonshak A, Torzillo G. Environmental stress physiology. In: Richmond A, editor. Handbook of microalgal culture: Biotechnology and applied phycology. Oxford, UK: Blackwell Publishing; 2004. p. 57-82. [ Links ]
Whitton B, Potts M. Introduction to the cyanobacteria. In: Whitton B, Potts M, editors. The ecology of cyanobacteria II: Their diversity in time and space. Dordrecht, The Netherlands: Kluwer Academic Publishers; 2012. p. 1-14. [ Links ]

