










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.22 no.2 Bogotá May/Aug. 2017
https://doi.org/10.15446/abc.v22n2.56809
DOI: http://dx.doi.org/10.15446/abc.v22n2.56809
ACTIVIDAD ANTIMICROBIANA DE PÉPTIDOS CATIÓNICOS DISEÑADOS A PARTIR DE UN PÉPTIDO NEUTRO
Antimicrobial Activity of Cationic Peptides Designed from Neutral Peptide
José Fernando OÑATE-GARZÓN1, Marcela MANRIQUE-MORENO1, Edwin PATIÑO GONZALEZ1.
1 Instituto de Química, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia. Calle 67 No. 53-108. Medellín, Colombia.
For correspondence. edwin.patino@udea.edu.co
Received: 18th August 2016, Returned for revision: 22nd February 2017, Accepted: 26th February 2017.
Associate Editor: Juan F. Gonzalez.
Citation/Citar este artículo como: Oñate-Garzón JF, Manrique-Moreno M, Patiño Gonzalez E. Actividad antimicrobiana de péptidos catiónicos diseñados a partir de un péptido neutro. Acta biol. Colomb. 2017;22(2):157-164. DOI:http://dx.doi.org/10.15446/abc.v22n2.56809
RESUMEN
Los péptidos antimicrobianos (PAMs) juegan un papel importante en la inmunidad innata de la mayoría de los organismos; ellos pueden tener actividad en bacterias, hongos, virus y parásitos. El mecanismo de acción de los PAMs catiónicos yace en la capacidad de interactuar con membranas microbianas, debido a la superficie aniónica de dichas membranas. La familia de las cecropinas fue identificada como una de las familias peptídicas más importantes en los insectos. Los péptidos de esta familia, no contienen residuos de cisteína y son clasificados como helicoidales. Para estudiar el efecto de la carga sobre la estructura, nosotros introducimos residuos cargados positivamente en los primeros 18 aminoácidos de la región N-terminal de la cecropina-D (WT), y se evaluó la actividad biológica de los péptidos modificados. Dos análogos de la cecropina-D con cargas netas de +5 y +9, fueron obtenidos por síntesis de fase sólida (SSP). Los cambios en los péptidos análogos fueron generados de la siguiente manera: péptido +5 con tres sustituciones (E6R, E8R and Q12K) y péptido +9 con cinco sustituciones (E1R, E6R, E8R, Q12K, and D16K). La actividad antibacteriana fue evaluada en dos grupos de bacterias, con el fin de investigar los efectos de las cargas positivas en dicha actividad. Los péptidos catiónicos mostraron una mayor actividad antimicrobiana tanto en bacterias Gram-negativas como en Gram-positivas, a diferencia del péptido WT. Las representaciones en 3D de los péptidos mostraron que ellos tienen una estructura α-hélice. Nuestros resultados demostraron que cambios en la carga de los péptidos incrementa la actividad antibacteriana.
Palabras clave: carga catiónica, concentración mínima inhibitoria, Gallería mellonella, péptidos antimicrobianos, predicción de estructuras.
ABSTRACT
Antimicrobial peptides (PAMs) play an important role in the innate defense systems of most organisms; they act against bacteria, fungi, viruses and parasites. The mechanism of action of cationic PAMs rely on their capacity to interact with the anionic microbial membrane surface. The cecropin family was identified as one of the most important peptides in insects. Such peptides do not contain cysteine residues and are classified as α-helical. To study the effect of the charge on the peptide structure, we introduced positive charge residues in the last 18 residues at the N-end of cecropin-D (WT) and evaluated the biological activity of the modified peptides. Two analogous peptides from cecropin-D were obtained by synthesis of a solid phase (SSP) with charges of+5 and +9. The analogous peptides were generated as followed: peptide +5 with three substitutions (E6R, E8R and Q12K) and peptide +9 with five substitutions (E1R, E6R, E8R, Q12K, and D16K). Antibacterial activity was evaluated to investigate the effects of the positive charge in these two analogue peptides against two groups of bacteria. The cationic peptides showed higher antimicrobial activity against Gram-negative and Gram-positive bacteria than the WT peptide. The 3D representations of the peptides showed that they have α-helical structure. Our results demonstrate that changes in the charge of peptides increase the antibacterial activity.
Keywords: antimicrobial peptides, cationic charge, Gallería mellonella, minimal inhibition concentration, structure prediction.
INTRODUCCIÓN
Los péptidos antimicrobianos (PAMs) son un mecanismo de defensa preformado presente desde bacterias hasta mamíferos (Maróti Gergely et al., 2011). La acción de la mayoría de estas moléculas antimicrobianas está determinada por su naturaleza catiónica (Bulet et al., 2004). Los PAMs pueden variar de diez hasta 80 amino ácidos de longitud lo que los hace moléculas altamente variables. Los resultados provenientes de estudios estructurales evidenciaron diferentes patrones moleculares, los cuales se pueden agrupar en tres amplias clases: (1) péptidos lineales de α-hélice sin residuos de cisteína, (2) péptidos de hojas β, y (3) péptidos lineales ricos en residuos de prolina y/o glicina (Bulet et al., 1999; Bulet et al., 2004; Guaní-Guerra et al., 2010; Wang y Lai, 2010; Scocchi et al., 2011). En cuanto a la actividad antimicrobiana, los PAM son moléculas eficientes ya que presentan cuatro importantes ventajas: (i) son moléculas selectivas, (ii) su acción para matar microorganismos es rápida, (iii) poseen actividad frente a un amplio espectro de microorganismos, y (iv) los resultados de diferentes investigaciones indican que los microorganismos no desarrollan resistencia contra los PAM (Zasloff, 2002a; Zasloff, 2002b; Boman, 2003; Cytryńska et al., 2007; Duclohier, 2010).
La práctica convencional se basa en aislar los PAMs directamente de los mismos organismos. Sin embargo, la obtención de nuevos PAMs, se puede realizar a partir de una secuencia conocida (molde) con actividad antimicrobiana reportada. El cambio de algunos aminoácidos en el péptido molde se puede utilizar para aumentar la carga del péptido y así obtener un péptido catiónico. La estructura topológica del péptido molde es conservada, debido a que las sustituciones de amino ácidos son realizadas de acuerdo a la descripción hecha por Bordo y Argos (1991). En este trabajo se reporta la generación de dos PAMs catiónicos con cargas de +5 y +9 a partir de una secuencia molde neutra como la cecropina-D, reportada por Cytrynska et al. (2007), donde demostraron que este péptido tiene actividad antibacteriana. Este trabajo tiene por objeto investigar la influencia del incremento de la carga en derivados de la cecropina-D y el efecto en su actividad antibacteriana. La interacción de los péptidos análogos y el péptido silvestre (neutro) con las membranas de las bacterias estará determinado por que las bacterias poseen cargas negativas en su membrana celular por su contenido de cardiolipina y fosfatidilglicerol.
MATERIALES Y MÉTODOS
Cepas Bacterianas
Escherichia coli (E. coli) ATCC 25922 y Staphylococcus aureus (S. aureus) ATCC 25923, fueron obtenidas del American Type Culture Collection (ATCC; Rockville, MD, USA).
Secuencias de los Péptidos
La secuencia del péptido molde, cecropina-D (Neutro), fue tomada de Cytryńska y colaboradores (Cytryńska et al., 2007). Esta secuencia consta de 39 residuos (ENFFKEIERAGQRIRDAIISAAPAVETLAQAQKIIKGGD) y tiene una carga igual a 0 a pH fisiológico. La secuencia molde se utilizó para generar los péptidos análogos +5 (ENFFKRIRRAGKRIRDAIISAAPAVETLAQAQKIIKGGD) y +9 (RNFFKRIRRAGKRIRKAIISAAPAVETLAQAQKIIKGGD). Para el péptido +5 se realizaron los cambios de E6R, E8R y Q12K. Para el péptido +9 se realizaron las mismas sustituciones mencionadas previamente, más los cambios de E1R, y D16K. Las cargas de los tres péptidos se calcularon con algoritmo de GenScript Corporation, Piscataway, NJ, USA; Web: http://www.genscript.com.
Análisis de Similitud de las Secuencias de los Péptidos
Las secuencias de los tres péptidos fueron analizadas por medio de un alineamiento múltiple utilizando el programa ClustalO (Clustal Omega) del paquete de JalView; http://www.ebi.ac.uk/~michele/jalview/contents.html (Waterhouse et al., 2009). Adicionalmente, se realizó un alineamiento entre secuencias (LALIGN, EMBnet-Server; http://www.ch.embnet.org/software/LALIGN_form.html) utilizando BLOSUM62 (Artimo et al., 2012). La similitud entre las secuencias es reportada en porcentaje.
Síntesis y Purificación de Péptidos
Antes de la síntesis de los péptidos, se calcularon las propiedades de los tres péptidos con el programa de GenScript (Peptide Property Calculator). Los péptidos fueron sintetizados sin amidar el carbono terminal mediante síntesis Fmoc en fase sólida (SFS) (Bulet et al., 2004). Cada secuencia fue cargada en un sintetizador automático de péptidos (433, Applicated Biosystems Synthesizer) en la Universidad de Lausanne (Suiza). Luego de la síntesis, los péptidos se purificaron al 95 % mediante cromatografía líquida en Fase Reversa (RP-HPLC), usando una columna preparativa Vydac C-18 y una mezcla de (A) H2O con TFA 0,1 % (v/v) y (B) acetonitrilo (ACN) con TFA 0,1 % (v/v) como fase móvil. Para la elución de los péptidos, se utilizó el siguiente gradiente: 30 min con 5-70 % de B en un flujo de 1 mL/min y una longitud de onda de detección de 220 nm.
Concentración Mínima Inhibitoria (CMI)
Los cultivos bacterianos se realizaron en medio nutritivo y se incubaron a 37 °C toda la noche. Luego, el cultivo se llevó a una DO600 = 0,1 (Aprox. 1 x 108 bacterias) y fue diluido hasta alcanzar una concentración bacteriana de aproximadamente 1x105 unidades formadoras de colonia/ mL. Se tomaron 90 μL de cultivo, se incubaron por un tiempo, entre 18-20 horas a 37 °C con 10 μL de péptido.
Las concentraciones de los péptidos estuvieron en el rango de 100 hasta 0,6125 μM, siendo 100 μM la máxima concentración y 0,6125 μM la mínima concentración utilizada. Como control positivo se utilizó 1 μM Polimixina-B y 1 μM Oxitetraciclina para E. coli y S. aureus, respectivamente. Como control negativo se utilizó Buffer Fosfato Salino, PBS 1X (0,1M NaH2PO4, 0,1M Na2HPO4, pH 7,4). Posterior a la incubación, se determinó la turbidez de cada muestra en un plato de 96 pozos con una λ625 usando un espectrofotómetro Multiskan Go (Thermo Fisher Scientific), con el fin de obtener la Concentración Mínima Inhibitoria (CMI). La CMI es la primera concentración de antibiótico en la que se observa una inhibición del crecimiento bacteriano. Todos los ensayos se realizaron por triplicado utilizando diferentes cultivos.
Modelamiento de la Estructura 3D
Para el modelo estructural de la cecropina-D (Neutro), la secuencia se obtuvo de los datos publicados por Cytrynska y colaboradores (Cytryńska et al., 2007). Las secuencias finales de amino ácidos de los tres péptidos fueron cargadas en el programa I-TASSER (http://zhanglab.ccmb.med.umich.edu/I-TASSER/download/) para el modelamiento de la estructura 3D. Los parámetros estructurales que determinaron el modelo final de cada péptido fueron: 1) la calidad del modelo (C-score), 2) la trayectoria más frecuente (Cluster Density) y 3) número de señuelos estructurales (No of Decoys) (Zhang., 2008). Los archivos PDB (Protein Data Bank; http://www.rcsb.org/pdb/home/home.do) generados por I-TASSER fueron visualizados en el programa PyMol; https://www.pymol.org/ (Schrödinger, San Diego, California, USA). La conformación estructural de los modelos de los tres péptidos y su calidad estereoquímica fue evaluada determinando los ángulos phi (Φ) y psi (Ψ) con el software RAMPAGE Ramachandran Plot Assessment; http://mordred.bioc.cam.ac.uk (Lovell et al., 2003).
Predicción de la Digestión Peptídica por Enzima Estafilocócica
Una de las formas de resistencia bacteriana es la producción de proteasas capaces de digerir los péptidos antimicrobianos presente en el medio. Las secuencias de los péptidos fueron analizadas para predecir los sitios de clivaje por la peptidasa estafilocócica I usando el programa PeptideCutter del servidor de ExPasy (http://web.expasy.org/peptide_cutter/).
RESULTADOS
Análisis de Similitud
Los 39 residuos del péptido Neutro fueron comparados con los péptidos análogos +5 y +9 para determinar la similitud entre ellos. Los resultados del alineamiento global (Clustal Omega) muestran que los valores de similitud se extendieron en un rango de 87-95 % (Fig. 1). La comparación de las secuencias por BLAST (pairwise alignment), demostró que los péptidos +5 y + 9 tienen una similitud del 95 %. Mientras que el péptido Neutro y el péptido +9 tienen una similitud del 87 %, y el péptido Neutro y el péptido +5 presentan una similitud del 92 % (Fig. 1).
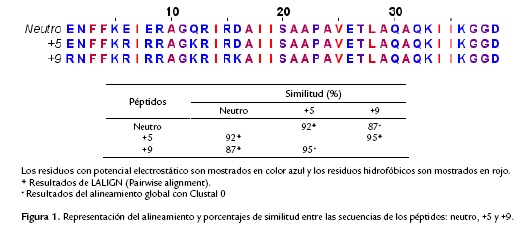
Secuencias de los Péptidos
El péptido silvestre (Neutro) (p85210, código UniProt KB), es una cecropina constituida por 39 aminoácidos aislada de la larva del insecto Galleria mellonella (Cytryńska et al. 2007). Esta cecropina de G. mellonella tiene una carga neutra a pH fisiológico, y su secuencia se utilizó como molde para la generación de los dos péptidos análogos con cargas de +5 y +9 (Fig. 1). Las cargas positivas se localizaron entre los primeros 18 residuos de la parte N-terminal (Fig. 1). El análisis por GenScript (Peptide Property calculator) generó como resultados las cargas de +5 y +9 para estas secuencias. Entonces, la sustitución de los residuos Glu6, Glu8 y Gln12 por Arg, Arg y Lys, respectivamente, resultó en un aumento de la carga del péptido en +5. Para el péptido con carga de +9, durante la síntesis se reemplazaron los aminoácidos de la siguiente manera: E1R, E6R, E8R, Q12K and D16K (figura 1). Los cambios de los amino ácidos se realizaron de acuerdo a los datos publicados por Bordo y Argos (1991) con el fin de no alterar la estructura topológica del péptido.
Concentración Mínima Inhibitoria (CMI)
Los tres péptidos fueron evaluados contra E. coli, y S. aureus. Los péptidos +5 y +9 mostraron las CMI más bajas en las bacterias Gram-negativas (Tabla 1). Para las bacterias Gram-positivas el péptido +5 necesitó una concentración 20 veces mayor que la utilizada para las bacterias Gram-negativas (de 2,5 μM hasta 50 μM). Por otro lado, en el péptido +9 su concentración se debió aumentar 4 veces más (de 1,25 μM hasta 5 μM) en las bacterias Gram-positivas (Tabla 1) para obtener el mismo efecto que en las Gram-negativas. Estos resultados muestran que el péptido +9 demostró la mayor actividad antibacteriana para ambos grupos de bacterias (Tabla 1). Por lo tanto, la diferencia en el aumento de la carga en la cara polar de la hélice N-terminal de los péptidos se puede relacionar con las diferencias en la actividad antimicrobiana, ya que, la hélice de la región C terminal no sufrió ningún cambio.

Estructura 3D de los péptidos
La modelación de la estructura 3D de los tres péptidos demostró que todos presentan una organización helicoidal. Los parámetros estructurales del modelo de cada péptido fueron: la calidad del modelo (C-score), el número de comparaciones estructurales del señuelo (Comparaciones, No. of Decoys) y la densidad de grupo que se define como la trayectoria del modelo más frecuente de la estructura que es por lo tanto el modelo de mejor calidad (Densidad de Grupo, Cluster density) (Tabla 2).
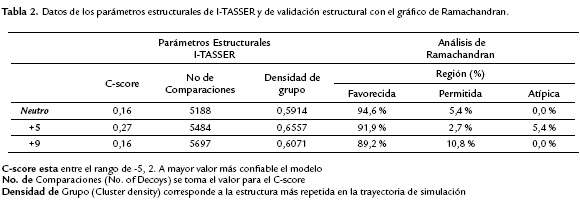
Cada péptido presenta dos hélices alfa con algunas diferencias entre ellas (Fig. 2). La hélice alfa de la región N-terminal del péptido neutro se extiende desde la Asn2 hasta la Ser20. La segunda hélice que va hasta el C-terminal se extiende desde la Ala24 hasta la Gly38. El lazo de conexión entre las dos hélices es de tres residuos conformado por Ala21, Ala22 y Pro23. Para el péptido +5, la hélice alfa de la región N-terminal es dos residuos más corta, esta hélice la conforman 17 residuos que se extienden desde Asn2 hasta Ile18. La segunda hélice está conformada de forma igual que en el péptido neutro. El lazo de conexión entre las dos hélices del péptido +5 se extiende dos residuos más, extendiéndose desde Ile19 hasta Pro23. Por otro lado, la estructura de la hélice N-terminal y el lazo de conexión en el péptido +9 son iguales al péptido neutro. Sin embargo, la segunda hélice (C-terminal) es dos residuos de glicina más corta (Gly37 y Gly38), esta hélice se extiende desde la Ala24 hasta la Lys36 (Fig. 2).
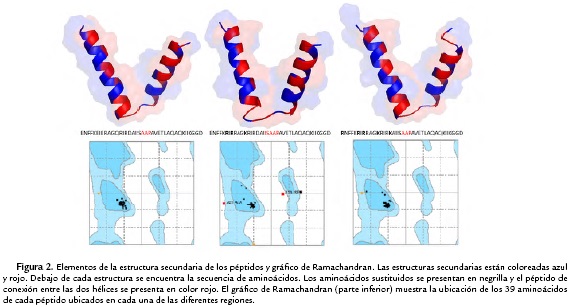
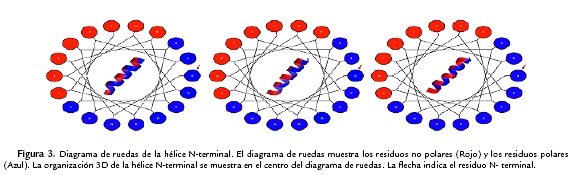
Las estructuras de los tres péptidos (archivos PDB) fueron analizadas con el programa RAMPAGE para determinar la distribución de los ángulos de torsión phi (Φ) y psi (Ψ) de los residuos de la cadena principal (Fig. 2, parte inferior y Tabla 2). Los resultados demuestran que para el péptidos neutro y +9, aproximadamente el 100 % de los residuos están en las regiones permitidas (región favorecida y región permitida). Sin embargo, para el péptido +5, aproximadamente el 5,4 % de los residuos se encuentran en las regiones atípicas (Tabla 2, Fig. 2). La organización de la cara polar o hidrofílica y la cara no-polar o hidrofóbica de la estructura 3-D de la hélice N-terminal se lograron visualizar en una figura 2-D con la aplicación de Kaeel (Fisher, 1996. http://kael.net/helical.htlm). La distribución espacial de los 18 amino acidos de la parte N-terminal de los tres péptidos muestran como la región no polar es idéntica en los tres péptidos (Fig. 3, residuos indicados en color rojo). Por otro lado, los cambios realizados en los péptidos análogos en la hélice N-terminal +5 y +9 son los que proporcionan la cationicidad de estos péptidos al tener un mayor número de residuos con carga positiva y son los responsables de las diferencias en la actividad antibacteriana (Tabla 1).
Predicción de la defensa de Staphylococcus aureus a la acción de los péptidos
Uno de los factores de virulencia que poseen lo microorganismos es la producción y liberación de proteasas al medio extracelular en el cual se encuentren. De acuerdo con nuestros resultados de CMI, los tres péptidos requieren de una mayor concentración para tener actividad contra S. aureus (Tabla 1). El péptido +9 requiere de cuatro veces más cantidad para inhibir el crecimiento de S. aureus que la que se utilizó para E. coli, mientras que con el péptido más +5 se utilizó 20 veces más que lo que se necesitó para las bacterias Gram negativas. Con el péptido Neutro no se observó inhibición del crecimiento de S. aureus aun con concentraciones tan altas como 100 μM (Tabla 1). Para determinar cualitativamente si la resistencia se debe en parte a proteasas excretadas por S. aureus, se realizó una predicción de sitios de corte en los tres péptidos. Para este fin, se utilizó el recurso bioinformático (ExPASY) para predecir los posibles sitios de corte en los péptidos Neutro, +5 y +9. Los resultados obtenidos con la herramienta para digestión de péptidos (peptide cutter from ExPASY) indica que los tres péptidos son digeridos por la enzima peptidasa estafilocócica I (Staphylococcal peptidase I) (Tabla 3). Nuestro análisis revela que el péptido neutro es el más fragmentado por la enzima peptidasa estafilocócica I. Por otro lado, los péptidos +5 y +9 son cortados por la enzima en dos y una ocasión, respectivamente.
DISCUSIÓN
La síntesis de péptidos sintéticos derivados de la secuencia conocida de la cecropina D, permitió investigar su aplicabilidad en la obtención de nuevas moléculas antimicrobianas. La combinación de los análisis de sustitución segura de residuos (Bordo y Argos., 1991), el alineamiento de secuencias (Waterhouse et al., 2009) y los modelos estructurales (Zhang., 2008), permitieron obtener dos moléculas nuevas con una alta homología a la secuencia molde y comparar la actividad antibacteriana.
Los datos de alineamiento demuestran que las secuencias tienen una alta similitud tanto a nivel de secuencia como estructural (Fig. 1 y Fig. 2), lo que sugiere que las diferencias en la actividad antibacteriana se deben a los cambios en las cargas positivas presentes en los péptidos análogos. De acuerdo a los resultados de la actividad antibacteriana (CMI, Tabla 1), los péptidos catiónicos tuvieron mayor actividad en contra de las bacterias Gram-negativas, mientras que el péptido Neutro presentó actividad antimicrobiana a concentraciones mucho más altas (Tabla 1). El aumento de residuos con carga positiva en la cara polar (Fig. 3) y en consecuencia, el aumento de la carga neta del péptido, han sido demostrados como uno de los factores importantes en la actividad antimicrobiana de estas moléculas (Giangaspero et al., 2001; Jiang et al., 2008; Yin et al., 2012), debido a que la densidad de carga de la superficie de la membrana determina la magnitud de la atracción electroestática y la atracción electroestática a su vez, conduce las moléculas del péptido cargadas positivamente hacía las membranas lipídicas de las bacterias cargadas negativamente. Los estudios de actividad antimicrobiana con el péptido V13K (carga de +7) demostraron que la reducción de la carga hasta +1 ó hasta -5, eliminó completamente la actividad contra Pseudomona aeruginosa (Jiang et al., 2008).
La influencia del aumento de las cargas en los péptidos para favorecer las interacciones electrostáticas y para tener una mejor actividad antibacteriana, también se ha evaluado en péptidos como las magaininas aisladas de anfibios. Sin embargo, aunque en los estudios con magaininas la carga tuvo un papel fundamental en la actividad antibacteriana, también hubo un límite máximo de cargas positivas (+5) necesarias para optimizar la acción contra la bacteria (Dathe et al., 2001). Para el péptido +9 análogo de la cecropina D de G. mellonella, la carga no restringió la actividad antibacteriana. Esto sugiere, que la amplia diversidad en cuanto a la carga de los PAMs en los organismos responde a su importante papel en la inmunidad innata.
Aunque varios reportes demuestran que el incremento de la carga en los péptidos está correlacionada con una mayor actividad antibacteriana, se hace necesario mantener los motivos estructurales del péptido molde en los péptidos análogos por la relación existente de estructura-función (Hwang., 1998; Kim et al., 2011). La figura 3 muestra que los cambios de los aminoácidos en la hélice N-terminal no alteró la estructura de la cecropina, ya que se mantuvo el sector hidrofílico más grande que el sector hidrofóbico en la hélice N-terminal, característica encontrada en la familia de las cecropinas (Kim et al., 2011). La conservación de las características antes mencionadas en los péptidos análogos +5 y +9 son importantes, porque demuestra que la anfipaticidad de la hélice no se alteró, así la parte hidrofóbica tendrá interacción con las cadenas acilicas de los lípidos, después de que la parte hidrofílica de la hélice N-terminal interaccione con las cabezas polares de los fosfolípidos de las membranas microbianas (Lee., 2010; Kim et al., 2010; Hwang et al., 2011; Kim et al., 2011; Lee et al., 2013). Interesantemente, los péptidos +5 y +9 tuvieron menos efecto sobre la bacteria Gram-positiva (S. aureus), a pesar de que en sus membranas existen mayor proporción de fosfolípidos aniónicos que en las Gram-negativas (Teixeira et al., 2011). Estos datos están en concordancia con los resultados previamente publicados por Kim y colaboradores, en donde se demuestra que los péptidos de la familia de las cecropinas tienen selectividad por las bacterias Gran-negativas (Kim et al., 2011).
Por otro lado, la resistencia de S. aureus hacia estos péptidos antimicrobianos puede estar soportada por la susceptibilidad de los péptidos a la digestión por proteasas secretadas por esta bacteria (Tabla 3). Diferentes investigaciones demuestran cómo estas proteasas extracelulares son mediadores claves en la virulencia de estafilococos (Lowy., 2003; Kim et al., 2011; Kolar et al., 2013). Las investigaciones en análisis de proteomica de S. aureus, revelaron un aumento en la abundancia de ciertas proteínas incluyendo diez proteasas. Además, la deleción de los genes de las proteasas ocasionó una mayor sensibilidad en las bacterias a los efectos de péptidos antimicrobianos (Kolar et al., 2013). Para los tres péptidos estudiados, encontramos que el péptido Neutro es el más susceptible a la digestión con la peptidasa estafilocócica, seguido por el péptido +5 y +9 (Tabla 3).
En la modelación estructural de los péptidos se demuestra que sus estructuras son similares a la estructura del péptido Papiliocina (PDB 2LA2) encontrada en el portal del PDB y determinada por resonancia magnética nuclear (NMR). El péptido Papiliocina es una cecropina proveniente de mariposa (Papilio xuthus), esta cecropina posee 37 aminoácidos y tiene una carga de +8 a pH fisiológico (Kim et al., 2011). Las cecropinas fueron los primeros péptidos helicoidales descubiertos y su estructura está determinada por dos hélices alfa unidas por un lazo flexible similar a la estructura mostrada por la Papiolicina. Las características de carga de las α-hélices de las cecropinas es que la hélice C-terminal es más hidrofóbica que la hélice N-terminal (Lee et al., 2013). Los estudios de la actividad antibacteriana de péptido Papiliocina demostraron que la cationicidad influyó en una actividad antibacteriana mayor (Lee et al., 2013). En este estudio se muestra que la estructura terciaria del péptido neutro de G. mellonella (Cecropin-D like) no fue alterada por los cambios en los aminoácidos introducidos en los péptidos +5 y +9 y que los 39 residuos de estos péptidos se organizaron en las dos α-hélices características de las cecropinas y que estas dos hélices están unidas por una bisagra de 3-5 residuos. Al igual que los datos reportados por Kim y colaboradores (Kim et al., 2011), la cationicidad de los péptidos +5 y +9 fue un factor determinante en la actividad antimicrobiana. Estudios previos han demostrado que la región N-terminal es la responsable de la actividad antibacteriana y que el aumento de los residuos catiónicos en esta región de los péptidos como la dermaseptina y péptidos derivados de catelicidinas tiene como efecto un aumento de la capacidad del péptido para lisar microorganismos (Nicolasq., 1994; Skerlavaj et al., 1996).
Nuestros resultados confirman estos hallazgos ya que los cambios se realizaron en la parte N-terminal del péptido neutro para obtener los péptidos +5 y +9. Estos cambios disminuyeron la hidrofobicidad de 0,232 del péptido neutro hasta 0,193 y 0,178 para +5 y +9, respectivamente. Los cambios en la hidrofobicidad también se pudieron observar en los tiempos de retención de la cromatografía en fase reversa en donde el péptido neutro mostró un tiempo de retención de 1,849 min mientras que los péptidos +5 y +9 mostraron tiempos de retención similares 1,441 y 1,442 min (datos no mostrados).
CONCLUSIONES
En conclusión, los péptidos análogos a la ceropina D de Galleria mellonella tienen la mayor actividad antibacteriana. Aunque, no conocemos con exactitud los mecanismos moleculares responsables de la actividad antibacteriana, nuestros resultados proporcionan una vía funcional para expandir el número de péptidos con actividad antimicrobiana lo cual puede significar una opción para uso terapéutico.
AGRADECIMIENTOS
Oñate-Garzón le agradece a COLCIENCIAS por la financiación del doctorado y por toda la colaboración que hizo posible este trabajo.
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
REFERENCIAS
Artimo P, Jonnalagedda M, Arnold K, Baratin D, Csardi G, De Castro E, et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012;40:597-603. Doi:10.1093/nar/gks400. [ Links ]
Boman HG. Antibacterial peptides: basic facts and emerging concepts. J Inter Med. 2003;254:197-215. Doi:10.1046/j.1365-2796.2003.01228.x. [ Links ]
Bordo D, Argos P. Suggestions for "safe" residue substitutions in site-directed mutagenesis. J Mol Biol. 1991;217:721-729. Doi:10.1016/0022-2836(91)90528-E. [ Links ]
Bulet P, Stöcklin R, Menin L. Antimicrobial peptides: from invertebrates to vertebrates. Immunol Rev. 2004;198:169-84. Doi:10.1111/j.0105-2896.2004.0124.x. [ Links ]
Bulet P, Hetru C, Dimarca JL, Hoffmann D. Antimicrobial peptides in insects; structure and function. Dev Comp Immunol. 1999;23:329-344. Doi:10.1016/S0145-305X(99)00015-4. [ Links ]
Cytryńska M, Mak P, Zdybicka-Barabas A, Suder P,Jakubowicz T. Purification and characterization of eight peptides from Galleria mellonella immune hemolymph. Peptides. 2007;28:533-546. Doi:10.1016/j.peptides.2006.11.010. [ Links ]
Duclohier H. Antimicrobial peptides and peptaibols, substitutes for conventional antibiotics. Acta biol. Colomb., 22(2):157-164, mayo-agosto 2017 Doi:10.2174/138161210793292500. [ Links ]
Dathe M, Nikolenko H, Meyer J, Beyermann M, Bienert M. Optimization of the antimicrobial activity of magainin peptides by modification of charge. FEBS Lett. 2001;501:146-150. Doi:10.1016/S0014-5793(01)02648-5. [ Links ]
Giangaspero A, Sandri L, Tossi A. Amphipathic alpha helical antimicrobial peptides. FEBS. 2001;268:5589-5600. Doi: 10.1046/j.1432-1033.2001.02494.x. [ Links ]
Guaní-Guerra E, Santos-Mendoza T, Lugo-Reyes SO, Terán LM. Antimicrobial peptides: general overview and clinical implications in human health and disease. Clin Immunol. 2010;135D1-11. doi:10.1016/j.clim.2009.12.004. [ Links ]
Hwang PM, Vogel HJ. Structure-function relationships of antimicrobial peptides. Biochem Cell Biol. 1998;76:235-246. Doi: 10.1139/o98-026. [ Links ]
Hwang B, Hwang JS, Lee J, Kim JK, Kim SR, Kim Y, et al. Induction of yeast apoptosis by an antimicrobial peptide, Papiliocin. Biochem Biophys Res Commun. 2011;408:89-93. Doi:10.1016/j.bbrc.2011.03.125. [ Links ]
Jiang Z, Vasil AI, Hale JD, Hancock REW, Vasil ML, Hodges RS. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic α-helical cationic antimicrobial peptides. Biopolymers. 2008;90:369-383. Doi:10.1002/bip.20911. [ Links ]
Kim J-K, Lee E, Shin S, Jeong K, Lee J-Y, Bae S-Y, et al. Structure and function of papiliocin with antimicrobial and antiinflammatory activities isolated from the swallowtail butterfly, Papilio xuthus. J Biol Chem. 2011;286:41296-41311. Doi:10.1074/jbc.M111.269225. [ Links ]
Kim SR, Hong MY, Park SW, Choi KH, Yun EY, Goo TW, et al. Characterization and cDNA cloning of a cecropin-like antimicrobial peptide, papiliocin, from the swallowtail butterfly, Papilio xuthus. Mol Cells. 2010;29:419-423. Doi:10.1007/s10059-010-0050-y. [ Links ]
Kolar SL, Ibarra JA, Rivera FE, Mootz JM, Davenport JE, Stevens SM, et al. Extracellular proteases are key mediators of Staphylococcus aureus virulence via the global modulation of virulence-determinant stability. Microbiology Open. 2013;2:18-34. Doi:10.1002/mbo3.55. [ Links ]
Lee J. Membrane perturbation induced by papiliocin peptide, derived from Papilio xuthus, in Candida albicans. J Microbiol Biotech. 2010;20:1185-1188. Doi:10.4014/jmb.1004.04014. [ Links ]
Lee E, Jeong KW, Lee J, Shin A, Kim JK, Lee J, et al. Structure-activity relationships of cecropin-like peptides and their interactions with phospholipid membrane. BMB Reports. 2013;46:282-287. Doi:10.5483/BMBRep.2013.46.5.252. [ Links ]
Lovell S, Davis I, Arendall IIIB, Bakker P, Word M, Prisant M, Richardson J, Richardson D. Structure validation by Cα geometry: φ Ψ and Cβ deviation Proteins. Struct, Funct, Bioinf. 2003;50:437-50. [ Links ]
Lowy FD. Antimicrobial resistance: the example of Staphylococcus aureus. The Journal of clinical investigation. Am Soc Clin Invest; 2003;11 1:1265-1273. Doi:10.1172/JCI18535. [ Links ]
Maróti Gergely G, Kereszt A, Kondorosi É, Mergaert P. Natural roles of antimicrobial peptides in microbes, plants and animals. Res Microbiol. 2011;162:363-374. Doi:10.1016/j.resmic.2011.02.005. [ Links ]
Nicolas P and Mor A. The NH2-terminal α-Helical Domain 1-18 of dermaseptin is responsible for antimicrobial activity. J Biol Chem. 1994;269:1934-1939. [ Links ]
Scocchi M, Tossi A, Gennaro R. Proline-rich antimicrobial peptides: converging to a non-lytic mechanism of action. Cell Mol Life Sci. 2011;68:2317-2330. Doi:10.1007/s00018-011-0721-7. [ Links ]
Skerlavaj B, Gennaro R, Bagella L, Merluzzi L, Risso A, Zanetti M. Biological characterization of two novel cathelicidin-derived peptides and identification of structural requirements for their antimicrobial and cell lytic activities. J Biol Chem. 1996;271:28375-28381. Doi:10.1074/jbc.271.45.28375. [ Links ]
Teixeira V, Feio MJ, Bastos M. Role oflipids in the interaction of antimicrobial peptides with membranes. Prog Lipid Res. 2012; 51: 149-177. Doi:10.1016/j.plipres.2011.12.005. [ Links ]
Wang Y-P, Lai R. Insect antimicrobial peptides: structures, properties and gene regulation. Zoo Res. 2010;31:27-34. Doi:10.3724/SP.J.1141.2010.01027. [ Links ]
Waterhouse AM, Procter JB, Martin DMa, Clamp M, Barton GJ. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics. 2009;25: 1189-1191. Doi:10.1093/bioinformatics/btp033. [ Links ]
Yin LM, Edwards MA, Li J, Yip CM, Deber CM. Roles of hydrophobicity and charge distribution of cationic antimicrobial peptides in peptide-membrane interactions. J Biol Chem. 2012;287:7738-7745. Doi:10.1074/jbc.M111.303602. [ Links ]
Zasloff M. Trypsin, for the defense. Nature Immunol. 2002a;3:508-510. Doi:10.1038/ni0602-508. [ Links ]
Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002b;415:389-395. Doi:10.1038/415389a. [ Links ]
Zhang Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics. 2008;9:40. Doi:10.1186/1471-2105-9-40. [ Links ]

