










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.21 no.2 Bogotá May./Aug. 2016
https://doi.org/10.15446/abc.v21n2.50164
Doi: http://dx.doi.org/10.15446/abc.v21n2.50164
CARACTERIZACIÓN DE MICROSITIOS PARA EL ESTABLECIMIENTO DE PLÁNTULAS DE Espeletia uribei (Asteraceae)
Characterization of Microsites for Seedling Establishment of Espeletia uribei (Asteraceae)
Angélica María GALLEGO MAYA1, 2, María Argenis BONILLA GÓMEZ2.
1 Maestría en Ciencias-Biología, Área Curricular de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, sede Bogotá. Bogotá, Colombia.
2 Grupo de investigación Biología de Organismos Tropicales, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, sede Bogotá. Bogotá, Colombia.
For correspondence. anmgallegoma@unal.edu.co
Received: 30th April 2015, Returned for revision: 8th September 2015, Accepted: 30th October 2015. Associate Editor: Susana Feldman.
Citation / Citar este artículo como: Gallego Maya AM, Bonilla Gómez MA. Caracterización de micrositios para el establecimiento de plántulas de Espeletia uribei (Asteraceae). Acta biol. Colomb. 2016;21(2):387-398. doi: http://dx.doi.org/10.15446/abc.v21n2.50164
RESUMEN
Con el fin de determinar si los micrositios de establecimiento están limitando el reclutamiento de las plántulas y la regeneración de Espeletia uribei, un frailejón endémico y valor objeto de conservación del Parque Nacional Natural Chingaza, se evaluaron diez variables bióticas y abióticas dentro de micrositios con plántulas y micrositios vacíos. Las variables que más se relacionaron con el establecimiento fueron: la distancia al adulto reproductivo más cercano, el tipo de zona de luz "abierta" y la estructura de vegetación "herbácea" asociada. La distancia al adulto reproductivo más cercano dificulta que las semillas alcancen micrositios más alejados debido al tipo de dispersión primaria barócora de la especie. Zonas de luz "abiertas" y vegetación asociada herbácea condicionaron la capacidad de germinación y establecimiento de la especie. Los resultados mostraron que el reclutamiento de E. uribei está limitado por la disponibilidad de micrositios que reúnan estas condiciones, un factor que es importante considerar para el planteamiento de futuros proyectos de manejo para la conservación y restauración de las poblaciones de la especie.
Palabras clave: distancia al adulto, frailejón, luz, microhábitat, páramo, reclutamiento, vegetación asociada.
ABSTRACT
In order to determine whether microsites are limiting seedling recruitment and regeneration of Espeletica uribei (an endemic frailejon, a valuable species subject to conservation in the Parque Nacional Natural Chingaza) ten biotic and abiotic variables were evaluated from E. uribei seedling microsites as well as from sites without seedlings. The variables associated with seedling establishment were distance from the nearest reproductive adult, the quality of exposure to light, and associated herbaceous structure. Reduced distances from the nearest reproductive adult made it difficult for the seeds to reach distant microsites due to the principle wind dispersal type (anemochory). Open gaps and associated herbaceous structure positively affect the germination capacity and establishment of the species. The results suggest that E. uribei recruitment is limited by the availability of microsites that meet these conditions. These are important factors to consider when planning future projects for the conservation and restoration of this species.
Keywords: distance to the adult, frailejon, light, microhabitat, paramo, recruitment, associated vegetation.
INTRODUCCIÓN
La dinámica de una población puede medirse en términos de nacimientos, muertes y transiciones entre diferentes estados, tamaños o edades. En las plantas, uno de estos procesos es el reclutamiento, donde una serie de factores bióticos y abióticos determinan el número de individuos nuevos que entran a la población (Rey y Alcántara, 2000).
Han sido numerosos los estudios alrededor de las estrategias de reclutamiento que tienen las plantas para establecerse y mantenerse en los hábitats. Algunos estudios han abordado aspectos relacionados con la producción de semillas (Eriksson y Ehrlen, 1992; Caspersen y Saprunoff, 2005; Sanguinetti y Kitzberger, 2009), los mecanismos de dispersión (Montenegro, 2000; Rey y Alcántara, 2000; Velasco, 2004; Díaz y Armesto, 2007; Contreras, 2009), la formación de bancos de semillas (Montenegro, 2000; Forget et al., 2005), los procesos de germinación (Schupp, 1995;; García y Monroy, 2005; Contreras, 2009), la depredación de semillas (Ruiz, 1977; Arango, 2001; Contreras, 2009), la disponibilidad de luz (Wenny, 2000; Méndez et al., 2006), las características del suelo, el efecto de la vegetación competidora o nodriza sobre el establecimiento y desarrollo de las plántulas (Méndez et al., 2006; Mora et al., 2006a), o el arribo y la elección de micrositios adecuados (Schupp, 1995; Rey y Alcántara, 2000; Moreno, 2008).
Un micrositio se define como el ambiente tanto aéreo como subterráneo que rodea a una planta y que le permite la supervivencia y desarrollo, al proporcionarle los requerimientos suficientes para realizar al menos un evento reproductivo exitoso durante su ciclo de vida (García y Monroy, 2005). En los ecosistemas de alta montaña, por ejemplo, existe una marcada heterogeneidad ambiental, generando un mosaico de parches habitables y no habitables para las especies (Vargas y Pérez-Martínez, 2014). Esta situación hace que el reclutamiento sea limitado por la disponibilidad de micrositios óptimos para la germinación y el establecimiento.
En general, comprender los factores que determinan el reclutamiento de las plantas es necesario para entender los procesos de regeneración en las poblaciones. En este estudio, se plantea la hipótesis de que las plántulas de E. uribei están más limitadas por las características de los micrositios para el establecimiento que por la producción de semillas por las plantas adultas.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó entre febrero de 2013 y enero de 2014 en el Parque Nacional Natural Chingaza, un área protegida inscrita a la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales de Colombia (UAESPNN). El Parque está ubicado en la Cordillera Oriental, entre los departamentos de Cundinamarca (zona nororiente de Bogotá) y el departamento del Meta.
El parque Chingaza presenta un rango altitudinal que va desde los 800 hasta los 4200 m s.n.m, pero con un mayor porcentaje de zonas sobre los 3300 m s.n.m. que incluyen ecosistemas típicos de alta montaña tropical andina, caracterizados por condiciones ambientales adversas como: variaciones diarias de temperatura, alta exposición de energía radiante, estacionalidad hídrica, sequía fisiológica, bajas temperaturas en el suelo y en el aire, baja presión atmosférica (con poca disponibilidad de oxígeno), vientos fuertes y disponibilidad limitada de polinizadores (Monasterio, 1986; Mora-Osejo y Sturm, 1994; Vargas y Pedraza, 2003).
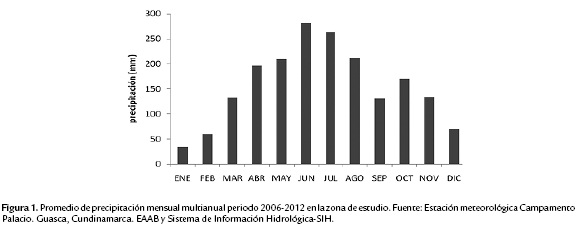
En el Parque Chingaza se presenta un régimen de lluvias unimodal-biestacional, con lluvia máxima entre junio y julio y con una época más seca entre diciembre y enero (Fig. 1). El promedio de precipitación media multianual (periodo 2006-2012) fue de 1891 mm y la temperatura promedio multianual (periodo 2008-2010) de 9.9 °C (Fuente: Estación meteorológica Campamento de Palacios, Guasca, Cundinamarca, Altitud: 3200 m s.n.m. Empresa de Acueducto y Alcantarillado de Bogotá (EAAB)-Sistema de Información Hidrológica-SIH).
La zona de estudio (4°43,318' N 73°49,961' W), es una franja de transición entre el límite superior del bosque alto andino y el subpáramo; allí E. uribei presenta su distribución natural. El área estudiada incluyó un terreno de 5200 m2 (65 * 80 m), con una altitud que oscilo entre los 3278 y 3297 m s.n.m. Con un ángulo de inclinación general de 22,78° y una pendiente de 0,42.
En el área se encontraron árboles pequeños y aislados dentro de la matriz de vegetación abierta, dominada principalmente por gramíneas, hierbas y arbustos, donde se destacan los matorrales (arbustales) con especies de la familia Asteraceae (Pentacalia, Diplostephium y Baccharis), Hypericaceae (Hypericum) y Ericaceae (Macleania, Gaultheria, Vaccinium y Pernettya spp.), así como pastizales de Calamagrostis effusa, pajonales de Agrostis sp. y arbustos como, Aragoa abietina, Diplostephium revolutum, Pernettya prostrata, Bucquetia glutinosa, entre otras plantas que no superan los cuatro metros de altura (Vargas y Pedraza, 2003).
Especie en estudio
Espeletia uribei Cuatrecasas 1978 (Asteraceae) es una especie endémica del Parque Chingaza, se encuentra entre el límite superior del bosque altoandino (3000 y 3200 m s.n.m.) y el subpáramo (3200 y 3500 m s.n.m.), así como en algunas áreas abiertas que quedan después de la destrucción del bosque, o también de forma aislada en los bosques achaparrados que crecen en los pliegues de las montañas y entre los abrigos rocosos (Vargas y Pedraza, 2003).
Espeletia uribei (Fig. 2) se distingue fácilmente de otros frailejones por presentar un tallo totalmente desnudo (sin presencia de necromasa) que alcanza hasta diez metros de altura, hojas grandes y coriáceas, e inflorescencias cortas y cubiertas por la roseta de hojas (Vargas y Pedraza, 2003). Las hojas de los adultos son glabras, es decir, que no presentan una capa densa de pelos, por lo que su estructura es más coriácea o dura a diferencia de E. grandiflora y E. killipii. Las inflorescencias son terminales con inflorescencias radiales (capítulos) inicialmente amarillas y rojizas al envejecer (Bonilla M y Gallego AG). Las semillas denominadas aquenios, son un complejo fruto-semilla. Las semillas son pequeñas de 1 x 2,5 mm de longitud y de forma ovoide triangular, la cubierta seminal es de color castaño, superficie lisa y consistencia endurecida, delgada y quebradiza. Los cotiledones y el embrión están recubiertos por una capa delgada. No presenta endospermo, su embrión es central, lineal, recto, de color blanquecino y está provisto de dos cotiledones carnosos y gruesos (Moreno, 2008).

La época de floración de la especie, durante esta investigación, inició a finales de junio hasta mediados de octubre del año 2013 y la época de dispersión de semillas fue durante la temporada seca entre los meses de septiembre de 2013 y enero de 2014. La dispersión primaria de Espeletia es barócora, donde las semillas pueden caer debajo de los padres, y una dispersión secundaria donde el viento o la escorrentía mueven las semillas a una mayor distancia de los adultos reproductivos.
Fases de campo y laboratorio
Para evaluar las características de los micrositios se sortearon de forma aleatoria 99 puntos dentro del área de estudio, caracterizando 40 micrositios de establecimiento de plántulas de E. uribei, y 59 micrositios vacíos o de no establecimiento, identificando y comparando así las condiciones que predominan cuando las plántulas de E. uribei están presentes.
Cada micrositio se delimitó con un cuadrante de 50 cm2 alrededor de la plántula o punto vacío, donde se registraron las siguientes variables: número de plántulas de E. uribei, estructura de la vegetación asociada (estratos), porcentaje de suelo desnudo, composición y número de especies asociadas, porcentaje de cobertura de cada especie asociada, porcentaje de vegetación muerta, cantidad de agua en el suelo (humedad del suelo), pH del suelo, tipo de zona de luz y distancia al adulto más cercano.
Se consideraron como plántulas los individuos que no presentaron estípite o presencia de un tallo. En cada plántula se midieron otras variables como: número de hojas, longitud de la hoja más larga y el aspecto físico de la plántula, para determinar la estructura de tamaños y el estado de las plántulas de E. uribei.
Las variables, suelo desnudo, cobertura de cada especie asociada y vegetación muerta se midieron como porcentaje de cobertura usando una cuadrícula; para la variable humedad del suelo, se tomó una muestra en la que se registró el peso húmedo y el peso seco, las muestras se analizaron en el laboratorio de Ecología de la Universidad Nacional. El suelo se secó en un horno de convección natural a temperatura constante de 65 °C, durante cinco días. A la muestra de suelo seco se le retiraron 5 g. que se disolvieron en agua destilada para medir posteriormente el pH del suelo.
La variable tipo de zona de luz se evaluó de acuerdo con la presencia-ausencia de luz que generó la estructura y cobertura de las especies asociadas, definiendo tres categorías: 1) abierta, se refiere a una zona donde llega directamente la radiación solar; 2) parcialmente cerrada, cuando un 50 % de la zona presenta sombra y 3) cerrada cuando es muy baja o nula la cantidad de luz que llega al suelo.
La variable estructura de la vegetación asociada, tuvo tres categorías: 1) herbácea: cuando las plantas presentaron un altura máxima de un metro, 2) arbustiva: se refiere a plantas entre uno y tres metros de altura, 3) herbáceo-arbustivo: cuando el 50 % de las plantas presentes eran hierbas de porte pequeño, y había presencia de un arbusto. Para la variable de composición de especies asociadas se usó la guía de plantas ilustradas de Chingaza (Madriñan, 2004), así como la recolección de ejemplares (permiso de estudio 003 de 13 de febrero de 2013. Parques Nacionales Naturales de Colombia) para la posterior identificación botánica en el herbario de la Universidad Nacional.
Análisis estadístico
Para determinar si hubo diferencias significativas entre los micrositios de establecimiento de plántulas y los micrositios vacíos se realizó estadística descriptiva, pruebas de normalidad, homocedasticidad y análisis de varianza. Las pruebas de Wilcoxon y Kolmogorov-Smirnov se aplicaron para las variables: porcentaje de vegetación muerta, pH, porcentaje de suelo desnudo y distancia al adulto más cercano; la prueba t-Test para las variables: cantidad de agua en el suelo y número de especies asociadas; y el Test de Chi2 y test G con corrección de Williams, para las variables cualitativas tipo de vegetación asociada y tipo de zona de luz.
Para evaluar cuáles variables tuvieron mayor relación y probabilidad de ocurrir con el establecimiento de las plántulas se realizó una regresión logística.
Los análisis estadísticos y gráficos, se hicieron usando el programa R Versión 3.0.0 (R Development Core Team, 2008) y RWizard Software (Guisande et al., 2014), empleando y modificando los scripts de Guisande et al. (2011).
RESULTADOS
En los micrositios de establecimiento se encontraron en promedio dos plántulas, con un valor máximo de 13 plántulas (N = 40; máx = 13; mín = 1; SD ± 2,79). El tamaño de las plántulas, de acuerdo con la longitud de la hoja más larga, fue en promedio de 10,97 cm (N = 40; máx = 31; mín = 1,5; SD ± 7,42), con cuatro hojas en promedio por individuo (N = 40; máx = 15; mín = 2; SD ± 1,67). Estos datos presentaron un intervalo amplio y una desviación estándar alta, sugiriendo que las plántulas tuvieron una distribución desigual de tamaños.
El 53,8 % de las plántulas evaluadas no presentó ningún daño, mientras que 34,6 % presentaron hojas secas y 11,5 % tuvieron señales de herbivoría.
El porcentaje de vegetación muerta en los micrositios con plántulas fue en promedio de 16 % (N = 40; máx = 30; mín = 0; SD ± 7,55) y para micrositios vacíos de 23 % (N = 59; máx = 80; min = 0; SD ± 16,40). No hubo diferencias significativas en el porcentaje de vegetación muerta entre estos dos tipos de micrositios (Wilcoxon W = 1389; p = 0,1365 y Kolmogorov-Smirnov D = 0,2398; p = 0,12; Fig. 3).
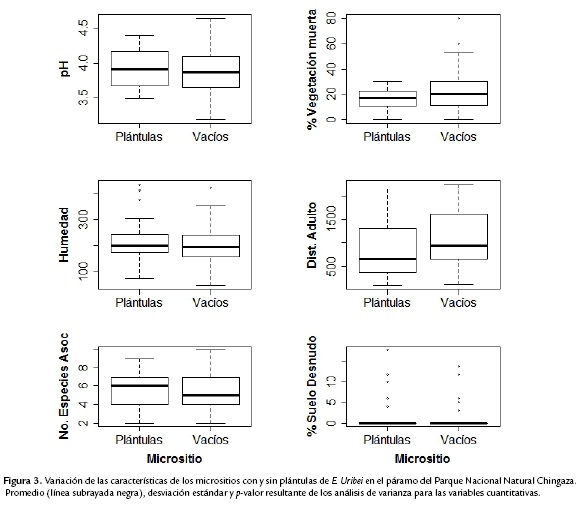
El pH del suelo no presentó diferencias significativas entre micrositios vacíos y micrositios con plántulas (Wilcoxon W= 1136,5; p = 0,75; Kolmogorov-Smirnov D = 0,1021; p = 0,96; Fig. 3).
La cantidad de agua en el suelo en micrositios con plántulas fue en promedio de 213,05 g (N = 40; máx = 433,84; mín = 74,53; SD ± 77,19) y para micrositios vacíos de 199,51 g (N = 59, máx = 421,55; mín = 49,41; SD ± 66,85). Sin embargo, no hubo diferencias significativas entre los micrositios (t = -0,8405; df = 79,157; p = 0,4032; Fig. 3).
En cuanto al porcentaje de suelo desnudo, no hubo diferencias entre micrositios vacíos y micrositios con plántulas (Wilcoxon W = 1144,5; p = 0,69; Kolmogorov-Smironov D = 0,0822; p = 0,99), sus valores medios fueron de 1,10 % y 1,55 % respectivamente (Fig. 3).
El número promedio de especies asociadas fue de cinco en ambos tipos de micrositios (m. vacíos: N = 59; máx = 10; mín = 2; SD ± 1,80; m. llenos: N = 40; máx = 9; mín = 2; SD ± 1,89; Fig. 3) y no hubo diferencias significativas entre micrositios con plántulas y micrositios vacíos (t-Test, t = -0,2376; df = 80,95; p = 0,8128).
La distancia al adulto más cercano tuvo un valor promedio de 1124,22 cm para micrositios vacíos (N = 59; máx = 2250; mín = 100; SD ±599,10) y de 825,5 cm en micrositios de establecimiento de plántulas (N = 40; máx = 2150; mín = 90; SD ±560,52), presentándose diferencias significativas entre los micrositios (Wilcoxon W = 1547,5; p = 0,008; Fig. 3).
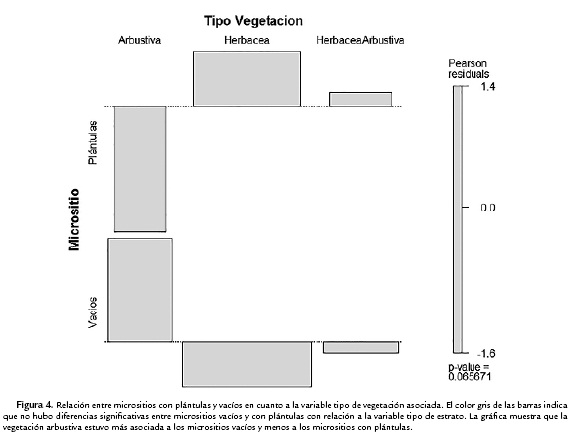
El tipo de vegetación asociada, tanto para los micrositios vacíos como para micrositios con plántulas fue el estrato herbáceo. No hubo diferencias significativas en el tipo de vegetación entre los micrositios (estadístico X2 de Pearson = 5,4462; df = 2; p = 0,065; test G sin corrección G = 6,1723; X2 df = 2; p = 0,045). Sin embargo, la categoría "estrato arbustivo" fue menos frecuente en los micrositios con plántulas (Fig. 4).
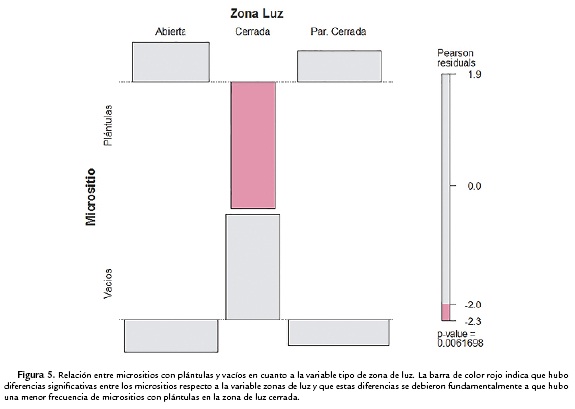
En la variable zona de luz hubo diferencias significativas entre los dos tipos de micrositios (estadístico X2 de Pearson d = 10,1762; df = 2; p = 0,0070), la categoría zona de luz cerrada no se presentó en ningún micrositio donde hubo plántulas de E. uribei (Fig. 5).
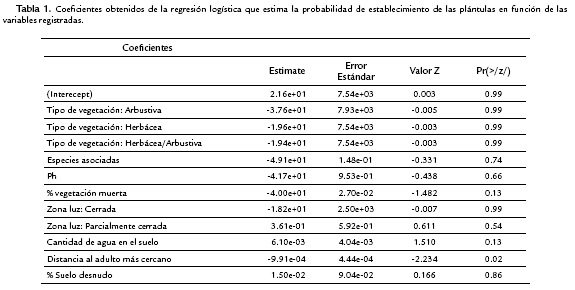
El establecimiento de las plántulas disminuyó cuando el tipo de vegetación arbustiva estuvo presente (r = -3,76e+01; p = 0,99). Igualmente, la mayor probabilidad de que hubiera establecimiento de plántulas estuvo asociada con la variable zona de luz parcialmente cerrada (r = 3,618e-03; p = 0,54). La distancia al adulto reproductivo más cercano fue la variable más relacionada con el establecimiento de las plántulas (r = -9,91e-04; p = 0,02; Tabla 1).
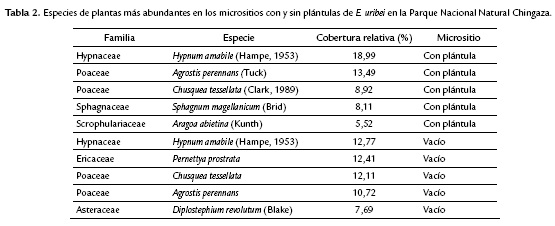
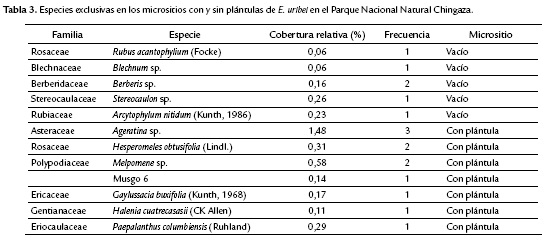
Se registraron un total de 48 especies asociadas a los micrositios, perteneciente a 23 familias y 33 géneros. Las especies más abundantes y comunes entre los micrositios vacíos y llenos fueron: Hypnum amabile (Hampe, 1869), Chusquea tessellata (Munro, 1989), Agrostis perennans (Tuck) y Pernettya prostrata (Cav.DC.)(Tabla 2). Las coberturas y frecuencias de las especies exclusivas fueron muy bajas (Tabla 3) y no hubo diferencias significativas entre los porcentajes de cobertura relativa de las especies y los tipos de micrositios (Wilcoxon W = 1280; p = 0,1851; Kolmogorov-Smirnov D = 0,2553; p = 0,09341).
DISCUSIÓN
El establecimiento de los nuevos reclutas, o individuos que entran a la población, es un proceso crítico en la vida de las plantas debido a que depende en gran parte de micrositios óptimos que brinden nutrientes y protección necesaria para la germinación, el crecimiento y la supervivencia de las semillas y las plántulas (Eriksson y Ehrlen, 1992; Chambers y MacMahon, 1994; Begon et al., 1995; Schupp, 1995, Chambers et al., 1999; Fenner y Thompson 2005).
En ambientes con condiciones ambientales drásticas como los páramos, el reclutamiento de las plántulas es un factor limitante en la dinámica de las poblaciones, sin embargo, son poco los estudios sobre las estrategias que les permiten a las semillas y las plántulas persistir en estos ambientes (Vargas y Pérez-Martínez, 2014). Entre los estudios se reportan el género Espeletia (Smith, 1981), Senecio keniodendro en los páramos africanos (Smith y Young 1982), P. dasylirioides (Augspurger, 1985), Puya spp. (Miller, 1987; Mora et al., 2006b), E. Spicata (Estrada y Monasterio, 1988), E. granfilora (Fagua y González, 2007) y E. killipii (Moreno, 2008).
En el páramo las semillas y plántulas están sometidos a temperaturas cercanas al punto de congelamiento, periodos de sequía con altas temperaturas en la superficie del suelo, inestabilidad del sustrato, bajos niveles de oxígeno y suelos ácidos que producen gran cantidad de mortalidad de plántulas (Vargas y Pérez-Martínez, 2014). Para superar estas restricciones, muchas plantas requieren condiciones más favorables que son encontradas a pequeña escala (de semilla y de plántula) en los llamados "micrositios favorables" para la germinación y establecimiento. Estos micrositios son un factor decisivo en la regeneración de muchas especies de plantas en los páramos y constituye un tema de estudio muy importante para la conservación y restauración ecológica en estos ecosistemas estratégicos (Vargas y Pérez-Martínez, 2014).
Las variables que determinaron el establecimiento exitoso de las plántulas de E. uribei en los micrositios de establecimiento fueron la zona de luz abierta, el tipo de vegetación asociada herbácea y la distancia al adulto más cercano. Aunque, las otras variables evaluadas no presentaron relaciones de significancia, es necesario tener en cuenta que en ocasiones pueden llegar a afectar la germinación y el establecimiento de las plántulas.
El porcentaje de vegetación muerta no tuvo en este estudio una relación significativa con el establecimiento; sin embargo, puede llegar a afectar las etapas de germinación y emergencia de las plántulas de forma positiva, cuando su presencia determina la cantidad y calidad de nutrientes y minerales presentes en el suelo, y que serán usados por las plántulas en sus procesos fisiológicos, así como el papel que ejercen en la regulación de condiciones microambientales (McGee y Birmingham, 1997; Caspersen y Saprunoff, 2005). Al contrario, para otras plántulas es mejor contar con sustratos con poca vegetación muerta, pues, algunas semillas son incapaces de penetrar las capas gruesas de la hojarasca y acceder al suelo mineral (McGee y Birmingham, 1997; McGee, 2001).
Aunque las plántulas de E. uribei no estuvieron limitadas por el porcentaje de vegetación muerta se observó que en los micrositios de establecimientos hubo un porcentaje bajo de restos de vegetación (16 % en promedio). Esto puede estar relacionado con una mayor supervivencia de las plántulas en los microhábitats con acumulación de vegetación muerta con respecto a zonas de suelo totalmente desnudos o pedregosos (Marañón et al., 2004), debido a que se reducen las amplitudes de temperatura, aumenta la humedad del suelo y se regulan los niveles de luz según la masa y el espesor de la cubierta (Donath y Eckstein, 2010). Así mismo, al ser una capa delgada, no limitó el crecimiento de las plántulas.
El pH del suelo no presentó diferencias entre los micrositios vacíos y llenos, ni relevancia sobre el establecimiento, sino que correspondió a las características generales del suelo de Chingaza, los cuales se caracterizan por ser ácidos, con baja saturación de bases, alta capacidad catiónica de cambio, pobres en sílice, ricos en aluminio, con alta capacidad de absorción de agua, con predominio de arcillas, y, con sustancias húmicas muy ácidas y poco evolucionadas (Malagón y Pulido, 2000). No obstante, se debe reconocer que la heterogeneidad en la composición química del suelo tiene un conocido efecto diferencial sobre el reclutamiento de las especies. Es conocida la distribución preferente de las especies de plantas en determinados tipos de suelos; por ejemplo las calcícolas en suelos calizos de pH básico, las silicícolas en suelos silíceos de pH ácido, y las halófilas en suelos salinos (Marañón et al., 2004). En el caso de los suelos ácidos, la disponibilidad elevada del aluminio y sus efectos tóxicos, parece ser el principal limitante para la germinación de las semillas y el establecimiento de las plántulas, de las especies más sensibles (Marañón et al., 2004) pero como se mencionó para este estudio el pH no fue transcendental, valdría la pena evaluar otras variables químicas del suelo y su efecto en el establecimiento.
El porcentaje de suelo desnudo, que ha sido reportado para algunas especies como un factor positivo para el reclutamiento (Mora et al., 2006), no mostró asociación con los micrositios, y al contrario, sus valores fueron muy bajos y no significativos para el establecimiento, ya que de acuerdo con nuestras observaciones durante el trabajo de campo, E. uribei se ve favorecido por un suelo cubierto que lo protege de las heladas, la alta radiación ultravioleta y de posibles depredadores de semillas. En contraste, Moreno (2008) encontró que para E. grandiflora, también estudiada en el Parque Chingaza, la mayor germinación de semillas y establecimiento de plántulas se presentó en zonas libres de vegetación, en donde la capa edáfica fue alterada, es decir, en micrositios donde la cobertura de suelo desnudo estuvo entre 50-70 %.
La cantidad de agua en el suelo no constituyó un factor relevante en el establecimiento de las plántulas de E. uribei. Correspondió al hábitat de esta especie que se caracteriza por ser un zona semihumeda, ligeramente inclinada y de suelos bien drenados (Vargas y Pedraza, 2003).
La distancia al adulto reproductivo más cercano fue un factor relacionado con el proceso de reclutamiento de la especie, ya que hubo diferencias significativas con los micrositios vacíos, la mayoría de las plántulas de E. uribei se encontraron cerca a los adultos parentales, a mayores distancias era menor la posibilidad de encontrar plántulas, esto es efecto primordialmente del mecanismo de dispersión primaria barocora, donde las semillas pueden caer debajo de los padres, y una dispersión secundaria donde el viento o la escorrentía mueven las semillas a una mayor distancia de los adultos reproductivos (Zasada et al., 1992; Chambers y MacMahon, 1994).
La variable de vegetación asociada a los micrositios se midió en dos aspectos, la estructura de la vegetación (estratos) y la composición y cobertura de las especies. En cuanto a la composición y cobertura no se encontró que hubiera relación con el establecimiento de las plántulas de E. uribei pues la mayoría de las especies fueron comunes en ambos micrositios (vacíos - con plántulas) y las que fueron exclusivas presentaron una cobertura inferior al 1 % y una frecuencia muy baja. Aunque estudios desarrollados en especies de frailejón asocian la presencia de musgos a la germinación y establecimiento de las plántulas (Moreno, 2008), en este trabajo se encontró relación entre la presencia de musgos y la presencia de plántulas de E. uribei.
Con respecto a la estructura y tamaño de la vegetación asociada a los micrositios, hubo una relación significativa con el establecimiento. En este sentido, los micrositios con vegetación de porte herbáceo tuvieron mayor establecimiento de plántulas en comparación con los micrositios donde la vegetación acompañante era de porte arbustivo. En condiciones microclimáticas de estrés, las plantas vecinas actúan a menudo como facilitadoras o como competidoras (Tielborger y Kadmon, 2000; Brooker et al., 2008). La competencia de las plántulas con la vegetación asociada al micrositio se presenta principalmente en tres casos: i) a nivel de sistema radical disputando nutrientes y agua, ii) por la luz, y iii) por sus efectos alelopáticos sobre el crecimiento de las plántulas (Mora et al., 2006). En el caso de E. uribei se halló que las hierbas, a diferencias de los arbustos, facilitaron el establecimiento de las plántulas, porque permitieron una mayor disponibilidad de luz.
Una condición que también estuvo asociada fuertemente a los micrositios de establecimiento de E. uribei fue el tipo de zona de luz abierta. Esto se relacionó con la influencia que tiene el tipo y la cantidad de radiación disponible sobre numerosos procesos fisiológicos, morfogenéticos y reproductivos de las plantas (Kohen et al., 1995). Las plántulas de E. uribei, se establecieron en micrositios con vegetación herbácea y zonas de luz abiertas, que en conjunto proporcionaron las condiciones necesarias para el reclutamiento.
Moreno (2008) reportó que los micrositios adecuados para la germinación y establecimiento de Espeletia killipii en Chingaza deben tener espacios abiertos y asociados a especies de Sphagnum magellanicum y una cobertura no muy densa de otras especies para su establecimiento. E. grandiflora igualmente necesita de microclaros. Condiciones similares para E. uribei quien requiere de zonas abiertas y de cobertura vegetal de tipo herbáceo para establecerse.
La limitación sobre el reclutamiento de E. uribei concuerda con lo que varios autores proponen respecto al reclutamiento de otras especies en ecosistemas de alta montaña (Monasterio, 1986; Guariguata y Azócar, 1988; Fagua, 2002; Mora et al., 2006b; Graae et al., 2011). En dichos estudios, se plantea que el establecimiento de las plántulas se presenta preferiblemente bajo el abrigo de otras plantas o de rocas, las cuales forman micrositios protegidos y húmedos, donde condiciones como el sobrecalentamiento de la superficie del suelo durante el día, las temperaturas de congelamiento en la noche, los períodos de baja disponibilidad de agua y la inestabilidad del sustrato son más tolerables.
Con respecto a la estructura de tamaño se encontró un tamaño desigual en las plántulas de E. uribei entre los micrositios, esto puede deberse al aporte de semillas de diferentes épocas reproductivas; o ser evidencia de la conveniencia relativa de los micrositios, en términos de los recursos que le proporcionan a la semilla en cada fase de su desarrollo, siendo así convenientes para su germinación pero no necesariamente para su establecimiento, lo que Schupp (1995) llamó "conflicto semilla-plántula". Por esto, es posible que las diferencias en la estructura de tamaños de las plántulas de E. uribei estudiadas sea el resultado de las condiciones del micrositio, las cuales favorecieron o retardaron la emergencia de las plántulas, así como su crecimiento, su supervivencia y transición a las siguientes etapas.
La heterogeneidad en la estructura de tamaño en las plántulas de E. uribei es una situación común dentro de las poblaciones vegetales y se asocia a diferencias en el acervo genético de los individuos, el tiempo de germinación, las tasas de crecimiento, interacciones de competencia, de facilitación (Hutchings, 1997; Silvertown y Charlesworth, 2001; Lopera, 2010), efectos denso-dependientes, depredación por herbívoros, el ataque de patógenos, la caída de ramas o la falta de recursos suficientes para el crecimiento (Gilbert et al., 2001).
Adicionalmente, al evaluar el estado de las plántulas durante el periodo de estudio, se encontró que la mayoría de los nuevos individuos no presentaron signos de daño o señales de herbivoría u hongos, lo que indica que hasta el momento, los micrositios trabajados fueron adecuados para el establecimiento de las plántulas.
CONCLUSIONES
Espeletia uribei presentó requerimientos particulares para establecerse, lo que mostró que el patrón de reclutamiento de las plántulas de esta especie está determinado por las condiciones del micrositio. Entre las condiciones que debe reunir un micrositio para el establecimiento de E. uribei se encontraron principalmente la vegetación herbácea asociada y la disponibilidad marcada de luz.
AGRADECIMIENTOS
Las autoras agradecemos al programa de Jóvenes Investigadores e Innovadores de Colciencias, a la Dirección de Investigación Sede Bogotá (DIB) y a la Facultad de Ciencias de la Universidad Nacional de Colombia por el financiamiento para la realización de esta investigación otorgado a AM Gallego dentro de la convocatoria 566 año 2012 (Convenio 0729 de 2012). A la Subdirección de Gestión y Manejo de Áreas Protegidas de Parques Nacionales Naturales de Colombia por el permiso de estudio de investigación científica en diversidad biológica 003 de 13 de febrero de 2013. Al Parque Nacional Natural Chingaza, a los funcionarios y guardabosques del Parque que nos brindaron su ayuda y compañía en el desarrollo de la investigación en la fase de campo.
REFERENCIAS
Arango S. El impacto del efecto de bosque, los depredadores de semillas y la herbivoría de plántulas, en la regeneración de árboles en fragmentos remanentes de bosque tropical montano Andes Centrales Colombia (Monografía). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2001. 183 p. [ Links ]
Begon M, Harper J, Townsend C. Ecología individuos, poblaciones y comunidades. Barcelona: Ediciones Omega. S.A.; 1995. p. 5-165. [ Links ]
Brooker RW, Maestre FT, Callaway RM, Lortie CL, Cavieres LA, Kunstler G, et al. Facilitation in plant communities: the past, the present, and the future. J Ecol. 2008;96(1):18-34. Doi:10.1111/j.1365-2745.2007.01295.x. [ Links ]
Caspersen JP, Saprunoff M. Seedling recruitment in a northern temperate forest: the relative importance of supply and establishment limitation. Can J For Res. 2005;4(35):978-989. Doi:10.1139/X05-024. [ Links ]
Chambers JC, MacMahon JA. A Day in the Life of a Seed: Movements and Fates of Seeds and Their Implications for Natural and Managed Systems. Annu Rev Ecol Syst. 1994;25:263-292. Doi:10.1146/annurev.es.25.110194.001403. [ Links ]
Chambers J, Vander Wall S, Schupp E. Seed and Seedling Ecology of Piñon and Juniper Species in the Pygmy Woodlands of Western North America. Bot Rev. 1999;65(1):1-38. Doi:10.1007/BF02856556. [ Links ]
Contreras LF. Efecto de la depredación pre-dispersión de semillas sobre la germinación y supervivencia de plántulas (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2009. 96 p. [ Links ]
Díaz M, Armesto J. Limitantes físicos y bióticos de la regeneración arbórea en matorrales sucesionales de la Isla Grande de Chiloé, Chile. Rev Chil Hist Nat. 2007;80:13-26. Doi:10.4067/S0716-078X2007000100002. [ Links ]
Donath T, Eckstein L. Effects of bryophytes and grass litter on seedling emergence vary by vertical seed position and seed size. Plant Ecol. 2010;207:257-268. Doi:10.1007/s11258-009-9670-8. [ Links ]
EAAB Empresa de Acueducto y Alcantarillado de Bogotá ESP. Sistema de Información Hidrológica, Estación Campamento Palacios, Parque Nacional Natural Chingaza; 2013. [ Links ]
Eriksson O, Ehrlén J. Seed and microsite limitation of recruitment in plant populations. Oecología. 1992;91:360-364. Doi:10.1007/BF00317624. [ Links ]
Fagua JC. Estrategias de reproducción sexual de una población de Espeletia grandiflora H & B, en el Parque Nacional Natural Chingaza (Cundinamarca) (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2002. 136 p. [ Links ]
Fagua JC, González V. Growth Rates, Reproductive Phenology, and Pollination Ecology of Espeletia grandiflora (Asteraceae), a Giant Andean Caulescent Rosette. Plant Biol. 2007;9:127-135. Doi:10.1055/s-2006-924544. [ Links ]
Fenner M, Thompson K, editors. The Ecology of Seeds. Cambridge: Cambridge University Press; 2005. 250 p. [ Links ]
Forget P, Lambert J, Hulme P, Vander Wall SB, editors. Seed Fate: Predation, Dispersal and Seedling Establishment. Cambridge: CABI Publishing; 2005. p. 1-31. [ Links ]
García SR, Monroy AA. Micrositios del pasto navajita (Bouteloua gracilis) en comunidades de pastizal y de matorral del altiplano Mexicano. TIP Rev Esp Cienc Quim Biol. 2005;8:61-70. [ Links ]
Gilbert GS, Harms KE, Hamill DN, Hubbell SP. Effects of seedling size, El Niño drought, seedling density, and distance to nearest conspecific adult on 6-year survival of Ocotea whitei seedlings in Panamá. Oecologia. 2001;127:509-516. Doi:10.1007/s004420000616. [ Links ]
Graae B, Ejrnaes R, Lang SI, Meineri E, Ibarra PT, Bruun HH. Strong microsite control of seedling recruitment in tundra. Oecología. 2011;166:565-576. Doi:10.1007/s00442-010-1878-8. [ Links ]
Guariguata M, Azocar A. Seed Bank Dynamics and Germination Ecology in Espeletia timotensis (Compositae), an Andean Giant Rosette. Biotropica. 1988;1(20):54-59. [ Links ]
Guisande C, Vaamonde A, Barreiro A. Tratamiento de datos con R, STATISTICA Y SPSS. España: Ediciones Díaz de Santos; 2011. 996 p. [ Links ]
Hutchings MJ. The structure of plant populations. En: Crawley MJ, editor.Plant Ecology. 2 ed. Oxford: Blackwell Scientific Publishing; 1997. 45 p. [ Links ]
Kohen E, Santus R, Hirschberg JG. Photobiology. London: Academic Press; 1995. p. 227-257. [ Links ]
Lopera MC. Estructura poblacional, crecimiento y supervivencia en los bancos de plántulas del castaño Scleronema micranthum en un bosque amazónico (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2010. 118 p. [ Links ]
Madriñan S. Flora ilustrada del Páramo de Chingaza: guía de campo plantas comunes. Bogotá: Universidad de los Andes; 2004. 64 p. [ Links ]
Malagón D, Pulido C. Suelos del páramo colombiano. In: Rangel O, editor. Colombia. Diversidad Biótica III. La Región de Vida Paramuna. Bogotá: Universidad Nacional de Colombia; 2000. 15 p. [ Links ]
Marañón T, Camarero JJ, Castro J, Díaz M, Espelta JM, Hampe A, et al., Stand-level effects on the role of decaying logs as vascular plant habitat in Adirondack northern hardwood forests. J Torrey Bot Soc. 2001;128:370-380. Doi: 10.2307/3088669. [ Links ]
McGee GG. Stand-level effects on the role of decaying logs as vascular plant habitat in Adirondack northern hardwood forest. J Torrey Bot Soc. 2001;128:370-380. [ Links ]
McGee GG, Birmingham JP. Decaying logs as germination sites in northern hardwood forests. N J Appl For. 1997;14:178-182. [ Links ]
Méndez M, Dorantes A, Dzib G, Argaez, Duran R. Germinación y establecimiento de plántulas de Pterocereus gaumeri, una cactácea columnar, rara y endémica de Yucatán México. Bol Soc Bot Méx. 2006;79:33-41. [ Links ]
Monasterio M. Adaptive Strategies of Espeletia in the Andean desert páramo. En: Vuilleumier F, Monasterio M, editores. High altitude tropical biogeography. USA: Oxford University Press; 1986. p. 267-303. [ Links ]
Montenegro, A. Estrategias de dispersión y regeneración por bancos de semillas en dos comunidades de bosque alto andino, Embalse de San Rafael, La Calera, Cundinamarca (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2000. 124 p. [ Links ]
Mora A, Valdez JI, Pérez G, Musálem MA, Vaquera H. Establecimiento y desarrollo de plántulas de Tabebuia rosea (Bignoniaceae) en una selva sub-caducifolia manejada en la costa Pacífica de México. Rev Biol Trop. 2006;54(4):1215-1225. [ Links ]
Mora F, Chaparro H, Vargas O, Bonilla MA. Dinámica de la germinación, latencia de semillas y reclutamiento de plántulas en Puya cryptantha y P. trianae, dos rosetas gigantes de los páramos colombianos. Ecotropicos. 2006;20(1):31-40. [ Links ]
Mora-Osejo LE, Sturm H. Clima. In: Estudios Ecológicos del Páramo y del Bosque Altoandino, Cordillera Oriental de Colombia. Bogotá: Academia Colombiana de Ciencias Exactas, Físicas y Naturales; 1994. p. 15-34. [ Links ]
Moreno LA. Estrategias de Reclutamiento de Espeletia killipii y E. grandiflora en el Parque Nacional Natural Chingaza (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2008. 238 p. [ Links ]
Rey P, Alcántara J. Recruitment dynamics of a fleshy-fruited plant (Olea europaea) connecting patterns of seed dispersal to seedling establishment. J Ecol. 2000;88(4):622-633. Doi:10.1046/j.1365-2745.2000.00472.x. [ Links ]
Ruiz LF. Efecto de la depredación pre-dispersión de semillas sobre la germinación y supervivencia de plántulas de Aniba perutilis (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 1977. 198 p. [ Links ]
Sanguinetti J, Kitzberger T. Efectos de la producción de semillas y la heterogeneidad vegetal sobre la supervivencia de semillas y el patrón espacio-temporal de establecimiento de plántulas de Araucaria araucana. Rev Chil Hist Nat. 2009;82(3):319-335. Doi:10.4067/s0716-078x2009000300001. [ Links ]
Schupp EW. Seed-seedling conflicts, habitat choice, and patterns of plant recruitment. A J Bot. 1995;82(3):399-409. [ Links ]
Silvertown J, Charlesworth D. Introduction to plant population biology. USA: Blackwell Science; 2001. 360 p. [ Links ]
Tielborger K, Kadmon R. Temporal environmental variation tips the balance between facilitation and interference in desert plants. Ecology. 2000;81:1544-1553. [ Links ]
Vargas O, Pedraza P. Parque Nacional Natural Chingaza. Bogotá: Gente Nueva Editorial; 2003. 228 p. [ Links ]
Vargas O, Pérez-Martínez LV, editores. Semillas de plantas de páramo: ecología y métodos de germinación aplicados a la restauración ecológica. Bogotá: Universidad Nacional de Colombia; 2014. p. 17-63. [ Links ]
Velasco L. Dinámica de la dispersión de plantas ornitócoras, reclutamiento y conectividad en fragmentos de bosque altoandino secundario Reserva Natural Protectora Cogua Cundinamarca (Trabajo de Grado). Bogotá: Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia; 2004. 136 p. [ Links ]
Wenny D. Seed dispersal, seed predation, and seedling recruitment of a neotropical montane tree. Ecol Monogr. 2000;70(2):331-351. [ Links ]
Zasada JC, Sharik TL, Nygren M. The reproductive process in boreal forest trees. In: Shugart HH, Leemans R, Bonan GB, editors. A systems analysis of the global boreal forest. Cambridge: Cambridge University Press; 1992. p. 20-43. [ Links ]

