










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkActa Biológica Colombiana
versión impresa ISSN 0120-548X
Acta biol.Colomb. vol.21 no.3 Bogotá set./dic, 2016
https://doi.org/10.15446/abc.v21n3.52938
DOI: http://dx.doi.org/10.15446/abc.v21n3.52938
THE MANDIBULAR MUSCLES OF Puma concolor (MAMMALIA, CARNIVORA, FELIDAE)
Músculos mandibulares de Puma concolor (Mammalia, Carnivora, Felidae)
Romina Paola LLANOS1; Analía ANDRADE1; Lucio Manuel IBIRICU2; Marcelo CARRERA3.
1 Instituto Patagónico de Ciencias Sociales y Humanas. Centro Nacional Patagónico (CENPAT-CONICET). Bvd. Brown 2915. Puerto Madryn, Argentina.
2 Instituto Patagónico de Geología y Paleontología. Centro Nacional Patagónico (CENPAT-CONICET). Bvd. Brown 2915. Puerto Madryn, Argentina.
3 Cátedra de Mastozoología y Cátedra de Anatomía Sistemática y Aplicada en Mamíferos, Facultad de Ciencias Naturales, Universidad Nacional de la Patagonia San Juan Bosco. Bvd. Brown 3051. Puerto Madryn, Argentina.
For correspondence. rllanos@cenpat-conicet.gob.ar
Received: 7th September 2015, Returned for revision: 4th January 2016, Accepted: 28th January 2016.
Associate Editor: Nubia Estela Matta Camacho.
Citation/Citar este artículo como: Llanos RP, Andrade A, Ibiricu LM, Carrera M. The mandibular muscles of the Puma concolor (Mammalia, Carnivora, Felidae). Acta biol. Colomb. 2016;21(3):657-660. DOI: http://dx.doi.org/10.15446/abc.v21n3.52938
ABSTRACT
This manuscript describes the mandibular muscles of the puma (Puma concolor), a widely sparse American carnivorous mammal. The muscles on the right and left side of the head of three specimens -two adult males and one young female- were dissected and photographed. This study increases our knowledge on the mandibular soft tissue and adds important anatomical information to the poorly known and documented musculature of this felid, the largest of the subfamily Felinae.
Keywords: mandible, musculature, origin and insertion, Puma.
RESUMEN
En este trabajo se describen los músculos mandibulares del puma (Puma concolor), un mamífero carnívoro de amplia distribución en América. Se diseccionaron y fotografiaron los músculos izquierdos y derechos de la cabeza de tres individuos, dos machos adultos y una hembra juvenil. Este estudio incrementa nuestro conocimiento de los tejidos blandos mandibulares y por lo tanto, aporta información anatómica valiosa de la escasamente conocida y documentada musculatura de este félido, el más grande de la subfamilia Felinae.
Palabras clave: mandíbula, musculatura, origen e inserción, Puma.
The puma (Puma concolor, Linnaeus 1771) is among the largest felids of the world. Its size is just below the size of tigers (Panthera tigris, Linnaeus 1758), lions (Panthera leo, Linnaeus 1758), jaguars (Panthera onca, Linnaeus 1758) and leopards (Panthera pardus, Linnaeus 1758) (Nowell and Jackson, 1996). Nevertheless, specific features (e.g. shape of its nose, morphology of its feet, shape of its pupils, lacking the specialized larynx and hyoid) relate the puma with the smaller felids (Sunquist and Sunquist, 2002). Even, its skull has reached pantherine cranial size but retaining the cranial proportions of small cats (Werdelin, 1983). The species was originally nested as Felis concolor (Linneaus, 1771) but was recently recognized as Puma concolor, with Puma accepted as a separate genus by Ewer (1973) (Culver, 2010). It was placed in the subfamily Felinae. This species is also commonly known as the mountain lion, cougar, or catamount (Nowell and Jackson, 1996).
The puma is widely distributed in America, ranging from Alaska to southernmost Chile and Argentina (Currier, 1983; Iriarte et al., 1990; Sunquist and Sunquist, 2002). It occupies diverse habitats, including arid desert, tropical rainforest and cold coniferous forest (from sea level up to 5800 m in the Andes) (Redford and Eisenberg, 1992). Size of pumas increases with latitude and varies among the subspecies (Iriarte et al., 1990); average weights range between 53 kg and 72 kg for males and between 34 kg and 48 kg for females (Currier, 1983; Nowell and Jackson, 1996). This species is listed in the International Union for Conservation of Nature and Natural Resources (IUCN) red list as Least concern (IUCN, 2015).
There are few studies describing the anatomy of pumas, the second largest feline in America, with an important occurrence not only geographically but also in abundance. Concha et al. (2004) described the intrinsic muscles of thoracic limb. The muscles of pelvic limb were reported by Caro-Munizaga and Concha (2014). Antón et al., (2004) provided information on the muscle attachments of the mastoid region of puma. Hast (1989) presented an anatomical study of larynxes of several felid species, including the puma. However, the musculature participating in the movements of the jaw has been poorly studied. Our aim was to describe the muscles of the jaw, which are directly related to masticatory function in the puma, describing their origins and insertions. This study will enhance our understanding of jaw musculature of this American mammal and will also constitute an important source of information on soft tissue anatomy for other studies.
The muscles on the right and left side of the head of three specimens of puma, Puma concolor puma, were dissected. Faunal Department of Chubut Province (Dirección de Fauna y Flora Silvestre del Chubut, DFyFS) granted these puma specimens for the study. Collection numbers (DFyFS 288, 501 and 2e) are tentative until DFyFS completed the inventory of this faunal collection (housed at the Instituto Patagónico de Ciencias Sociales y Humanas, CENPAT-CONICET).
DFyFS 288 and DFyFS 501 corresponded to heads of adult males (between two and three years old). These animals were hunted in 2011 and 2013 in sheep ranges at the Biedma and Telsen departments respectively (north of Chubut Province, Patagonia, Argentina) and their heads were stored at the Centro Nacional Patagónico (CENPAT-CONICET). They were hunted by sheep farmers with permission of the provincial government. In 2005 puma was declared an injurious species for livestock in Chubut province and its capture is bounty (Law N° XVII 52). DFyFS 288 was preserved in 10% formaldehyde, so the observation of muscle fibers had some limitations related to this kind of preservation. DFyFS 501 was frozen at -20° C until dissection which took place 48 hours after freezing. DFyFS 2e corresponded to a young female (between eight months and one year old). This animal was hit by a car on August 2013 near Itsmo Ameghino, Peninsula Valdes Protected Area (Chubut Province, Argentina) and then transported to CENPAT for its study. The animal was frozen at -20° C until dissection. DFyFS 501 and DFyFS 2e were fresh, so the observation of muscle fibers was optimal.
In all specimens, dissection proceeded from the most external muscle layer and each muscle was documented with digital photographs. The origins and insertions of each muscle were recorded. A skull of puma was available for comparison and identification of skulls features during dissection. The muscular nomenclature used by Diogo et al. (2012) was employed. The following masticatory muscles were dissected in order to determine their origin and insertion: the masseter, temporalis, digastricus, pterygoideus medialis, pterygoideus lateralis, and mylohyoideus.
The masseter (Fig. 1A) is a broad muscle on the lateral side of the face and its function is to elevate the mandible. This muscle has been described in felids such as domestic cat (Felis catus, Linnaeus 1758), serval (Leptailurus serval, Schreber 1776), and tiger (Panthera tigris) (Sasaki et al., 2000; Laison et al., 2001; Diogo et al., 2012). In L. serval and P. tigris, the masseter is divided into three layers (ventral, superficial and deep bundles) but in the present study only two layers were distinguishable: a superficial bundle (superficialis), which was the largest, and a deeper bundle profundus. The division of the masseter muscle into three layers or bundles is a feature well represented within felids (Diogo et al., 2012). Nevertheless, in the P. concolor only two layers were present as in the small felids such as F. catus (Marcelo Carrera Pers. Obs., but see Laison et al., 2001) This muscular configuration could be related particularly to the taxonomic relationship of P. concolor within Felidae (i.e., the most closet relationship between Domestic cat lineage and Puma lineage versus Caracal + Panthera lineages, see Johnson et al., 2006). The masseter originates on the ventral border of the zygomatic arch and inserts onto the ventral part of the masseteric fossa. The origin and insertion are by tendons and fleshy fibers. The small deeper or ventral layer of the masseter attach to ventral and ventromedial portion of the ramus of the mandible. The layer superficialis, which occupies the major portion of the masseteric fossa, inserts mainly on the ventrolateral surface of the mandible (i.e., angular part). Both layers and bundles of the masseter muscle blended, posteriorly, the pterygoideus (see below).
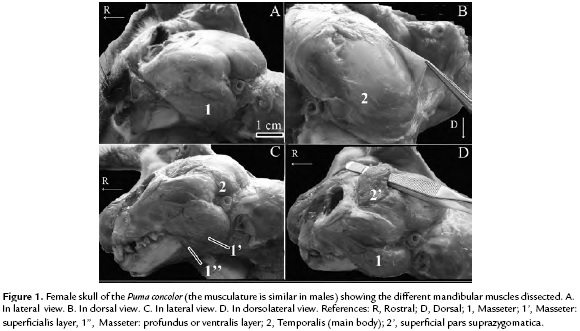
The temporalis (Fig. 1B) is a large muscle in carnivores (Turnbull, 1970) and its function is to elevate the mandible. As observed by Diogo et al., (2012) in servals and tigers and by Laison et al., (2001) in domestic cats, the temporalis is divided into two parts, superficial pars suprazygomatica and the largest portion of the muscle, the deeper main body (Fig. 1C). Both parts are separated by an aponeurosis, important feature of the temporalis muscle in carnivores. The main body is subdivided into an anterior and posterior superficial layer. The well-developed pars suprazygomatica (Fig. 1D) arises from the posterolateral portion of the skull and the medial side of the zygomatic arch. This muscle reaches the lateral part of the temporal crest. Both, the origin and the insertion, are via tendon and fleshy fibers. The largest layer of the temporalis (i.e., the deeper main body), originates from the temporal lines and the temporal fossa of the skull, also covering the caudal margin of the orbital ligament, whereas its insertion is in the coronoid process. The main body and its corresponding subdivision of the temporalis, is found in most groups of carnivores as well as other mammals, but the broad attachment of the deeper layer is a common feature present in carnivorans and just few other mammals (Diogo et al., 2012).
The digastricus, a mandibular depressor muscle (Fig. 2A), is divided into two parts: rostral and caudal portions (named as digastricus anterior and digastricus posterior by Diogo et al., 2012). The former is a mandibular muscle whereas the caudal portion is a hyoid muscle. Therefore, because the aim of this work was focused on the mandibular muscles, just the rostral portion of the digastricus is currently described. This is a relatively long and thin muscle. This ventral mandibular muscle originates at the mastoid process of temporal and jugular process of occipital of skull and inserts via fleshy fibers in the ventromedial surface of the mandible.
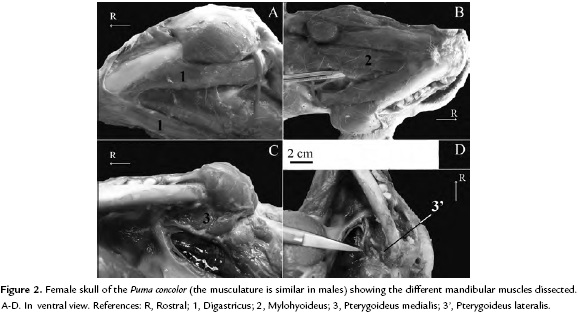
The mylohyoideus (Fig. 2B) is a flat muscle and blind to the geniohyoideus muscle. It extends from the mylohyoid line running from the ventromedial portion of the body of the mandible. This muscle, related to the movements of the tongue and the elevation of the hyoid, inserts via a flat tendon (i.e., aponeurotically) to the hyoid apparatus as in other felids (see Diogo et al., 2012).
Within the ventral mandibular muscles is the pterygoideus medialis and lateralis (Fig. 2C-D). The pterygoideus medialis is a robust muscle whereas the lateralis is a relatively thin muscle. The pterygoideus medialis, which is blended for the masseter, originates at the pterygopalatine fossa and inserts via tendon and fleshy fibers to the ventrolateral portion of the mandibular ramus. The pterygoideus lateralis, which is blended for the pterygoideus medialis (a feature present in all major carnivoran groups, see Diogo et al., 2012) originates at the lateral surface of the pterygoid bone and adjacencies and inserts to the condylar process of the mandible via a well-marked tendon.
The P. concolor is a widely sparse carnivoran mammal; however, several aspects particularly those related to its soft tissue anatomy are poorly known. In this context, this study provides novel information which can be used as a source for further studies for this or others species of Felidae. This study augments our knowledge of soft tissue mandibular anatomy of this still enigmatic American felid.
ACKNOWLEDGMENTS
We especially thank to Dirección de Fauna y Flora Silvestre de la Provincia del Chubut (Faunal Department of Chubut province) for offering us the animals for this study and to the anonymous reviewers for their suggestions. Gabriela Blanco helped with the English translation.
REFERENCES
Antón M, Salesa MJ, Pastor JF, Sánchez IM, Fraile S, Morales J. Implications of the mastoid anatomy of larger extant felids for the evolution and predatory behavior of sabretoothed cats (Mammalia, Carnivora, Felidae). Zool J Linn Soc. 2004;140(2):207-221. Doi: http://dx.doi.org/10.1111/j.1096-3642.2003.00093.x. [ Links ]
Caro-Munizaga C, Concha I. Descripción anatómica de los músculos del miembro pélvico del puma (Puma concolor). Int J Morphol. 2014;23(3):1085-1094. Doi: http://dx.doi.org/10.4067/S0717-95022014000300055. [ Links ]
Concha I, Adaro L, Borroni C, Altamirano C. Consideraciones anatómicas sobre la musculatura intrínseca del miembro torácico del puma (Puma concolor). Int J Morphol. 2004;22(2):121-125. Doi: http://dx.doi.org/10.4067/S0717-95022004000200004. [ Links ]
Culver M. Lessons and insights from evolution, taxonomy, and conservation genetics. In: Hornocker M, Negri S, editors. Cougar, Ecology and Conservation. Chicago: University of Chicago Press; 2010. p. 27-40. [ Links ]
Currier MJP. Felis concolor. Mamm Species. 1983;200:1-7. Doi: http://dx.doi.org/10.2307/3503951. [ Links ]
Diogo R, Pastor F, De Paz F, Potau JM, Bello-Hellegouarch G, Ferrero EM, Fisher RE. The head and neck muscles of the serval and tiger: homologies, evolution, and proposal of mammalian and a veterinary muscle ontology. Anat Rec. 2012;295(12):2157-2178. Doi: http://dx.doi.org/10.1002/ar.22589. [ Links ]
Ewer RF. The carnivores. Ithaca: Cornell University Press; 1973. p. 212. [ Links ]
Hast MH. The larynx of roaring and no roaring cats. J Anat. 1989;163:117-121. [ Links ]
Iriarte JA, Franklin WL, Johnson WE, Redford KH. Biogeographic variation of foods habits and body size of the America puma. Oecologia. 1990;85(2):185-190. Doi: http://dx.doi.org/10.1007/BF00319400. [ Links ]
International Union for Conservation of Nature and Natural Resources (IUCN), 2015. http://www.iucnredlist.org/details/18868/0. [ Links ]
Johnson WE, Eizirik E, Pecon-Slattery J, Murphy WJ, Antunes A, Teeling E, O'Brien SJ. The Late Miocene radiation of modern Felidae: A genetic Assessment. Science. 2006;311(5757):73-77. Doi: http://dx.doi.org/10.1126/science.1122277. [ Links ]
Laison F, Lautrou A, Azérad J, Pollin B, Lévy G. Superficial architecture of the jaw-closing muscles of the cat (Felis catus): the temporo-masseteric complex. C R Acad Sci III. 2001;324(9):855-862. Doi: http://dx.doi.org/10.1016/S0764-4469(01)01347-6. [ Links ]
Nowell K, Jackson P. Wild Cats. Status, Survery and Conservation Action Plan. Gland: IUCN/SSC Cat Specialist Group; 1996. 382 p. [ Links ]
Redford KH, Eisenberg JF. Mammals of the Neotropics, The Southern Cone: Chile, Argentina, Uruguay, Paraguay. Vol. 2. Chicago: University of Chicago Press; 1992. p. 172-173. [ Links ]
Sasaki M, Endo H, Yamamoto M, Kazuyoski A, Hayashi Y. The superficial layer of the musculus masseter and the well developed process of the maxilla in the tiger Panthera tigris. Mammal Study. 2000;25(1):27-34. Doi: http://dx.doi.org/10.3106/mammalstudy.25.27. [ Links ]
Sunquist M, Sunquist F. Wild cats of the World. Chicago: The University of Chicago Press; 2002. p. 253-255. [ Links ]
Turnbull WD. Mammalian masticatory apparatus. Fieldiana Geol. 1970;18(2):149-356. Doi: http://dx.doi.org/10.5962/bhl.title.5442. [ Links ]
Werdelin L. Morphological patterns in the skulls of cats. Biol J Linn Soc Lond. 1983;19(4):375-391. Doi: http://dx.doi.org/10.1111/j.1095-8312.1983.tb00793.x. [ Links ]

