










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.21 no.2 Bogotá May./Aug. 2016
https://doi.org/10.15446/abc.v21n2.43814
Doi: http://dx.doi.org/10.15446/abc.v21n2.43814
DISTRIBUCIÓN ESPACIAL, COMPOSICIÓN Y DENSIDAD DE EDAFOFAUNA EN HOJARASCA DE BOSQUE Y CAFETAL (MONTENEGRO, COLOMBIA)
Spatial Distribution, Composition and Density of the Leaflitter Edaphofauna in Forest and Coffee Plantation (Montenegro, Colombia)
Diana Marcela RUEDA-RAMÍREZ1, Amanda VARELA2.
1 Laboratorio de Acarología, Departamento de Entomología y Acarología, Escola Superior de Agricultura Luiz de Queiroz (ESALQ), Universidade de São Paulo (USP), Piracicaba. Av. Pádua Dias, 11, Edificio Prof. Salvador de Toledo Piza Jr. (Pavilhão de Zoologia). São Paulo, Brasil.
2 Laboratorio de Ecología de Suelos y Hongos Tropicales, Departamento de Biología, Pontificia Universidad Javeriana. Carrera 7 nº. 43-82, edificio 53, laboratorio 406 B. Bogotá, Colombia.
For correspondence. dianaru@gmail.com
Received: 16th June 2014, Returned for revision: 16th June 2015, Accepted: 23th November 2015. Associate Editor: Víctor López Martínez.
Citation / Citar este artículo como: Rueda-Ramírez DM, Varela A. Distribución espacial, composición y densidad de edafofauna en hojarasca de bosque y cafetal (Montenegro, Colombia). Acta biol. Colomb. 2016;21(2):399-412. doi: http://dx.doi.org/10.15446/abc.v21n2.43814
RESUMEN
Se determinaron los cambios en composición, densidad y distribución espacial de la edafofauna de hojarasca entre dos usos de suelo (cafetal y bosque subandino) para entender el efecto del uso de suelo. Asimismo, se establecieron las relaciones entre las variables de la edafofauna y atributos de la hojarasca. Se colectaron muestras de hojarasca superficial durante la época seca y se extrajo la edafofauna, manualmente y mediante embudos Berlese modificados. Esta se identificó hasta el menor nivel taxonómico posible y se clasificó en grupos funcionales. La composición faunística fue diferente entre los usos de suelo. La similitud en la composición fue menor al 50 % en dos de los tres grupos funcionales encontrados, pero la densidad total no presentó diferencias. En general hubo coeficientes de variación altos en la densidad faunística intermuestral para los diferentes grupos taxonómicos y, en consecuencia, una baja detectabilidad del patrón espacial. Sin embargo, se detectó un patrón espacial en la abundancia para Coleoptera, larvas de Diptera y biomasa de hojarasca, en el cafetal. La estructura fue más difícil de detectar en bosque, por los pequeños parches en este. La varianza de la abundancia fue mayor en el cafetal, para todos los grupos, excepto Acaridida y Formicidae. La densidad de Formicidae aumentó al hacerlo la humedad de la hojarasca, pero solo en cafetal. Se concluye que el establecimiento del cafetal afecta de manera diferencial a taxones y grupos funcionales de la edafofauna. Esto sienta las bases para definir los efectos del cambio de uso de suelo y los factores ambientales asociados con la distribución espacial de la edafofauna.
Palabras clave: análisis espacial, cafetal, geoestadística, grupos funcionales, SADIE.
ABSTRACT
Changes in composition, density and spatial distribution on the litter edaphic fauna between two land uses (a coffee plantation and a tropical Andean forest) were established, in order to understand the soil use effects. Also, the relationships between the biotic variables and some litter features were evaluated. Superficial litter samples were collected during dry season. The edaphic fauna was extracted manually and with modified Berlese funnels; it was identified up to the lower taxonomic level and classified in functional groups. Differences in fauna composition were observed between the two land uses. Composition similarity was lower than 50 % for almost all functional groups found, but total density did not differ between the land uses. In general, there was a high variation in density among samples resulting in higher coefficients of variation for different taxonomic groups and consequently low detectability of spatial pattern. However a spatial pattern was detected only for Coleoptera, Diptera larvae and litter biomass in the coffee plantation. The structure in forest was more difficult to detect, due to the smaller patches on it. The variance of abundance was significantly higher in the coffee plantation for all groups, except Acaridida and Formicidae. Density of Formicidae increased with the enhancement of litter moisture, but only in the coffee plantation. In conclusion, the establishment of coffee plantation differentially affects taxa and functional groups of soil fauna. This is the basis for establishing the effects of soil change use, and the environmental factors associated with spatial distribution of edaphic fauna.
Keywords: coffee plantation, functional groups, geostatistics, SADIE, spatial analysis.
INTRODUCCIÓN
Cuando se presenta eliminación de la vegetación nativa, acompañada de la mecanización y el uso de productos de síntesis química, se evidencia intensificación en el uso del suelo y pérdida de biodiversidad (De la Rosa y Negrete-Yankelevich, 2012). También se presentan cambios en la distribución de la vegetación, y en consecuencia, en la calidad del suelo. Estas modificaciones pueden afectar a la edafofauna, que actúa de manera importante en procesos que varían espacialmente, como la descomposición, la herbivoría y el ciclo de nutrientes (Thiele-Bruhn et al., 2012), y, de esta manera, pueden aumentar la propensión al surgimiento de insectos plaga e infecciones por patógenos microbianos (Matson et al., 1997). Desde años atrás se ha reconocido la importancia de la presencia de heterogeneidad espacial y temporal, que puede influir sobre procesos ecosistémicos (Crist, 1998; Ettema y Wardle, 2002).
Los estudios de estructura espacial de la edafofauna se han centrado en la comparación entre usos de suelo: sabana nativa colombiana y pastizal (Jiménez et al., 2001; Jiménez y Rossi, 2006); campo agrícola abandonado y área boscosa adyacente dominada por arce rojo (Görres et al., 1998); pastizal de Raygrass y bosque de podocarpos/latifolias (Ettema y Yates, 2003), o entre etapas sucesionales (Negrete-Yankelevich et al., 2006; De la Rosa y Negrete-Yankelevich, 2012). En cuanto a los organismos se ha trabajado con lombrices (Decaëns y Rossi, 2001; Jiménez et al., 2001; Jiménez y Rossi, 2006), nematodos (Görres et al., 1998; Ettema y Yates, 2003) y colémbolos (Chust et al., 2003), encontrándose un impacto difícil de definir en las lombrices, una estructura más fina en bosque para los nematodos, y para los colémbolos, un papel importante como descriptores del paisaje. Particularmente en Colombia los estudios se han enfocado en las lombrices de las sabanas de los Llanos Orientales (Decaëns y Rossi, 2001; Jiménez et al., 2003; Jiménez y Rossi, 2006; Decaëns et al., 2009; Jiménez et al., 2012; Jiménez et al., 2014). Sin embargo, se desconoce qué sucede en otros ecosistemas y con grupos de organismos diferentes a las lombrices, donde el conocimiento de su dinámica espacial puede ser útil para entender procesos del ecosistema en los cuales participan activamente.
Por otra parte, la caficultura se estableció como una actividad socio-económica primordial para el desarrollo del país, especialmente en las últimas décadas del siglo XX, por lo que se han realizado diversos esfuerzos para mantener su competitividad en el mercado internacional. La búsqueda de tecnologías más eficientes con este fin ha permitido que se hayan desarrollado investigaciones en diferentes áreas (Cadena-Gómez, 2005), como la biotecnológica, la edafológica (Suárez de Castro y Rodríguez-Granda, 1962; Gómez-Aristizábal y Rivera-Posada, 1993; Ramírez-Builes et al., 2010), la climática (Camayo-Vélez et al., 2003), agronómica para el desarrollo de variedades resistentes a plagas y enfermedades con una mayor productividad (Castillo y Moreno, 1988; Alvarado-Alvarado et al., 2005; Castro-Caicedo y Cortina-Guerrero, 2009), el manejo de plagas y enfermedades (Bustillo, 1990), y la enfocada en biodiversidad (Salazar-Yepes et al., 2002; Botero et al., 2006; Sánchez-Clavijo et al., 2010), entre otras. Con todo, no han sido prácticamente explorados aspectos importantes para la definición de estrategias de manejo de potenciales plagas, como la distribución de artrópodos del suelo y factores asociados.
Este estudio se realizó en un cafetal, uno de los cultivos de mayor importancia económica para Colombia, con el fin de ampliar el conocimiento sobre cómo la intensificación en el uso del suelo puede afectar a nivel espacial a la edafofauna, Los objetivos fueron: 1) establecer si la presencia de un cafetal afecta la densidad, composición y estructura espacial de la meso- y macrofauna presente en la hojarasca; 2) conocer los atributos de la hojarasca y su patrón espacial, y 3) establecer si los cambios en la edafofauna estaban relacionados con los atributos de la hojarasca en la zona de estudio.
MATERIALES Y MÉTODOS
Para el estudio se seleccionó un cultivo de café (Coffea arabica L.) asociado con plátano (Musa sp.), de aproximadamente 1,5 ha, establecido desde hace 35 años (sistema común en la zona) y un área de bosque subandino secundario de aproximadamente 8 ha, con más de 50 años (uso de suelo de referencia; comunicación de propietarios de la finca), en el Municipio de Montenegro, cuenca del río la Vieja (Quindío, Colombia). El cultivo era fumigado con oxicloruro de cobre y Lorsban para el control de plagas, fertilizado con Triple 15 NPK, se realizaban zocas cada cuatro años y eliminación de plantas arvenses tres veces al año.
En la zona se presenta una humedad promedio anual de 79,5 %, una temperatura ambiental entre 18 y 24 °C, y precipitaciones entre 1000 a 3000 mm, con un régimen bimodal: dos épocas de lluvia de Marzo a Mayo, y de Septiembre a Noviembre (Díaz-Perilla et al., 1996). Los suelos se desarrollaron a partir de cenizas volcánicas y tienen las asociaciones cartográficas Alejandría-Isabela, Tebaida-Alejandría y consociación Armenia, con los componentes taxonómicos Typic Hapludalfs-Typic Hapludolls, Typics Distropepts-Typic Hapludalfs y Typic Hapludans (Díaz-Perilla et al., 2004; Alcaldía Municipal de Montenegro, 2006). El suelo del bosque se caracterizó como franco-arenoso (textura establecida con método de Bouyucos), con un pH (con potenciómetro de un 1: 1 v / v en agua destilada) entre 5,9 y 6,7, humedad (por el método gravimétrico, con una precisión de 0,01 g) entre 28 y 33 % y un contenido de materia orgánica (método de incineración) entre 7 y 14 %. El suelo del cafetal se caracterizó como franco-arenoso, con un pH entre 4,8 y 5,9, humedad entre 23 y 37 % y un contenido de materia orgánica entre 8 y 23 %.
Los dos usos de suelo, bosque y cafetal, se encuentran cada uno en una finca, "La Ilusión" y "La AlegrÍa", localizadas en 04º32'12,1" N-75º51'58,3" W, a 1171 m s.n.m. y 04º32'47,1"-75º49'49,8" W a 1208 m s.n.m., respectivamente. El muestreo fue realizado en Julio de 2006, la época de menor precipitación durante el año. En cada uso de suelo se trazó una cuadrícula sobre el suelo de 20 m x 20 m subdividida en 2,5 x 2,5 m, para un total de 81 puntos de muestreo, correspondientes a las intersecciones. En cada intersección se demarcaron dos cuadrados de 25 x 25 cm en los que se recolectó la hojarasca presente en bolsas plásticas con cierre hermético, que fueron almacenadas en neveras de poliestireno a 4 °C, para permitir su conservación. Para la extracción de la fauna en cada punto, una de las bolsas fue revisada manualmente y la otra fue colocada en embudos Berlese modificados o Tullgreen. Los individuos colectados (tanto en la revisión manual como en la muestra procesada por Berlese) se sumaron y se determinó la densidad de los individuos encontrados en cada muestra.
La identificación taxonómica se realizó hasta el menor nivel taxonómico posible con ayuda de claves (Krantz, 1978; Arnett et al., 1980; Borror et al., 1989; Naumann et al., 1990; Hoffman et al., 1996; Lawrence et al., 2000; Adis, 2002; Fernández y Sharkey, 2006; Krantz y Walter, 2009) y la asesoría de expertos en Aranae, Hymenoptera, Coleoptera y Acari. En la mayoría de los casos la determinación llegó hasta familia y en otros casos hasta orden, debido a la falta de material de referencia apropiado. Posteriormente, cada grupo taxonómico se clasificó en un grupo funcional de acuerdo a Lavelle (1996), teniendo en cuenta una revisión sobre la preferencia alimentaria más destacada. Para cada muestra se calculó la proporción de cada grupo funcional con respecto al total de individuos. Los ejemplares colectados están depositados en el Museo Javeriano de Historia Natural de la Pontificia Universidad Javeriana, Bogotá, Colombia.
La determinación del porcentaje de humedad de la hojarasca se estableció gravimétricamente, por secado a 80 ºC, durante 48 horas (Andrades-Rodríguez, 1996). Usando el dato de peso seco obtenido por este procedimiento se calculó también la biomasa de la hojarasca.
Para los análisis se seleccionaron solo los grupos con más de un individuo (en promedio/muestra) o con más de 50 individuos en total. La relación de las características de la hojarasca con las de la fauna se estableció mediante un test de Mantel con el programa zt (versión 1.1., Bonnet y Van de Peer, 2003). Se calcularon los coeficientes de varianza (CV) para la densidad de la fauna y de la humedad de la hojarasca y se realizó una prueba F para establecer las diferencias entre las varianzas. Para determinar las diferencias en la densidad y composición del ensamblaje de grupos funcionales de la fauna edáfica entre el cultivo de café y el bosque se realizó una prueba de U de Mann-Whitney y, se calculó el índice de Jaccard, respectivamente. Se usaron tablas de contingencia con un nivel de significancia de 95 % para establecer diferencias en la proporción de los grupos funcionales entre los dos usos del suelo.
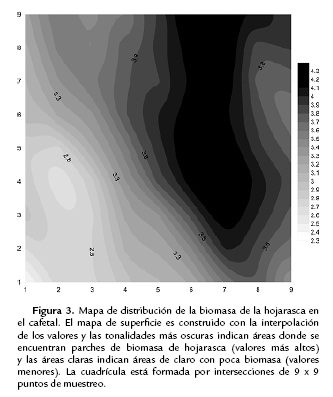
Para el análisis espacial de la humedad y biomasa de la hojarasca se utilizó geoestadística con el programa VarioWin (Pannatier, 1996) y SADIE (por sus siglas en ingles: Spatial Analysis by Distance IndicEs) mediante el programa SADIEshell (Conrad, 2001), para la abundancia de la fauna. Se calculó el índice de agrupación (Ia; basado en la distancia que tendrían que recorrer los individuos para estar uniformemente distribuidos) y la probabilidad, Pa (que los datos observados sean más agregados que lo que es esperado, por una permutación al azar con un α de 0,05). También se calculó el vi (índice de parche: adscrito a unidades o puntos de muestreo en los que el conteo de individuos es mayor a la media de las unidades) y el vj (índice de claro: adscrito a unidades o puntos de muestreo en los que el conteo de individuos es menor a la media de las unidades) de acuerdo con Perry (1998), Perry et al. (1999) y Perry y Dixon (2002). Posteriormente se realizaron mapas de distribución de la variable procesados en Surfer (versión 8.05, Golden software, Inc., Golden, U.S.A). Con el fin de establecer el tamaño de los parches, cada punto de intersección de la cuadrícula se tomó como un punto del área que conforma un parche o claro.
RESULTADOS
Densidad de la fauna de la hojarasca
Se encontró un total de 6682 individuos distribuidos en ocho clases: Clitellata, Gastropoda, Diplopoda, Chilopoda, Symphyla, Entognatha, Insecta (Ectognatha), y Arachnida (Tabla 1). Aunque la densidad promedio de la fauna de la hojarasca en el cafetal fue dos veces mayor (56,26 ± 10,72) a la del bosque (27,96 ± 3,24), no hubo diferencias significativas (U = 3 013, p = 0,82), probablemente debido a la gran variación intraespecífica, principalmente en el bosque, lo que sugiere una mayor heterogeneidad en este. También, considerando todos los grupos se encontró un poco más del doble de individuos en el cafetal (4557) que en el bosque (2125), lo que equivale a 900 individuos/m2 en cafetal, contrastando con aproximadamente 425 individuos/m2 en bosque.

Los taxones más abundantes fueron dos subórdenes de Acari (Oribatida, del orden Acariformes y, Gamasida perteneciente al orden Parasitiformes), un Orden completo de Entognatha (Collembola), el Orden Coleoptera (excluyendo las larvas), una familia del Orden Hymenoptera (Formicidae), las larvas de Diptera y la Clase Diplopoda. A diferencia de lo esperado el cultivo de café presentó los coeficientes de variación (CV; entre 180 y 300 % en cultivo y entre 130 y 220 en bosque) y las varianzas (entre 8 y 800 en cultivo y entre 0,22 y 109 en bosque) más altos para la mayoría de los grupos taxonómicos, excepto para Formicidae (S2 = 12,39 y CV = 459,71 en cultivo y S2 = 15,92 y CV = 383,99 en bosque).
Composición de la fauna de la hojarasca
La distribución de los grupos taxonómicos en el bosque y en el cultivo fue desigual en algunos casos. Para taxones con un grado de determinación menos específico como Oligochaeta, Gastropoda, Psocoptera, Actinedida, Gamasida (No Uropodidae) y Gamasida (Uropodidae), las diferencias dentro de cada una de las coberturas no fueron evidentes, mientras para grupos con un nivel de identificación más específica se observaron diferencias entre el bosque y el cafetal (Tabla 1). El total de grupos taxonómicos compartidos por los dos usos de suelo fueron 43, mientras que el número de grupos exclusivos de bosque y cafetal fueron 30 y 31, respectivamente.
Se observó una baja similitud en la composición faunística entre bosque y cafetal (menor a 40 %). Tanto en bosque como en el cafetal, la mayor densidad de individuos estuvo representada por Acari, encontrándose 66 % en bosque y 58 % en cultivo.
Después de Acari, el grupo mejor representado en el bosque fue Collembola (11 %); los grupos que siguieron en abundancia fueron Diptera (6 %), Hymenoptera (4 %) y Pseudoescorpionida (3 %). En el cafetal, el segundo grupo más abundante, pero con una mayor proporción que en el bosque con respecto a otros grupos, también fue Collembola (17 %); después de este, los grupos más abundantes en cultivo fueron Coleoptera (11 %) y Diptera con 5 % (Fig. 1).
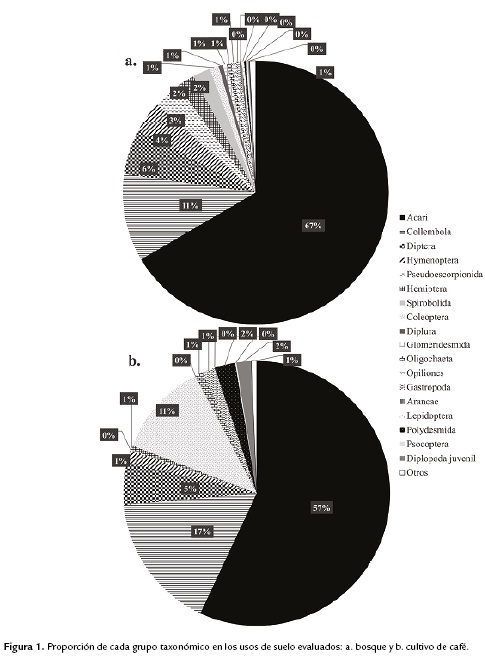
Se evidenció una tendencia de aumento en la abundancia de Collembola en el cultivo; la razón Acari/Collembola fue hasta 1,8 veces mayor en cultivo (3,4) que en el bosque (6,2).
Estructura espacial de la fauna de la hojarasca
Coleoptera y las larvas de Diptera presentaron una estructura espacial agregada, detectable y significativa, en el cafetal (Pa = 0,04 en los dos casos). Los demás taxones no mostraron ninguna estructura de agregación, como se observa en la Fig. 2.

Composición y densidad de los grupos funcionales
Sólo la proporción de los transformadores de hojarasca fue mayor en el cafetal que en el bosque (χ2 = 26,16, p < 0,05). La composición en términos de presencia/ausencia de los grupos funcionales de los micropredadores e ingenieros del ecosistema evidencia disimilaridad entre el cafetal y el bosque (41 y 18 %, respectivamente). Por otra parte, con base en el número de individuos de los grupos taxonómicos que conforman a los transformadores de hojarasca, se evidenció una alta similitud en la composición de la edafofauna entre bosque y cafetal (78 %). La relación entre la biomasa de la hojarasca y la densidad de los transformadores de hojarasca, fue baja y significativa únicamente en el bosque (R = 0,27, p < 0,05).
En la categoría de micropredadores se incluyeron los taxones en los cuales existe reporte en la literatura que se alimentan de otros invertebrados y microorganismos (Lavelle, 1996). En el bosque este grupo presentó 2207 individuos y estuvo conformado por Chilopoda, Collembola, Diplura (Japigidae), Protura, "Blattaria", algunos Coleoptera (Staphylinidae y Scydmaenidae), Araneae, Opiliones (Cosmetidae), Pseudoescorpionida y todos los Acari. En el cultivo este grupo presentó un total de 3497 individuos y estuvo conformado por Chilopoda, Collembola, Diplura (Japigidae), "Blattaria", algunos Coleoptera (Staphylinidae, Scydmaenidae, Ptillidae, Histeridae, Cucujidae y Corylophidae), Pseudoescorpionida, Araneae y todos los pertenecientes a Acari.
Los transformadores de hojarasca estuvieron conformados por los taxones conocidos por consumir materia orgánica y que al defecar crean un medio para los microorganismos (Lavelle, 1996). Este grupo presentó 201 individuos en bosque y 493 individuos en cultivo de café. En el bosque los transformadores de hojarasca estuvieron conformados por Gastropoda, Diplopoda, larvas de Diptera, Psocoptera y algunos Coleoptera (Nitidulidae, Ptilodactylidae e Hydrophilidae). En el cultivo esta categoría estuvo conformada también por Gastropoda, Diplopoda, larvas de Diptera y Psocoptera; sin embargo las familias de Coleoptera presentes fueron además de Nitidulidae y Ptilodactylidae, presentes también en el bosque, Leiodidae, Aphodiidae y Scarabaeidae.
Los ingenieros del ecosistema son un grupo que cambia la estructura local y la composición del suelo. En este estudio los ingenieros del ecosistema estuvieron conformados únicamente por Oligochaeta y Formicidae, de acuerdo con la agrupación realizada por Jones et al. (1994). En esta categoría funcional se encontraron un total de 90 individuos en bosque y de 108 individuos en cultivo de café.
Estructura espacial de los atributos de la hojarasca y su relación con la fauna edáfica
De la evaluación entre variables abióticas y grupos de fauna se destaca la relación positiva entre la humedad de la hojarasca (entre 40 y 85 % de humedad, en promedio 61 ± 9 %) y la densidad de Formicidae en el cafetal (R = 0,38, p = 0,03) y, la biomasa de la hojarasca en bosque (R = 0,41, p = 0,005). Las densidades de los demás grupos de fauna con mayor número de individuos no mostraron relación alguna con las variables abióticas medidas de la hojarasca.
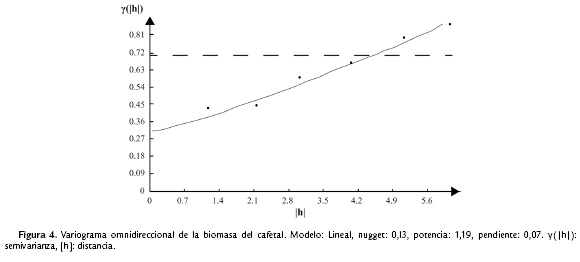
En cuanto a las variables abióticas las distancias evaluadas en el estudio no mostraron dependencia entre los puntos de muestreo (es decir un efecto de nugget puro, el cual representa ausencia total de correlación entre los puntos por la variación a distancias mucho menores que el intervalo de muestreo más pequeño), a excepción de la biomasa de la hojarasca con el cultivo (Fig. 3).
La variación de la biomasa de hojarasca en el cultivo (entre 4,16 y 128,79 g) fue de tipo anisotrópico. El modelo lineal que se ajustó para el variograma omnidireccional (en conjunto para todas las direcciones) indica que la autocorrelación entre puntos no se estabiliza y por lo tanto no se observa la distancia a la que los puntos evaluados en el área empiezan a ser independientes entre sí (Fig. 4). Por esta razón no se pudo establecer el tamaño de los parches.
Se encontró un parche para la biomasa de la hojarasca en el cultivo, centrado en (x = 7, y = 4; considerando el origen en la esquina inferior izquierda, punto x = 0 y y = 0) que coincide con parches encontrados para Diplopoda y Gamasida (No Uropodidae). Sin embargo, esta coincidencia no fue significativa para ninguno de los grupos.
DISCUSIÓN
Densidad de la fauna de la hojarasca
La densidad encontrada fue entre 1,5 y diez veces menor en este estudio comparado con otros estudios realizados en sabana nativa, bosque subandino, selva de galería y en un transecto bosque-pastizal en México (Benito et al., 2004; Feijoo y Knapp, 1998; Decaêns et al., 2003; Feijoo et al., 2005; Rueda et al., 2011). Estas diferencias en la densidad de la fauna encontrada pudieron deberse al método de extracción, el cual se centra principalmente en la mesofauna y deja de incluir a muchos grupos, como algunos tipos de ácaros (André et al., 2002; Coleman y Whitman, 2005).
También, se considera que influyeron el número de muestreos, la edad del bosque y su historia de intervención. Aunque se realizó un sólo muestreo, este tipo de diseños implican un gran esfuerzo tanto en campo como en laboratorio y muchos de los estudios que se realizan con éste, no tienen repeticiones (Görres et al., 1998; Jiménez et al., 2001; Chust et al., 2003; Negrete-Yankelevich, 2004; Jiménez y Rossi, 2006; Negrete-Yankelevich et al., 2006; Rossi et al., 2006; Jiménez et al., 2012). Por otra parte al comparar la densidad encontrada con la de estudios realizados en bosques de montaña de edad similar, las diferencias siguen siendo altas (Feijoo y Knapp, 1998; Negrete-Yankelevich, 2004), por lo que la época en la que se realizó el muestreo (de baja precipitación) pudo contribuir a que las condiciones no fueran las adecuadas para la colonización de la hojarasca por parte de la fauna, tal como lo reportó Ilieva-Makulec et al., (2006).
Estudios realizados en Colombia y en otros países han registrado que algunas prácticas agrícolas, como la labranza y el uso de productos de síntesis química, afectan negativamente la biomasa y diversidad de la macrofauna y la densidad de ácaros edáficos, respectivamente (Marín et al., 2002; Bedano et al., 2006). Sin embargo en el presente estudio se registró una abundancia dos veces más alta en el cultivo que en el bosque, probablemente por la presencia de sombrío y la poca remoción de material vegetal de la superficie del suelo, en el cafetal. Esto sugiere que características propias del sistema son importantes para la presencia y estructura de la edafofauna. Esta abundancia más alta en el cultivo se observó en puntos específicos, con picos de abundancia de ciertos grupos (larvas de Coleoptera y Staphylinidae adultos), posiblemente como consecuencia de condiciones climáticas particulares, como una mayor cantidad de lluvia en cultivo días antes del muestreo (10,5 mm y 25,3 mm en el bosque y en el cultivo, respectivamente, [CRQ, 2006]), una mayor densidad vegetal en el bosque, que hace que la cantidad de agua de lluvia que llega al suelo sea menor, y el uso de insecticidas como clorpirifos, al cual se le han atribuido efectos negativos sobre algunos invertebrados del suelo (Jânsch et al., 2006; Fountain et al., 2007), favoreciendo a otros. Un resultado similar al de este estudio fue observado por Ettema y Yates (2003) en Nueva Zelanda, con una mayor abundancia de artrópodos en el pastizal en relación con un bosque de latifoliadas. Sin embargo vale mencionar que hubo un mayor número de géneros y de intercambio genético en el bosque tanto a pequeña y mediana escala. Valdría la pena evaluar si en cultivos perennes como el del presente estudio este patrón sería similar.
Composición de la fauna de la hojarasca
El orden Collembola se conoce como tolerante a la aplicación de insecticidas y a contaminación de distintos tipos, como la presencia de metales pesados (Greenslade, 1990), lo que explicaría el aumento en su proporción con respecto a los otros grupos taxonómicos en el cafetal de este estudio, coincidiendo con reportes anteriores que indican que la razón Acari/Collembola es menor en sistemas perturbados, por el aumento de estos últimos (Rodríguez, 1998; Rueda et al., 2011). Otras condiciones que pueden afectar su abundancia y capacidad reproductiva son el contenido de materia orgánica (Eaton et al., 2004), humedad y luminosidad, entre otras (Butcher et al., 1971), por lo que estos factores deben también tenerse en cuenta para establecer su efecto en bajo las condiciones de los usos de suelo estudiados.
Estructura espacial de la fauna de la hojarasca
Pocos estudios sobre la distribución espacial de Coleoptera están disponibles para comparar con nuestros resultados. Se han encontrado patrones espaciales de distribución detectables por debajo de 0,5 m y hasta 150 m, en distintos sistemas con algún grado de intervención y ciertos grupos de artrópodos (Frouz y Paoletti, 2000; Holland et al., 2007; Nansen et al., 2004; Negrete-Yankelevich, 2004; Rueda et al., 2011; De la Rosa y Negrete-Yankelevich, 2012; Jimenez et al., 2014), que no involucran cultivos. En este estudio Coleoptera y Diptera mostraron un patrón detectable a un espaciamiento de 2,5 m en el cultivo de café asociado con plátano. Lo anterior reafirma la existencia de diferentes escalas en las que se distribuye la fauna, dependiendo de factores como el contenido de materia orgánica, la cobertura vegetal y la altura de la vegetación, con los cuales algunos estudios anteriores encontraron correlación positiva. Con esta base y los resultados del presente estudio, es claro que la distribución espacial de los organismos no se puede generalizar para hábitats más intervenidos y menos intervenidos, debido a que los taxa se ven afectados diferencialmente, por su sensibilidad diferencial a las perturbaciones. Por tanto, se sugiere usar ciertos grupos indicadores como Collembola o algunos transformadores de hojarasca, con el fin de identificar diferencias en el grado o en el tipo de perturbación.
Por otro lado, la detección del patrón espacial de estos dos taxones únicamente en cultivo, lleva a plantear que en el bosque la estructura espacial es más fina, es decir, con parches más pequeños que no pudieron ser detectados por las distancias de muestreo utilizadas. El hecho de que el bosque presente una mayor heterogeneidad en la vegetación y en los recursos que aporta la misma al suelo permite una agregación espacial mayor de taxones, como lo reportan De la Rosa y Negrete-Yankelevich (2012) y Jiménez et al. (2012), al posibilitar la coexistencia de especies potencialmente competidoras y facilitar la agrupación en espacios más pequeños dentro de áreas con propiedades de suelo similares.
La pequeña escala espacial utilizada en este estudio es insuficiente para profundizar en los patrones en los que se distribuye la fauna edáfica y, por tanto, se destaca la necesidad de ampliar los análisis a una perspectiva multi-escala como lo proponen Jiménez et al. (2012). Seleccionar a priori una distancia apropiada entre puntos para el muestreo de la fauna del suelo es un asunto complejo (Decaëns y Rossi, 2001; Jiménez et al., 2001; Jiménez y Rossi 2006), por lo que es necesario realizar investigaciones adicionales con diferentes taxones y ecosistemas, para ampliar el conocimiento y poder ofrecer estrategias tanto para optimizar los muestreos como para generar un mejor manejo de los suelos.
Composición y densidad de los grupos funcionales
La mayor proporción de los transformadores de hojarasca en el cafetal posiblemente se debe a los grandes aportes de la cobertura de sombrío (hojas de plátano), sitios en donde la hojarasca alcanza espesores mayores (aproximadamente hasta 10 cm). Sin embargo, la relación entre la biomasa de la hojarasca y la densidad de los transformadores de hojarasca (conformados como taxones como Gastropoda, Diplopoda y larvas de Diptera) sólo fue significativa en el bosque, probablemente como consecuencia de que estos organismos se relacionan tanto con características de la composición vegetal como la calidad de ésta (Swift et al., 1979; Lavelle, 1997; Lavelle y Spain, 2001), la cual es más variada en el bosque debido a la alta riqueza de plantas. Por tanto el grupo de transformadores de hojarasca puede convertirse en un indicador de perturbación, dándole relevancia a la clasificación funcional.
La disimilaridad entre los usos de suelo en la composición de micropredadores e ingenieros del ecosistema se puede atribuir al tipo de hojas, su grosor, composición química y espesor de la hojarasca que, por ser características de la fuente alimenticia, afectan la distribución y composición (Lavelle, 1997; Lavelle y Spain, 2001), o, a aspectos metodológicos como la realización de un sólo muestreo. De igual manera el nivel de determinación de los grupos taxonómicos puede influir al cambiar el número de taxones usados en la comparación.
La alta similitud entre el bosque y el cafetal en la composición de los transformadores de hojarasca puede significar que esta parece no marcar diferencias importantes entre los dos usos del suelo. Más bien pueden ser características de la vegetación o del suelo, las que afectan directamente a los micropredadores y a los ingenieros del ecosistema. Por lo anterior, se debería hacer una evaluación de los grupos funcionales en capas superficiales del suelo, especialmente en los primeros 5 cm, donde hay mayor cantidad de materia orgánica y en los que se puede encontrar la mayor concentración de la fauna (Iglesias-Briones et al., 2009; Silva, 2002).
En el presente trabajo se resalta la importancia de la escala en la determinación de la distribución espacial de la fauna edáfica, así como de la respuesta distintiva de los grupos de organismos a modificaciones en el ambiente. De hecho, el cultivo afectó diferencialmente la distribución espacial de algunos de los taxones de los artrópodos de la hojarasca, como Coleoptera y las larvas de Diptera, lo que se manifestó en la variación en su distribución. Es importante detectar, en el ámbito espacial, este tipo de cambios fácilmente reconocibles, relacionados con características del suelo, de la hojarasca o de la vegetación, porque esto permitiría manejar los cultivos y las plagas de manera diferencial. Así, se le da énfasis a los sitios del cultivo que tienen mayor densidad de insectos que pueden potencialmente convertirse en plaga y prioridad a los focos que permiten la proliferación o colonización de grupos de importancia económica, como especies fitófagas que están dentro de grupos encontrados únicamente en cultivo (por ejemplo: Thripidae, algunas larvas, Scutigerellidae). De manera adicional, el conocimiento de la dinámica espacial de los grupos del suelo que están involucrados en procesos importantes en este, como en el ciclo de nutrientes, puede ser de gran utilidad para un manejo eficiente de los recursos, ya que se conseguiría un aprovechamiento de las funciones que estos organismos realizan naturalmente en los ecosistemas, como es el caso de Collembola y de varios grupos de Acari.
Estructura espacial de los atributos de la hojarasca y su relación con la fauna edáfica
La relación entre la humedad y la densidad de Formicidae fue reportada anteriormente por Levings y Windsor (1982). Esta relación encontrada únicamente en el cultivo, coincide con variaciones marcadas en la humedad de la hojarasca en ciertos puntos del área. La relación de la humedad con la biomasa de la hojarasca sugiere que la alta variabilidad en la hojarasca puede crear mayor cantidad de sitios donde se encuentra agua. Sin embargo, es necesario evaluar otras características como el pH, la temperatura y la calidad de la hojarasca, entre otras, que puedan explicar mejor esa relación.
La detección de un patrón espacial de la biomasa únicamente en el cultivo apoya también la hipótesis de una escala más pequeña para el caso de la distribución de la biomasa de la hojarasca en bosque, la cual puede estar determinada por la distribución de la vegetación.
Los parches de la biomasa de la hojarasca que coinciden con los parches de Diplopoda y Gamasida pueden ser explicados por el consumo de material vegetal (Bueno, 2003), que haría que se concentraran en estos sitios, al igual que factores no evaluados como el grado de descomposición y el contenido de calcio y metabolitos secundarios, como lo reporta Levings y Windsor (1982).
CONCLUSIONES
El cambio en el uso del suelo de bosque a cafetal fue evidente en la composición faunística y en la distribución espacial de algunos grupos, mas no en la densidad. En los patrones espaciales se determinó un aumento en los tamaños de los parches para Coleoptera, larvas de Diptera y biomasa de la hojarasca, con un patrón de distribución agregado para estos dos grupos de insectos. Se sugiere entonces la realización de un estudio espacio-temporal en estos usos del suelo con el fin de evaluar si hay cambios de los patrones en el tiempo y cómo son estos cambios. Este trabajo permite establecer la trascendencia que tiene la escala en la determinación de la distribución espacial de la fauna edáfica, así como de la respuesta diferencial de los grupos de organismos a modificaciones en el ambiente. Es importante detectar cambios en el ámbito espacial relacionados con características de suelo, de la hojarasca o de la vegetación fácilmente reconocibles, porque esto permitiría manejar los cultivos y las plagas de manera diferencial, dándole énfasis a los sitios del cultivo que tienen mayor densidad de insectos que pueden ser potenciales plagas, y dando prioridad a los focos que permiten la proliferación o colonización de grupos de importancia económica.
AGRADECIMIENTOS
Agradecemos al Centro de Investigaciones en Biodiversidad y Recursos Genéticos (CIEBREG) por el apoyo financiero, dentro del proyecto: "Valoración de los bienes y servicios de la biodiversidad para el desarrollo sostenible de paisajes rurales colombianos: Complejo ecorregional de los Andes del Norte", y en particular a la Pontificia Universidad Javeriana, como parte de este Centro de Excelencia (ID 00000330). Adicionalmente a Juan José Jiménez por los comentarios y consejos en el proyecto inicial, así como a todas las personas que nos apoyaron en la identificación de los grupos taxonómicos: Alexander Sabogal (Universidad Nacional de Colombia), Fernando Fernández (Universidad Nacional de Colombia), Luis Gabriel Pérez (Pontificia Universidad Javeriana), Germán Amat (Universidad Nacional de Colombia), Eric Manzano (Pontificia Universidad Javeriana), Jorge Noriega (Pontificia Universidad Javeriana) y Orlando Cómbita (The Ohio State University).
REFERENCIAS
Adis J. Amazonian Arachnida and Myriapoda. Identification keys to all classes, orders, families, some genera, and lists of known terrestrial species. Sofia: Pensoft publishers; 2002. 590 p. [ Links ]
Alonso-Zarazaga MA, Lyal CHC. A catalogue of family and genus names in Scolytinae and Platypodinae with nomenclatural remarks (Coleoptera: Curculionidae). Zootaxa. 2009;2258:1-134. [ Links ]
Andrades-Rodríguez M. Prácticas de edafología y climatología. Logroño: Servicio de publicaciones Universidad de la Rioja; 1996. 69 p. [ Links ]
André HM, Ducarme X, Lebrun P. Soil biodiversity: myth, reality or conning? Oikos. 2002;96(1):3-24. [ Links ]
Alcaldía Municipal De Montenegro-Quindío. Plan básico de ordenamiento territorial 2000-2006, Diagnostico Municipal. Montenegro: Municipio de Montenegro, Secretaria de Planeación Municipal; 2006. 130 p. [ Links ]
Alvarado-Alvarado G, Posada-Suárez HE; Cortina-Guerrero HA. CASTILLO: Nueva variedad de café con resistencia a la roya. Avances Tec Cenicafé. 2005;337:1-8. [ Links ]
Arnett RH, Downie NM, Jacques HE. How to know the beetles. 2 ed. New York: WCB McGraw-Hill; 1980. 500 p. [ Links ]
Bedano JC, Cantú MP, Doucet ME. Influence of three different land management practices on soil mite (Arachnida: Acari) densities in relation to a natural soil. App Soil Ecol. 2006:32(3):293-304. [ Links ]
Benito NP, Brossard M, Pasini A, Guimarâes MF, Bobillier B. Transformation of soil macroinvertebrates populations alter native vegetation conversion to pasture cultivation (Brazilian Cerrado). Eur J Soil Biol. 2004;40(3-4):147-154. [ Links ]
Bonnet E, Van De Peer Y. ZT: a software tool for simple and partial Mantel tests [Internet]. Ghent: Ghent University; 2003. 12 p. [Cited 2014 May 25] Available from: http://www.jstatsoft.org/v07/i10/paper. [ Links ]
Borror D, Triplehorn CA, Johnson NF. An introduction to the study of insects. San Diego: Editorial Saunders College; 1989. 864 p. [ Links ]
Botero JE, Arbeláez D, Lentijo G, Castellanos O, Franco N. Herramientas de la conservación de la Biodiversidad. Biocarta. 2006;9:1-4. [ Links ]
Bueno J. Los diplópodos del suelo en la selva alta de los Tuxtlas. In: Álvarez-Sánchez J, Naranjo-García E, editors. Ecología del suelo en la selva tropical húmeda de México. Xalapa: Universidad Nacional Autónoma de México, Instituto de Biología, Facultad de Ciencias, Instituto de Ecología, A.C.; 2003. p. 226-236. [ Links ]
Bustillo PA. Perspectivas de manejo integrado de la broca del café Hypothenemus hampei en Colombia. In: Seminario sobre la broca del café. Medellín: Socolen; 1990. p. 91-105. [ Links ]
Butcher JW, Snider R, Snider RJ. Bioecology and of edaphic collembola acarina. Annu Rev Entomol. 1971:16:249-288. [ Links ]
Brusca RC, Brusca GJ. Invertebrates. 2 ed. Sundeland: Sinauer Associates, Inc; 2003. 936 p. [ Links ]
Cadena-Gomez G. Desarrollos científicos de Cenicafé en la última década. Rev Acad Colomb Cienc. 2005;29(110):89-99. [ Links ]
Camayo-Vélez GC, Chaves-Córdoba B, Arcila-Pulgarín J, Jaramillo-Robledo A. Desarrollo floral del cafeto y su relación con las condiciones climáticas de Chinchiná-Caldas. Cenicafé. 2003;54(1):35-49. [ Links ]
Castillo ZJ, Moreno R. La variedad Colombia: Selección de un cultivar compuesto resistente a la roya del cafeto. Bogotá: Editolaser; 1988. 171 p. [ Links ]
Castro-Caicedo BL, Cortina-Guerrero HA. Evaluación de resistencia a Ceratocystis fimbriata Ell Halsdt. Hunt. en progenies F5 de café Brobón resistenta x Caturra. Cenicafé. 2009;60(2):115-125. [ Links ]
Chust G, Pretus JL, Ducrot D, Bedos A, Deharveng L. Response of soil fauna to landscape heterogeneity: Determining optimal scales for biodiversity modelling. Conserv Biol. 2003;17(6):1712-1723. Doi:10.1111/j.1523-1739.2003.00564.x. [ Links ]
Coleman DC, Whitman WB. Linking species richness, biodiversity and ecosystem function in soil systems. Pedobiologia. 2005;49(6):479-497. [ Links ]
Conrad. SADIEshell. Graphical User Interface (GUI). Versión 1.22; 2001. [ Links ]
Corporación Autónoma Regional del Quindío (CRQ). Valores Totales Diarios de Precipitación (mm) [Internet]. Montenegro, Quindío, Colombia: Estación 470001628 VIVERO MONTENEGRO. Subdirección de Control y Seguimiento Ambiental-Subdirección de Ejecución de Políticas Ambientales, Equipo de Aguas; 2006. [Cited 2014 May 25] Available from: http://www.crq.gov.co/PRECIPITACION2006.mht. [ Links ]
Crist TO. The spatial distribution of termites in shortgrass steppe: a geostatistical approach. Oecologia. 1998;114(3):410-416. [ Links ]
Decaëns T, Rossi JP. Spatio temporal structure of earthworm community in a tropical pasture. Ecography. 2001;24(6):671-682. [ Links ]
Decaëns T, Lavelle P, Jiménez JJ, Escobar G, Rippstein G, Schneidmadl J, et al. El impacto del uso de la tierra en la macrofauna del suelo de los Llanos Orientales de Colombia. In: Jiménez JJ, Thomas RJ, editors. El arado natural: La comunidades de macroinvertebrados del suelo en las sabanas Neotropicales de Colombia. Cali: Centro Internacional de Agricultura Tropical (CIAT); 2003. p. 21-45. [ Links ]
Decaëns T, Jimenez JJ, Rossi JP. A null-model analysis of the spatiotemporal distribution of earthworm species assemblages in Colombian grasslands. J Trop Ecol. 2009;25(4):415-427. [ Links ]
Deyrup M. A New Species of Flightless Pygmy Mole Cricket from a Florida Sand Ridge (Orthoptera: Tridactylidae). Fla Entomol. 2005;88(2):141-145. [ Links ]
De La Rosa IN, Negrete-Yankelevich S. Distribución espacial de la macrofauna edáfica en bosque mesófilo, bosque secundario y pastizal en la reserva La Cortadura, Coatepec, Veracruz, México. Rev Mex Biodivers. 2012;83(1):201-215. [ Links ]
Díaz-Perilla PA, Roveda G, Guevara GA. Suelos del Departamento del Quindío. Bogotá: Instituto Geográfico Agustín Codazzi (IGAC); 1996. 205 p. [ Links ]
Díaz-Perilla PA, Alvarado-Mora PD, Poveda-Hoyos G. Estudio general de suelos y zonificación de tierras [Recurso electrónico]. 2ª edición. Bogotá: Instituto Geográfico Agustín Codazzi; 2004. [ Links ]
Eaton RJ, Barbercheck M, Buford M, Smith W. Effects of organic matter removal, soil compaction, and vegetation control on Collembolan populations. Pedobiologia. 2004:48(2):121-128. [ Links ]
Edwards CA. Bionomics of symphyla (Tesis de doctorado). Madison, WI: University of Wisconsin; 1957. 165 p. [ Links ]
Ettema CH, Wardle DA. Spatial soil ecology. Trends Ecol Evol. 2002;17(4):177-183. [ Links ]
Ettema CH, Yeates GW. Nested spatial biodiversity patterns of nematode genera in a New Zealand forest and pasture soil. Soil Biol Biochem. 2003;35(2):339-342. [ Links ]
Feijoo A, Knapp EB. El papel de los macroinvertebrados como indicadores de fertilidad y perturbación de suelos de laderas. Suelos Ecuat. 1998;28:254-259. [ Links ]
Feijoo A, Zuñiga MC, Camargo C. Signs to detect regeneration and degradation of agroecosystems in the coffee growing region of Colombia [Internet]. Livest Res Rural Dev. 2005;17(3):[2 pantallas]. [Cited 2009 Jul 31] Available from: URL: http://www.lrrd.org/lrrd17/3/feij17025.htm. [ Links ]
Fernández F, Sharkey MJ, editors. Introducción a los Hymenoptera de la región Neotropical. Bogotá: Sociedad Colombiana de Entomología y Universidad Nacional de Colombia; 2006. 894 p. [ Links ]
Fountain MT, Brown VK, Gange AC, Symondson WOC, Murria PJ. The effects of the insecticide chlorpyrifos on spider and Collembola communities. Pedobiologia. 2007;51(2):147-158. [ Links ]
Frouz J, Paoletti MG. Spatial distribution of different life stages of one Dipteran community along hedgerow and field margin. Landsc Urban Plan. 2000(1-2);48:19-29. [ Links ]
Futuyma DJ. Food plant specialization and environmental predictability in Lepidoptera. Am Nat. 1976;110(972):285-292. [ Links ]
Gómez-Aristizábal A, Rivera-Posada H. La conservación de los suelos y la sostenibilidad de la productividad en la zona cafetera. Avances Técnicos Cenicafé. 1993;190:1-8. [ Links ]
Görres JH, Dichiaro MJ, Lyons JB, Amador JA. Spatial and temporal patterns of soil biological activity in a forest and an old field. Soil Biol Biochem. 1998;30(2):219-230. [ Links ]
Greenslade PJ. Collembola. In: Naumann ID, Carne PB, Lawrence JF, Nielsen ES, Spradbery JP, Taylor RW, et al, editors. The Insects of Australia. Volume I. New York: Cornell University Press; 1990. p. 252-264. [ Links ]
Grimaldi D, Engel MS. Evolution of the Insects. New York: Cambridge Universitu Press; 2005. 755 p. [ Links ]
Hoffman RL, Golovatch SI, Adis J, Morais JW. Practical keys to the orders and families of millipedes of the Neotropical region (Myriapoda: Diplopoda). Amazoniana. 1996;XIV(1/2):1-35. [ Links ]
Holland JM, Thomas CFG, Birkett T, Southway S. Spatio-temporal distribution and emergence of beetles in arable fields in relation to soil moisture. Bull Entomol Res. 2007;97(1):89-100. [ Links ]
Iglesias-Briones MJ, Ostle NJ, Mcnamara NP, Poskitt J. Functional shifts of grassland soil communities in response to soil warming. Soil Biol Biochem. 2009;41(2):315-322. [ Links ]
Ilieva-Makulec K, Olejniczak I, Szanser M. Response of soil micro- and mesofauna to diversity and quality of plant litter. Eur J Soil Biol. 2006;42(Sup 1):S244-S249. [ Links ]
Jânsch S, Frampton GK, Rômbke J, Van Den Brink PJ, Scott-Fordsmand JJ. Effects of pesticides on soil invertebrates in model ecosystem and field studies: a review and comparison with laboratory toxicity data. Environ Toxicol Chem. 2006;25(9):2490-2501. [ Links ]
Jiménez JJ, Rossi J-P. Spatial dissociation between two endogeic earthworms in the Colombian "Llanos". Eur J Soil Biol. 2006;42(Sup 1):S218-S224. [ Links ]
Jiménez JJ, Rossi J-P, Lavelle P. Spatial distribution of earthworms in acid-soil savannas of the Eastern plains of Colombia. Appl Soil Ecol. 2001;17(3):267-278. [ Links ]
Jiménez JJ, Rossi J-P, Lavelle P. Distribución espacial de las lombrices en las sabanas de los suelos ácidos de los llanos orientales. In: Jiménez JJ, Thomas RJ, editors. El arado natural: La comunidades de macroinvertebrados del suelo en las sabanas Neotropicales de Colombia. Cali: Centro Internacional de Agricultura Tropical (CIAT); 2003. p. 115-134. [ Links ]
Jiménez JJ, Decaëns T, Rossi J-P. Soil environmental heterogeneity allows spatial co-occurrence of competitor earthworm species in a gallery forest of the Colombian 'Llanos'. Oikos. 2012:121(6):915-926. [ Links ]
Jiménez JJ, Decaëns T, Lavelle P, Rossi J-P. Dissecting the multi-scale spatial relationship of earthworm assemblages with soil environmental variability. BMC Ecology. 2014:14(26):1-19. Doi: 10.1186/s12898-014-0026-4. [ Links ]
Jones CG, Lawton JH, Shachak M. Organisms as ecosystem engineers. Oikos. 1994;69:373-386. [ Links ]
Jung S, Lee S. Molecular phylogeny of the plant bugs (Heteroptera: Miridae) and the evolution of feeding habits. Cladistics. 2012;28(1):50-79. [ Links ]
Kondo T, Gullan PJ, Williams DJ. Coccidology. The study of scale insects (Hemiptera: Sternorrhyncha: Coccoidea). Revista Corpoica-Ciencia y Tecnología Agropecuaria. 2008;9(2):55-61. [ Links ]
Krantz GW, editor. A manual of acarology. 2 ed. Inc. Corvallis: Oregon state University Book Stores; 1978. 508 p. [ Links ]
Krantz GW, Walter DE, editors. A Manual of Acarology. 3 ed. Lubbock: Texas Tech University Press; 2009. 704 p. [ Links ]
Lavelle, P. Diversity of soil fauna and ecosystem function. Biol Int. 1996;33:3-16. [ Links ]
Lavelle P. Faunal activities and soil Processes: Adaptative strategies that determine ecosystem function. Adv Ecol Res. 1997;27:93-132. [ Links ]
Lavelle P, Spain A. Soil ecology. Norwell: Kluwee academic publishers; 2001. 654 p. [ Links ]
Lawrence JE, Hastings AM, Dallwitz MJ, Paine TA, Zurcher EJ. Beetles of the world [Versión electrónica]. Version 1.0 for Microsoft Windows. Canberra: CSIRO Entomology; 2000. [ Links ]
Levings SC, Windsor DM. Seasonal and annual variation in litter arthropod populations. In: Leigh EG, Rand AS, Windsor DM, editors. The ecology of a Tropical Forest: Seasonal changes and long terms Rhythms. Washington, D.C.: Smithsonian Institution Press; 1982. p. 355-388. [ Links ]
Marín EP, Feijoo A, Peña J. Cuantificación de la macrofauna en un vertisol bajo diferentes sistemas de manejo en el Valle del Cauca. Suelos Ecuat. 2002;31(2):146-151. [ Links ]
Matson PA, Parton WJ, Power AG, Swift MJ. Agricultural Intensification and Ecosystem Properties. Science. 1997;277(5325):504-509. [ Links ]
Nansen C, Subramanyam B, Roesli R. Characterizing spatial distribution of trap captures of beetles in retail pet stores using SADIE® software. J Stored Prod Res. 2004;40(5):471-483. [ Links ]
Naumann ID, Carne PB, Lawrence JF, Nielsen ES, Spradbery JP, Taylor RW, et al, editors. The insects of Australia. Volume I-II. Ithaca Cornell University Press; 1990. 1137 p. [ Links ]
Negrete-Yankelevich S. Integrating soil macroinvertebrate diversity, litter decomposition and secondary succession in a tropical montane cloud forest in Mexico (Tesis de doctorado). Edinburg: University of Edinburg; 2004. 345 p. [ Links ]
Negrete-Yankelevich S, Fragoso C, Newton AC, Russell G, Heal OW. Spatial patchiness of litter, nutrients and macroinvertebrates during secondary succession in a Tropical Montane Cloud Forest in Mexico. Plant Soil. 2006;286(1-2):123-139. [ Links ]
Pannatier Y. VarioWin (Software for Spatial Data Analysis in 2D). Version 2.21. New York: Springer-Verlag; 1996. [ Links ]
Perry JN. Measures of spatial pattern for counts. Ecology. 1998;79(3):1008-1017. [ Links ]
Perry JN, Dixon P. A new method for measuring spatial association in ecological count data. Ecoscience. 2002;9(2):133-141. [ Links ]
Perry JN, Winder L, Holland JM, Alston RD. Red-blue plots for detecting clusters in count data. Ecol Lett. 1999;2(2):106-113. [ Links ]
Ramírez-Builes VH, Jaramillo-Robledo A, Arcila-Pulgarín J, Montoya-Restrepo EC. Estimación de la humedad del suelo en cafetales a libre exposición solar. Cenicafé. 2010;61(3):251-259. [ Links ]
Reitz SR, Yu-Lin G, Zhong-Ren L. Thrips: Pests of Concern to China and the United States. Agric. Sci. China. 2011;10(6):867-892. [ Links ]
Rodríguez MC. Evaluación y diagnóstico de la macrofauna y mesofauna edáficas de los suelos restaurados por industrias APASCO en cerro Buenavista (Tesis de maestría). Xalapa: Instituto de Ecología, A.C.; 1998. 136 p. [ Links ]
Rossi JP, Huerta E, Fragoso C, Lavelle P. Soil properties inside earthworm patches and gaps in a tropical grassland (La Mancha, Veracruz, Mexico). Eur J Soil Biol. 2006;42(Supplement 1):S284-S288. [ Links ]
Rueda DM, Negrete-Yankelevich S, Fragoso C. Escala de independencia espacial de la mesofauna edáfica en un transecto bosque-pastizal del Jardín Botánico "Francisco Javier Clavijero". Acta Zool Mex (nueva serie). 2011;27(1):191-195. [ Links ]
Salazar-Yepes M, Buriticá-C P, Cadena-Gómez G. Implicaciones de los estudios de biodiversidad de los Uredinales (royas) en la región cafetera colombiana. Cenicafé. 2002;53(3):219-238. [ Links ]
Sánches-Clavijo LM, Vélez JG, Durán SM, García R, Botero JE. Estudio Regional de la biodiversidad en los paisajes cafeteros de Támesis, Antioquia. Boletín Técnico Cenicafé. 2010;35:1-72. [ Links ]
Silva ES. Ácaros (Arthropoda: Acari) edáficos da Mata Atlântica e cerrado do Estado de São Paulo, com ênfase na superfamilia Rhodacaroidea (Tesis de Mestrado). Piracicaba: Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo; 2002. 100 p. [ Links ]
Suárez De Castro F, Rodríguez-Granda A. Investigaciones sobre la erosión y la conservación de los suelos en Colombia. Federación Nacional de Cafeteros de Colombia. Bogotá: Antares, Ltda.; 1962. 473 p. [ Links ]
Swift MJ, Heal OW, Anderson JM. Decomposition in terrestrial ecosystems. Studies in ecology, vol. 5. Oxford: Blackwell; 1979. 372 p. [ Links ]
Tashiro H. Insecta: Coleoptera. Scarabaeidae Larvae. In: Dindal DL, editor. Soil Biology Guide. USA: A Wiley-Interscience Publication. p. 583-666. [ Links ]
Thiele-Bruhn S, Bloem J, De Vries FT, Kalbitz K, Wagg C. Linking soil biodiversity and agricultural soil management. Curr Opin Environ Sustain. 2012;4(5):523-528. [ Links ]
Tilgner EH. Systematics of Phasmida (Tesis de doctorado). Athens: University of Georgia; 2002. 113 p. [ Links ]

