










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.21 no.2 Bogotá May./Aug. 2016
https://doi.org/10.15446/abc.v21n2.48675
Doi: http://dx.doi.org/10.15446/abc.v21n2.48675
INTERACTIONS BETWEEN CALVES OF AMAZONIAN MANATEES IN PERU: A STUDY CASE
Interacciones entre crías de manatí amazónico en Perú: un caso de estudio
Alexander Roldán ARÉVALO-SANDI1, Delma Nataly CASTELBLANCO-MARTÍNEZ2.
1 INPA - PPG-Ecologia, Instituto Nacional de Pesquisas da Amazônia. Manaus, Brazil.
2 CONACyT Research Fellow. Consejo Nacional de Ciencia y Tecnología, Universidad de Quintana Roo. Chetumal, Quintana Roo, Mexico.
For correspondence. alexander.arevalo85@gmail.com
Received: 28th January 2015, Returned for revision: 7th July 2015, Accepted: 4th November 2015. Associate Editor: Enrique Arbeláez-Cortés.
Citation / Citar este artículo como: Arévalo-Sandi AR, Castelblanco-Martínez DN. Interactions between calves of Amazonian manatees in Peru: a study case. Acta biol. Colomb. 2016;21(2):355-364. doi: http://dx.doi.org/10.15446/abc.v21n2.48675
ABSTRACT
Trichechus inunguis is an endemic species of the Amazon, which inhabits mainly in lakes and calm rivers. The objective of this study case was to describe the social behaviour of two female-orphaned calves, of T. inunguis in captivity. They were kept in the same pool at the facilities of the Amazon Rescue Center (ARC, Iquitos, Peruvian Amazon). Between February and October 2011, the individuals were observed during day and night times, completing 352 hours of observation. Through ad libitum observation of the individuals, we developed a catalogue of social behaviors that includes descriptions of 93 behaviours, classified in eight behavioural categories. The frequency of behaviours was assessed by instantaneous sampling (for states) and continuous recording (for events). Manatees displayed mainly social behaviours, and the most frequent interaction was the simultaneous starting of the same behaviour by both individuals. Most of social behaviours occurred during the day, but 'group resting' was recorded mostly at night. 'Synchronized breathing' and 'group feeding' did not have temporary variation between days or between daytimes. Regarding the use of the space, the majority of interactions occurred in shaded places. This study revealed strong interactions between confined calves, suggesting that social activity may play an important role in their learning process.
Keywords: endangered species, ethology, social behaviour, Trichechus inunguis.
RESUMEN
Trichechus inunguis es una especie endémica del Amazonas que vive principalmente en lagos y ríos de aguas tranquilas. El objetivo de este estudio de caso fue describir el comportamiento social de dos crías hembras huérfanas de T. inunguis en cautiverio, mantenidas en el mismo estanque en el Centro de Rescate Amazónico (CREA, Iquitos, Amazonía peruana). Entre febrero y octubre de 2011, las crías fueron observadas durante el día y la noche, para un total de 352 horas de observación. Mediante observación ab libitum de los individuos, desarrollamos un catálogo de comportamientos social que incluye descripciones de 93 comportamientos clasificados en ocho categorias comportamentales. La frecuencia de comportamientos fue evaluada por muestreo instantáneo (para estados) y registro continúo (para eventos). Los manatíes exhibieron principalmente comportamientos sociales, y la interacción más frecuente fue el inicio simultáneo del mismo comportamiento por ambos individuos. La mayoría de los comportamientos sociales ocurrieron durante el día, pero 'descanso grupal' fue registrado mayormente en la noche. 'Respiración sincronizada' y 'alimentación grupal' no tuvieron variación diaria entre días o entre horas del día. En lo que respecta al uso de espacio, la mayoría de interacciones ocurrió en lugares sombreados. Este estudio reveló fuertes interacciones entre crías en cautiverio, sugiriendo que la actividad social cumple un rol importante en los procesos de aprendizaje de los manatíes.
Palabras clave: comportamiento social, especie amenazada, etología, Trichechus inunguis.
INTRODUCTION
The Amazonian manatee Trichechus inunguis (Natterer, 1883) is the smallest manatee and exclusively inhabits freshwater environments. The species is endemic to the Amazon region, living along the Amazonian Basin mainly in calm rivers and lakes (Domning et al., 1981; Montenegro-Paredes, 1994; Rosas, 1994). The Amazonian manatee is a non-ruminant herbivore that feeds on a wide variety of aquatic and semiaquatic plants (Guterres et al., 2008). The species plays an important role in the ecosystem, returning more than half of the ingested material to the environment as feces and urine, and is therefore essential to primary production (Best, 1984; Timm et al., 1986). The commercial exploitation (Domning, 1982), along with the slow rate of reproduction (Rodrigues et al., 2008), have seriously reduced the Amazonian manatee population (IUCN, 2008). The species is currently considered Vulnerable, according to the Red List of Threatened Species of the International Union for Conservation of Nature (IUCN, 2015).
Amazonian manatees are difficult to observe in natural environments, due to their dark colour, long underwater times and solitary and cryptic behaviour. Also, the species' habitat is extensive and complex: manatees can easily camouflage in the turbid waters of the Amazon Basin rivers and abundant vegetation associated to the water bodies. Therefore, encounters with Amazon manatees in wild are very rare, and systematic observation of its natural behaviour is almost impossible. Keeping individuals in captivity with rehabilitation purposes provides a unique opportunity to develop ethological studies. Studies of manatee behaviour in captivity have provided useful information on training (Colbert et al., 2001; Lima et al., 2005), communication and auditory capabilities (Bullock et al., 1980; Gerstein et al., 1999; Mann et al., 2005; Colbert et al., 2008), visual acuity (Bauer et al., 2003; Maple and Segura 2015), tactile discrimination (Marshall et al., 1998) and learning behaviour (Gerstein, 1994), among others (Bauer et al., 2010). The results of these studies are useful not only to evaluate the conditions of a particular individual, but also to open new insights on the manatee behaviour knowledge.
Illegal trade in wild animals is a global conservation and animal welfare concern (Daut et al., 2004). Rehabilitation plans are often created to protect animal populations at risk of extinction, providing animals with a second chance to survive in the wild and contributing to the population's gene pool. Therefore, rehabilitation and reintroduction has a greater value when it contributes to species conservation and population growth (Adimey et al., 2012). An adequate study of manatee behaviour might allow us to recognize stress behaviours and other abnormal conditions that would serve later to validate a possible reintroduction to the wild (Castelblanco-Martínez and Zerda-Ordóñez, 2000; Manire et al., 2003; Normande et al., 2015). Also, by observing the interaction behaviours between conspecifics and with humans, researchers can infer if an animal will be able to survive in the wild, especially in areas where hunting pressure is a concern. Therefore the characterization of behavioural patterns could serve as a baseline for monitoring during rehabilitation for reintroduction process (Gomes et al., 2008).
Solitary or transient social systems are predominant among manatees (Marmontel et al., 1992), and therefore Amazonian manatees are considered semi-social or moderately social animals (Hartman, 1979; Reynolds III, 1981). It is widely accepted that the only readily identifiable social unit in sirenians is the cow-calf pair, with bonds persisting for 1-2 yrs (Anderson, 2002). 'Juvenile' or 'sub adult' manatees are weaned animals that are not fully grown (usually smaller than an adult), or sexually mature (Kunz et al., 1996). 'Adults' are biologically capable of producing offspring, in other words males that present spermatogenesis (Hernández et al., 1995) and females ovulating (Marmontel, 1995). In this study 'group behaviour' is understood as any interaction occurring between two or more individuals.
By observing and analysing two calves in captivity, we tested the following hypotheses: (i) Social behaviours are not important, in comparison with non-social behaviours, (ii) there is not a significant difference between categories of social behaviours, (iii) there is not a rhythm of social activities during the times of the day, and (iv) there is not a preference in the use of the space to develop social activities.
Therefore, the aim of this research was to identify and describe captive/orphan manatee social behaviour in order to gather baseline information that can be useful for decision making during manatee calves' rehabilitation. The specific objectives were: (1) To develop a catalog of social behaviour of T. inunguis individuals in order to obtain a descriptive baseline for orphan/captive manatee behaviour that can be used for further studies, (2) to infer the relevance of social activities for calves in rehabilitation in order to give advises for manatee management in captivity, and (3) to determine the temporal and spatial variation of social activities in order to define preferences of space and time to display social behaviours.
MATERIAL AND METHODS
Subjects
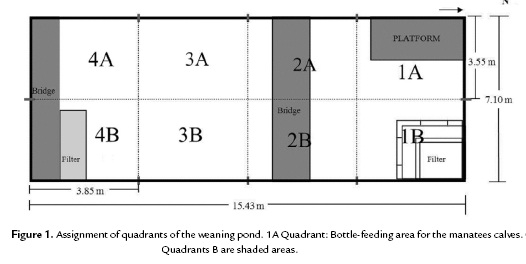
We studied the social behaviour of two orphan females of Amazonian manatees rescued at the Peruvian Amazon: (i) Nauta was 18-months-old and was rescued near Nauta town, Loreto Province on 14 March 2010, and (ii) Yuri was one-year-old and was rescued in Alto Amazonas Province on 26 November 2010. The rescued calves were transferred to individual quarantine ponds of the ARC facilities with the goal of rehabilitate them for further release. The dimensions of the quarantine ponds were 2.80 x 1.95 x 0.75 m, with an approximate capacity of 4.095 L. Once rehabilitated and habituated to bottle feeding, they were moved together to the weaning pond with dimensions of 15.43 x 7.10 x 0.75 m. with an approximate capacity of 82.16475 L (Fig. 1). The behavioural study was developed in the weaning pool. The manatees were bottle fed during the study; the daily diet consisted of milk solution mixed with antiflatulent ingredients and vegetable oil. As part of the weaning process, small quantities (30 kg per day), of water lettuce (Pistia stratiotes) were provided to familiarize the animals with vegetable food. The calves' growth was measured throughout the study.
Recognition phase
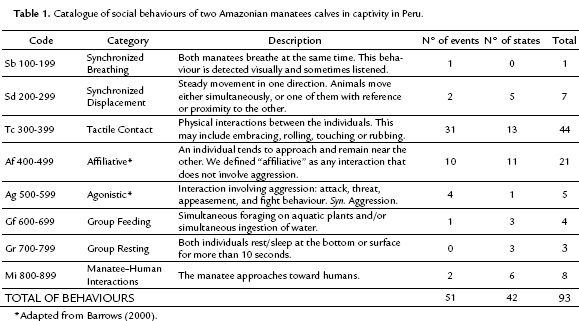
The research was conducted from February to August 2011, with the first phase aiming to obtain a catalog of social behaviours. The selected methods were continuous recording (Martin and Bateson, 1993) with focal and ad libitum sampling (no sampling restrictions; Lehner, 1996), which have been proved practicable for the collection of detailed behavioural data on manatees (Gomes et al., 2008). The animals were observed for sampling periods of 30 minutes, covering day and night times, during which all instances of social behaviour - interaction between the manatees or between a manatee and a human - were recorded (Martin and Bateson, 1993). The recording method selected allowed us to collect true frequencies and durations, and the times at which behaviour patterns stopped and started. Information about the behaviours' duration was also noted, in order to discriminate among 'states' (long-duration behaviours, or a duration meaningful behaviour, > 5-s) and 'events' (short-duration behaviours approaching to an instantaneous occurrence, momentary behaviours, or change of states, < 5-s) (Lehner, 1996).
The cumulative number of observed behaviours was plotted to determine the moment in which the majority of social behaviours was recorded. This analysis is based on the principle that as the number of observations increases, the number of new behaviours decreases, so that an asymptotic curve results (Lehner, 1996). When the asymptote in the curve was reached, it was assumed that most of the behaviours were observed, and preliminary observations were suspended. Moreover, it was obtained the sample coverage (Ɵ) by calculating the probability next behavioural act will be a new type. If Ɵ approaches 1, the probability of observing a new behavioural act is low (Lehner, 1996). Then, the catalogue of behaviours was organized discriminating between 'events' and 'states'. The behavioural categories were based on previous behavioural observations of manatees indicated in: Castelblanco-Martínez (2000), Charry (2002), Holguin-Medina (2008) and Mercadillo-Elguero et al. (2014); and were coded accordingly.
Establishment of social behaviour budgets
The purpose of the second phase was to determine the frequency of social activities (behaviour time budgets; Caraco, 1979) and to establish their temporal and spatial variation. Observations were taken during 333 hours (103.5 hours between 6:00 h and 12:00 h, 77 hours between 12:00 h and 18:00 h; 78 hours between 18:00 h and 24:00 h, 74 hours between 24:00 h and 6:00 h) between 7 April 2011 and 21 July 2011. The behaviour was recorded from a position that provided a full view of the entire enclosure, but precluded any interaction between the subjects and the observer (Gomes et al., 2008). Fluorescent lamps located on the study pond were used for night-time sampling, to easily locate the dyad.
Both types of social behaviours ('events' and 'states') were recorded simultaneously during 30-minutes sampling periods. However, 'states' were registered by instantaneous sampling and 'events' by continuous recording. During the instantaneous method, the 30-minutes sampling periods were subdivided into one minute intervals as determined by Castelblanco-Martínez (2000), so a total of 30 points/sampling period were defined. In each point, the observer scored the dyad 'state'. We also attempt to assess the directionality of social interactions, where ties reflect the initiator and recipient of the relationship (Wey et al., 2008). The initiator of the behaviour was recorded as follows: Yuri initiating an interaction with Nauta (Y→Na), Nauta initiating an interaction with Yuri (Na→Y), both individuals initiating an interaction simultaneously (Y↔Na), or one of the manatees initiating an interaction with humans (Na→H or Y→H). Behaviours were considered to be synchronized when displayed by both individuals simultaneously, and could be initiated by any one of them or both. 'Events' were recorded by focal-animal sampling and continuous recording; the researcher recorded a complete account of all behaviour units of interests (bouts), obtaining frequencies of each behaviour.
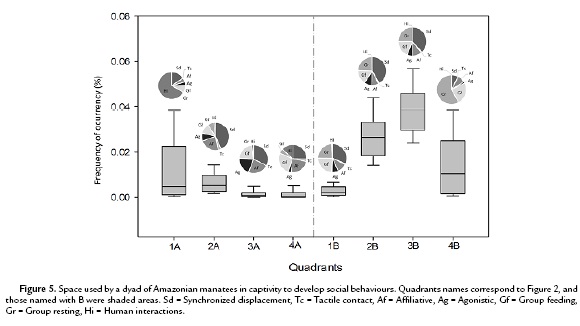
With the goal of assessing the horizontal use of space, the pond was divided into eight equal imaginary quadrants (Fig. 1). The position of the dyad was registered every minute, i.e., simultaneously with the 'states' recording.
Data analysis
'States' frequency was represented in terms of percentages. For each sample period, budgets of the 'states' of social interaction were compiled by summing the total of points (i.e. each sample record) when a 'state' was recorded and dividing this sum by the total of observation points (i.e., 30). Therefore, the 'state' frequency was determined by the formula fs = (ns/30)*100; where s = the state and n = number of points when the states were recorded. Frequency rates of 'events' were calculated for each sampling period by dividing the total number of bouts by the number of minutes of observation (fe= bouts/30). Budgets were also assessed separately for day and night samples to investigate the influence of the sunlight on social behaviour. The Friedman test (non-parametric alternative to the one-way ANOVA with repeated measures) was used to detect the difference between day (06:00 to 18:00) and night (18:00 to 06:00). Statistical analyses were done using PAST 3.x (Hammer et al., 2001). Finally, we established social activity budgets for each quadrant in order to examine any potential space preference in which to perform social interactions. For this, the relative frequency of social interactions per time unit was measured for each quadrant. The normality assumption was verified with the Shapiro-Wilk test. Overall comparisons between groups of social behaviour were made with Kruskal-Wallis one-way analysis of variance on ranks; followed by a Tukey pair-wise multiple comparison procedure. The paired t-test was applied to compare two groups of data (for example, between shaded and non-shaded quadrants). When the data failed to show normality, a Wilcoxon Signed Rank test was used. In this point, the statistical analyses were done using SigmaPlot (Kornbrot, 2000).
RESULTS
We completed 352 hours of observation. The asymptotic cumulative curve of behaviours was obtained after 104 samples (a sample consisting of 30-min observation period), with a total of 93 social behaviours described (52 events and 41 states, see Arevalo-Sandi 2012). The value obtained of the coverage sample (Ө) was 0.90. The majority (74 %) of state behaviours observed corresponded to social interactions. The catalog of social behaviours was divided into the following categories (codes in parenthesis): Synchronized breathing (100-199), synchronized displacement (200-299), tactile contact (300-399), affiliative (close-proximity behaviour that includes touching, grooming, and nuzzling; 400-499), agonistic (any social behaviour related to fighting, such as aggressive or submissive behaviours; 500-599), group feeding (600-699), group resting (700-799), and human interactions (800-899). See Table 1 for a detailed description of each category. The Kruskal-Wallis analysis of variance found a significant difference between social categories (H=230.207, df=2, p<0.001). The most common category was synchronized displacement (32 %), followed by group resting (25 %) and group feeding (18 %) (Fig. 2).
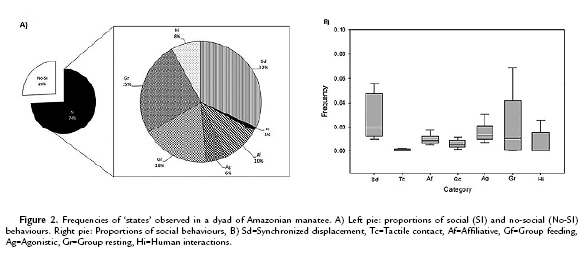
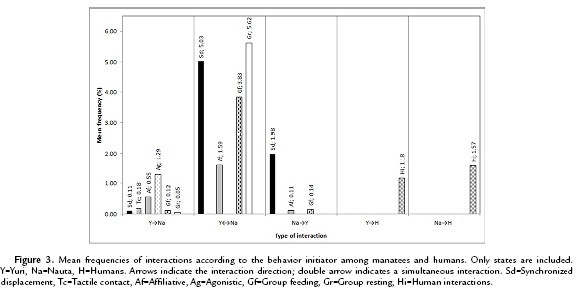
In most of the cases (68.7 %), the interaction was initiated simultaneously (Y ↔ Na) (Fig. 3). In this type of interaction, categories group resting and synchronized displacement showed the highest frequency. Yuri and Nauta played the initiator role in similar proportion (9.82 % and 9.68 % respectively). However, Nauta often initiated displacement conducts (synchronized displacement), while Yuri initiated mostly agonistic behaviours. Nauta was never observed starting agonistic interactions. Interactions with humans took place during bottle feeding, and also during the presence of visitors. In all the cases, both manatees started interactions with humans with similar frequency (Yuri = 5.03 %, Nauta = 6.72 %).
Social interactions were observed during both day and night, but were significantly more frequent during the night (Wilcoxon test: Z=4.286, p<0.001). Also, the daily rhythm varied depending on the category (Fig. 4). The statistical analysis showed significant differences in behavioural frequency between day (6:00 to 18:00) and night (18:00 to 6:00) for the following categories: Synchronized displacement (Friedman test statistics= 16.66, df=1, p<0.0001), tactic contact (Friedman test statistics=24, df=1, p<0.0001), affiliative (Friedman test statistics=24, df=1, p<0.0001), agonistic (Friedman test statistics =15.04, df=1, p<0.0001), group resting (Friedman test statistics=0.66, df=1, p=0.004) and human interactions (Friedman test statistics=0.66, df=1, p=0.00035). No significant differences were found for synchronized breathing and group feeding.
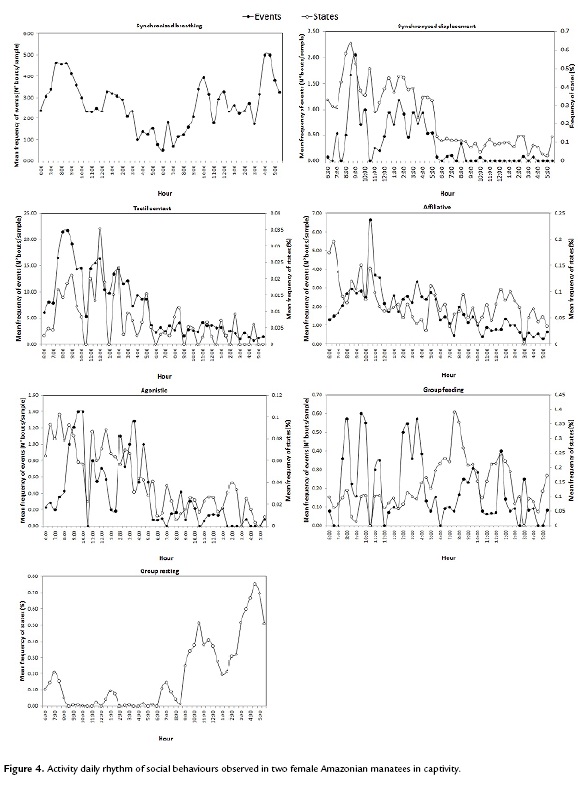
The use of space-frequency (%) differed significantly between quadrants (Kruskal Wallis; H=42.6, df=6, p<0.001) (Fig. 5). In general, shaded quadrants particularly 2B, 3B and 4B, were preferred while displaying social behaviours (paired t-test: t=4.45, df=47, p >0.001).
DISCUSSION
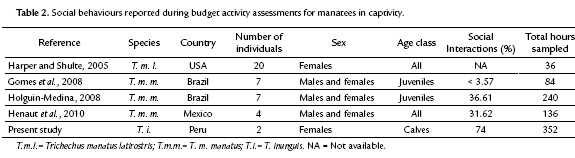
Both individuals showed a normal growing rate compared with other Amazonian calves in captivity (Sousa Lima, 1999). During the time of the study, Nauta grew from 50 to 62 kg (0.6 kg/week), while Yuri increased her weight from 28 kg to 40 (0.6 kg/week). As about the behaviours, the obtained sample coverage value (Ө=0.90) was slightly lower than those reported for Trichehus manatus (Ө=0.91; Mercadillo-Elguero et al., 2014, and for T. inunguis (Ө=0.94 and Ө=0.96; Castelblanco-Martínez, 2000 and Charry, 2002 respectively). However, we consider that the majority (90 %) of the probable social behaviours were included in the catalog. The total number of observed social behaviours was higher (74 %) than that obtained in other studies such as: Harper and Schulte (2005), Gomes et al. (2008), Holguin-Medina (2008) and Hénaut et al. (2010); probably due to the greater sampling effort conducted in the present study (Table 2).
The manatees observed in the mentioned studies (4 - 20 individuals) were calves, juveniles and adults, while manatees observed in the present study were just two orphaned calves (suckling, bottle-fed individuals). Due to the fact that these individuals were still in the early stages of development, it is to be expected that social interaction will be actively searched for more often (Bekoff, 1972). Hénaut et al. (2010) observed that a calf with his mother presented the highest number of interactions in a pool with several adults. Activities that may reinforce the female-calf bond are nuzzling, mouthing and synchronous breathing (Reynolds III, 1981). Orphan manatee calves have been observed to intently look for physical proximity to other individuals, regardless of age or sex (Castelblanco-Martínez, per. obs.). In this study, most of the interactions were initiated by the youngest calf (Yuri), while Nauta was usually indifferent to Yuri. This suggests that younger manatees look more intensely for attention from older individuals. Like other large mammals, sirenian calves gain nutritional independence gradually as they shift from a milk diet to solid food (Langer, 2003). The weaning is divided in two phases: 1) when only milk is taken and 2) a mixed period: in which the milk intake is combined with the consumption of solid vegetation. The longer the mixed-feeding period, the more time is available for ontogenic developments, for instance, learning how to obtain food, avoiding predators, and establishing immune competence (Langer, 2003). That means that even when the calf is relatively independent from the mother in terms of nutritional intake, a long calving period is needed to reinforce behaviours that could be decisive for survival. Previous studies suggested that during inter-individual contact, the manatee calves learn and develop social skills and recognition abilities by being exposed to conspecifics and thus build a type of social map used when manatees come together in groups or when encountering isolated individuals in the wild (Hénaut et al., 2010). Our findings reinforce the recommendation that, during rehabilitation processes, maintaining several manatee calves in the same pond might facilitate the creation of inter-individual bonds, which could be important for survival in wild if manatees are released together.
The synchronous surfacing (breathing) has been observed in wild Florida manatees (Hartman, 1979) and dugongs (Anderson, 1995). It is possible that it is an extension of behaviour learned at birth when a mother synchronizes her movements with those of her offspring. It has been suggested that breathing in synchronous fashion may help reduce an individual's chance of being captured by a predator (Hastie et al., 2003). Synchronous surfacing ensures closer side-by-side contact between animals, decreasing the odds of accidental separation, a constant hazard in turbid waters, especially for a calf (Hartman, 1979). Synchronicity in cow-calf pairs is well known (Hartman, 1979), and since both individuals lost their mothers at an early age and are of similar age, our results suggest that these two individuals may have been considering each other "substitute mothers". Bonde (2009) presented some intriguing genetic evidence of wild Florida manatee suckling adopted calves. If adoption in manatees is widespread, orphan calves prone to seek the company of an older conspecific must facilitate such adopting mechanism. Both studied manatees also searched intensively for human contact, as a response of feeding conditioning. This behaviour may be problematic in the future, but necessary during the bottle feeding period. For future plans of release, human contact must be limited to guarantee an adequate adaptation to the wild.
Even though social activities were recorded at any time, the manatees studied showed preference to rest simultaneously during the night. A previous study showed certain daily rhythm in the activities of a captive calf Amazonian manatee; with more display of breathing and displacement behaviours during daytime (Castelblanco-Martínez, 2000). However, it is not clear if the rhythm of activity is a consequence of the artificial bottle feeding schedule which occurs only during the day. The studied manatees showed predilection for indoor areas, maybe because these areas were perceived as safer. This behaviour would be useful in the natural habitat for a possible camouflage and thus avoid humans and possible predators, as mentioned by Charry (2002).
CONCLUSIONS
We present a study-case of captive/orphan Amazonian manatee social behaviour. Our study revealed strong interactions between the calves, suggesting that social activity may play an important role in the learning process of the individuals. However, caution is necessary in extrapolating results from our observations to natural behaviour. There are no studies on social behaviour of wild Amazonian manatees, and our findings should be only interpreted into the frame of particular circumstances, such as the confinement during a rehabilitation process. Despite of this, this work may represent the beginning of the collection of long-term baseline information about Amazonian manatees' behaviour in captivity. This might help implement adequate management in captivity in Peru and, consequently, increase the odds of a successful rehabilitation process.
ACKNOWLEDGEMENTS
This study was part of the bachelor thesis of ARA-S from Universidad Nacional de la Amazonía Peruana. This work was possible thanks to the Amazon Rescue Center, the Instituto de Investigaciones de la Amazonia Peruana and the Dirección Regional de la Producción Loreto. We thank Daryl Richardson and Luis Sigler of The Dallas World Aquarium-Zoo. We also acknowledge ARC's visitors, volunteers and interns, especially Roxana Pérez, for their help in data collection. Special thanks to Margherita Bandini for helping with the English correction, and to Fabrício B. Baccaro for the latest help in statistical analysis. Roberto Pezo Díaz and Javier Velásquez Varela for provided guidance and advice. We also would like to thank Artie Wong of Save the Manatee Club for providing bibliographic resources.
REFERENCES
Adimey NM, Mignucci-Giannoni A, Auil Gomez N, Da Silva VMF, Alvite C, Morales-Vela B, et al. Manatee rescue, rehabilitation, and release efforts as a tool for species conservation. In: Hines E, Reynolds J, Aragones L, Mignucci-Giannoni AA, Marmontel M, editors. Sirenian conservation: Issues and strategies in developing countries. Florida: University Press of Florida; 2012. p. 205-217. [ Links ]
Anderson PK. Scarring and photoidentification of dugongs (Dugong dugon) in Shark Bay, Western Australia. Aquat Mamm. 1995;21(3):205-211. [ Links ]
Anderson PK. Habitat, niche, and evolution of Sirenian mating systems. J Mamm Evol. 2002;9(1):55-98. Doi:10.1023/A:1021383827946. [ Links ]
Arévalo-Sandi AR. Comportamiento social del manatí amazónico (Trichechus inunguis) en cautiverio en un estanque artificial del Centro de Rescate Amazónico (CREA) en el IIAP". Iquitos: Universidad Nacional de la Amazonía Peruana; 2012. 65 p. [ Links ]
Barrows EM. Animal behavior desk reference: a dictionary of animal behavior, ecology, and evolution. Florida: CRC Press LLC; 2000. 936 p. [ Links ]
Bauer GB, Colbert DE, Gaspard JC, Littlefield B, Fellner W. Underwater visual acuity of Florida manatees (Trichechus manatus latirostris). Int J Comp Psychol. 2003;16(2):130-142. [ Links ]
Bauer GB, Colbert DE, Gaspard I, Joseph C. Learning about manatees: a collaborative program between New College of Florida and Mote Marine Laboratory to conduct laboratory research for manatee conservation. Int J Comp Psychol. 2010;23(4):811-825. [ Links ]
Bekoff M. The Development of social interaction, play, and metacommunication in mammals: an ethological perspective. Q Rev Biol. 1972;47:412-434. [ Links ]
Best RC. The Aquatic mammals and reptiles of the Amazon. In Sioli H, editor. The Amazon limnology and landscape: ecology of a mighty tropical river and its basin. Netherlands: Springer; 1984. p. 371-412. [ Links ]
Bonde RK. Population genetics and conservation of the Florida manatee: Past, present, and future. Florida: University of Florida; 2009. 144 p. [ Links ]
Bullock TH, Domning DP, Best RC. Evoked brain potentials demonstrate hearing in a manatee (Trichechus inunguis). J Mammal. 1980;61(1):130-133. Doi:10.2307/1379969. [ Links ]
Caraco T. Time budgeting and group size: a test of theory. Ecology. 1979;60(3):618-627. Doi:10.2307/1936082. [ Links ]
Castelblanco-Martínez DN. Estudio del comportamiento de un juvenil de manatí amazónico, Trichechus inunguis, en Cautiverio. Puerto Nariño-Amazonas, Colombia. Bogotá: Universidad Nacional de Colombia; 2000. 195 p. [ Links ]
Castelblanco-Martínez DN, Zerda-Ordóñez E. Study of the behavior of Amazonian manatee (Trichechus inunguis) in captivity (Puerto Nariño - Amazonas, Colombia). Acta biol Colomb. 2000;5(1):83. [ Links ]
Charry A. Patrones de comportamiento y uso de hábitat del manatí Amazónico Trichechus inunguis en condiciones de semicautiverio. Bogotá: Universidad Nacional de Colombia; 2002. 168 p. [ Links ]
Colbert DE, Fellner W, Bauer GB, Manire C, Rhinehart H. Husbandry and research training of two Florida manatees, Trichechus manatus latirostris. Aquat Mammals. 2001;27(1):16-23. [ Links ]
Colbert DE, Gaspard III JC, Reep R, Mann D, Bauer GB. Four-choice sound localization abilities of two Florida manatees, Trichechus manatus latirostris. J Exp Biol. 2008;212(13):2105-2112. Doi:10.1242/jeb.089227. [ Links ]
Daut EF, Brightsmith DJ, Peterson MJ. Role of non-governmental organizations in combating illegal wildlife-pet trade in Peru. J Nat Conserv. 2015;24:72-82. Doi:10.1016/j.jnc.2014.10.005. [ Links ]
Domning DP. Distribution and status of manatees Trichechus spp. near the mouth of the Amazon River, Brazil. Biol Conserv. 1981;19(2):85-97. Doi:10.1016/0006-3207(81)90044-6. [ Links ]
Domning, DP. Commercial exploitation of manatees Trichechus in Brazil c. 1785-1973. Biol Conserv. 1982;22(2):101-126. Doi:10.1016/0006-3207(82)90009-X. [ Links ]
Gerstein ER. The manatee mind: Discrimination training for sensory perception testing of West Indian manatees (Trichechus manatus). Mar Mammal Public Disp Res. 1994;1(1):10-21. [ Links ]
Gerstein ER, Gerstein L, Forsythe SE, Blue JE. The underwater audiogram of the West Indian manatee (Trichechus manatus). J Acoust Soc Am. 1999;105(6):3575-3583. Doi:10.1121/1.424681. [ Links ]
Gomes FFA, Vergara-Parente JE, Ferrari SF. Behaviour patterns in captive manatees (Trichechus manatus manatus) at Itamaracá Island, Brazil. Aquat Mamm. 2008;34(3):269-276. Doi:10.1578/AM.34.3.2008.269. [ Links ]
Guterres MG, Marmontel M, Ayub DM, Singer RF, Singer RB. Anatomia e morfologia de plantas aquáticas da Amazônia-utilizadas como potencial alimento por peixe-boi amazônico. Belém: IDSM; 2008. 187 p. [ Links ]
Hammer Ø, Harper DAT, Ryan PD. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001;4(1):9. [ Links ]
Harper J, Schulte B. Social interactions in captive female Florida manatees. Zoo Biol. 2005;24(2):135-144. Doi:10.1002/zoo.20044. [ Links ]
Hartman DS. Ecology and behavior of the manatee (Trichechus manatus) in Florida. New York: American Society of Mammalogists Special Publication No. 5; 1979. 153 p. [ Links ]
Hastie GD, Wilson B, Tufft LH, Thompson PM. Bottlenose dolphins increase breathing synchrony in response to boat traffic. Mar Mamm Sci. 2003;19(1):74-084. Doi:10.1111/j.1748-7692.2003.tb01093.x. [ Links ]
Hénaut Y, Becerra S, Machkour-M'Rabet S, Morales-Vela B, Winterton P, Delfour F. Activities and social interactions in captive Antillean manatees in Mexico. Mammalia. 2010;74(2):141-146. Doi:10.1515/MAMM.2010.016. [ Links ]
Hernández P, Reynolds III JE, Marsh H, Marmontel M. Age and seasonality in spermatogenesis of Florida manatees. In: O'Shea TJ, Ackerman B, Percival HF, editors. Population Biology of the Florida Manatee. Washington, DC: National Biological Service Information and Technology Report 1. US Department of the Interior. Washington, Technical Report Series; 1995. p. 84-97. [ Links ]
Holguin-Medina VE. Comportamento do peixe-boi (Trichechus manatus manatus) nos oceanários de Itamaracá: Manejo e condições abióticas. Recife: Universidade Federal Rural de Pernambuco; 2008. 115 p. [ Links ]
Iucnredlist.org. Cambridge: The IUCN Red List of Threatened Species 2008 [updated 2015-3; Cited 2015 sept 27]. Available from: http://www.iucnredlist.org/details/22102/0. [ Links ]
IUCN red list: The IUCN Red List of Threatened Species. Cambridge: IUCN, c2008 [cited 2015 Sept 27]. Available from: http://www.iucnredlist.org/details/22102/0. [ Links ]
Kornbrot D. Statistical software for microcomputers: SigmaPlot 2000 and SigmaStat2. Br J Math Stat Psychol. 2000;53:335-337. [ Links ]
Kunz TH, Wemmer C, Hayssen V. Assessment of sex, age and reproductive conditions of mammals, p. 270-279. In: Wilson DE, Cole FR, Nichols JD, Rudran R, Foster MS, editors. Measuring and monitoring biological diversity, standard methods for mammals. California: Smithsonian Books; 1996. 409 p. [ Links ]
Langer P. Lactation, weaning period, food quality, and digestive tract differentiations in Eutheria. Evolution. 2003;7(5):1196-1215. Doi: 10.1554/0014-3820(2003)057[1196:LWPFQA]2.0.CO;2. [ Links ]
Lehner PN. Handbook of ethological methods. Cambridge: University Press; 1996. 672 p. [ Links ]
Lima DDS, Vergara-Parente JE, Young RJ, Paszkiewicz E. Training of Antillean manatee Trichechus manatus manatus Linnaeus, 1758 as a management technique for individual welfare. Lat Am J Aquat Mammal. 2005;4(1):61-68. Doi:10.5597/lajam00071. [ Links ]
Manire CA, Walsh CJ, Rhinehart HL, Colbert DE, Noyes DR, Luer CA. Alterations in blood and urine parameters in two Florida manatees, Trichechus manatus latirostris, from simulated conditions of release following rehabilitation. Zoo Biol. 2003;22(2):103-120. Doi:10.1002/zoo.10074. [ Links ]
Mann DA, Colbert DE, Gaspard JC, Casper BM, Cook MLH, Reep RL, et al. Temporal resolution of the Florida manatee (Trichechus manatus latirostris) auditory system. J Comp Physiol B. 2005;191(1):903-908. Doi:10.1007/s00359-005-0016-2. [ Links ]
Maple TL, Segura VD. Advancing behavior analysis in zoos and aquariums. Behav Analyst. 2015;38(1)77-91. Doi:10.1007/s40614-014-0018-x. [ Links ]
Marmontel M, Odell DK, Reynolds III JE. Reproductive biology of South American manatees. In Hamlett WC, editor. Reproductive Biology of South American Vertebrates. New York: Springer; 1992. p. 295-312. [ Links ]
Marmontel M. Age and reproduction in female Florida Manatees. In: O'Shea TJ, Ackerman BB, Percival F, editors. Population Biology of the Florida Manatee. Population Biology of the Florida Manatee Washington, DC: National Biological Service Information and Technology Report 1, US Department of the Interior. Washington: Technical Report Series; 1995. p. 98-119. [ Links ]
Marshall CD, Huth GD, Edmonds VM, Halin DL, Reep RL. Prehensile use of perioral bristles during feeding and associated behaviors of the Florida manatee (Trichechus manatus latirostris). Mar Mamm Sci. 1998;14(2):274-289. Doi:10.1111/j.1748-7692.1998.tb00716.x. [ Links ]
Martin P, Bateson P. Measuring behaviour, an introductory guide. United Kingdom: Cambridge University Press; 1993. 200 p. [ Links ]
Mercadillo-Elguero MI, Castelblanco Martínez DN, Padilla Saldivar JA. Behavioral patterns of a manatee in semi-captivity: implications for its adaptation to the wild. J Mar Anim Ecol. 2014;7(2):31-41. [ Links ]
Montenegro-Paredes MI. Distribucion espacial de la vaca marina Trichechus inunguis (Mammalia: Sirenia) en el rio Amazonas, Trapecio Amazonico. Trianea. 1994;5:323-334. [ Links ]
Normande IC, Luna FDO, Malhado ACM, Borges JCG, Junior PCV, Attademo FL, et al. Eighteen years of Antillean manatee Trichechus manatus manatus releases in Brazil: lessons learnt. Oryx. 2015;49(02):338-344. [ Links ]
Reynolds III JE. Aspects of the social behavior and herd structure of a semi-isolated colony of West Indian manatees, Trichechus manatus. Mammalia. 1981;45(4):431-451. Doi: 10.1515/mamm.1981.45.4.431. [ Links ]
Rodrigues FR, Da Silva VMF, Barcellos JFM, Lazzarini SM. Reproductive anatomy of the female Amazonian manatee Trichechus inunguis Natterer, 1883 (Mammalia: Sirenia). Anat Rec. 2008;291(5):557-564. Doi:10.1002/ar.20688. [ Links ]
Rosas FCW. Biology, conservation and status of the Amazonian manatee Trichechus inunguis. Mammal Rev. 1994;24:49-59. Doi:10.1111/j.1365-2907.1994.tb00134.x. [ Links ]
Sousa Lima RS. Comunicação Acústica em Peixes-boi (Sirenia:Trichechidae): Repertório, discriminação vocal e aplicações no manejo e conservação das espécies no Brasil (Tese Pós-Graduação). Belo Horizonte, Brasil: Instituto de Ciências Biológicas. Universidad Federal de Minas Gerais; 1999. 74 p. [ Links ]
Timm RM, Albuja L, Clauson BK. Ecology, distribution, harvest and conservation of the Amazonian manatee, Trichechus inunguis, in Ecuador. Biotropica. 1986;18(2):150-156. Doi:10.2307/2388757. [ Links ]
Wey T, Blumstein DT, Shen W, Jordán F. Social network analysis of animal behaviour: a promising tool for the study of sociality. Anim Behav. 2008;75:333-344. Doi:10.1016/j.anbehav.2007.06.020. [ Links ]

