










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.22 no.2 Bogotá May/Aug. 2017
https://doi.org/10.15446/abc.v22n2.61161
DOI: http://dx.doi.org/10.15446/abc.v22n2.61161
ANÁLISIS ESPACIAL DE LA INCIDENCIA DE ENFERMEDADES EN DIFERENTES GENOTIPOS DE CACAO (Theobroma cacao L.) EN EL YOPAL (CASANARE), COLOMBIA
Spatial Analysis of Diseases Incidence in Different Cocoa Genotypes (Theobroma cacao L.) in Yopal (Casanare), Colombia
Néstor Julián CÁRDENAS PARDO1, Aquiles DARGHAN1, Miguel Darío SOSA RICO2, Alia RODRIGUEZ3.
1 Facultad de Ciencias Agrarias, Universidad Nacional de Colombia, Ciudad Universitaria, Sede Bogotá. Av. Carrera 30 n°. 45-03. Bogotá D.C., Colombia.
2 Programa de Ingeniería Agronómica-Utopía, Facultad de Ciencias Agropecuarias, Universidad de la Salle. El Yopal, Casanare, km 12 vía Yopal - Matepantano, Colombia.
3 Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Ciudad Universitaria, Sede Bogotá. Av. Carrera 30 n°. 45-03. Bogotá D.C., Colombia.
For correspondence. njcardenaspa@unal.edu.co
Received: 23rd November 2016, Returned for revision: 27th March 2017, Accepted: 26th April 2017.
Associate Editor: Xavier Marquínez Casas.
Citation/Citar este artículo como: Cárdenas Pardo NJ, Darghan A, Sosa Rico MD, Rodriguez A. Análisis espacial de la incidencia de enfermedades en diferentes genotipos de cacao (Theobroma cacao L.) en El Yopal (Casanare), Colombia. Acta biol. Colomb. 2017;22(2):209-220. DOI: http://dx.doi.org/10.15446/abc.v22n2.61161
RESUMEN
Las enfermedades "pudrición negra del fruto" y "moniliasis" son reportadas como limitantes en plantaciones de cacao en Colombia. Fueron evaluados los patrones espaciales de incidencia para estas dos enfermedades en un cultivo de cacao durante un monitoreo de trece semanas en campo. Semivariogramas e interpolación kriging fueron usados para analizar el comportamiento espacial de "pudrición negra del fruto", los estadísticos Join Count e Indice de Moran se emplearon para estudiar el comportamiento espacial de "moniliasis". La incidencia de "pudrición negra del fruto" presentó estructura de dependencia espacial de acuerdo a los semivariogramas durante el tiempo de evaluación, mientras que la autocorrelación espacial de sitios de infección de "moniliasis" se distribuyó de forma aleatoria entre las plantas de cacao según el Indice de Moran. El rango de dispersión estimado de "pudrición negra del fruto" varió entre 21,74 y 40,23 m y los mapas kriging evidenciaron el progreso espacio-temporal de la enfermedad durante las semanas evaluadas. Las condiciones experimentales dadas y la aproximación de los análisis espaciales, contribuyeron al entendimiento de la dinámica espacial de dos enfermedades limitantes en tres clones de cacao en la Orinoquía Colombiana.
Palabras clave: cacao, dependencia espacial, dispersión, modelos estadísticos, moniliasis, pudrición negra del fruto.
ABSTRACT
"Black pod" and "frosty pod" diseases are reported as limiting in cocoa plantations in Colombia. Spatial patterns of incidence for these two diseases were evaluated during a thirteen weeks long period of monitoring in the field. Semivariogram and kriging interpolation were used to analyze the spatial behavior of "black pod" and Join Count and Moran's Index statistics were used to analyze the spatial behavior of "frosty pod". The incidence of "Black pod" has a spatial dependence structure according to semivariogramas during the evaluation time, while spatial autocorrelation of "moniliasis" infection sites was randomly distribuited among cocoa plants according to the Moran's Index. The estimated dispersion range of "Black pod" was from 21.74 to 40.23 m y the kriging maps showed the spatio-temporal progression of the disease during the evaluated weeks. The experimental conditions given and spatial analysis approach, contributed to the understanding of the spatial dynamics of two limiting diseases in three clones of cocoa in the Colombian Orinoquia.
Keywords: black pod rot, cocoa, disease, dispersal, frosty pod rot, spatial dependence, statistical models.
INTRODUCCIÓN
El cacao (Theobroma cacao L.) es un sistema productivo agrícola de interés creciente en Colombia (ICA, 2013; Rodríguez y Vera, 2015). Sin embargo, enfermedades como "moniliasis" (MS) y "pudrición negra del fruto" (PNF) son reportadas como limitantes en este cultivo a nivel nacional (Jaimes y Aranzazu, 2010; Jaimes et al., 2011; Correa et al., 2014; Rodríguez y Vera 2015; Ramírez, 2016). Ambas enfermedades generan pudrición en el fruto de cacao, órgano de interés agronómico y comercial en las plantaciones (Ramírez, 2016). MS impacta más del 40 % de la producción anual de cacao en el país (Correa et al., 2014), mientras que la incidencia de PNF viene aumentando en los últimos años y se reporta como la mayor limitante en algunos lotes productivos monoclonales en Colombia (Rodríguez y Vera 2015; Ramírez, 2016). En el mundo, esta última reporta pérdidas anuales entre 10 y 30 % en producción de granos, con valores superiores en condiciones de mayor humedad (McMahon y Purwantara, 2004).
La MS es causada por Moniliophthora roreri (Cif y Par) Evans et al. (Aime y Phillips-Mora, 2005), patógeno hemi-biótrofo conocido como un ascomicete anamórfico, perteneciente a la familia Marasmiaceae (Aime y Phillips-Mora, 2005), cuyas estructuras de infección son esporas que para su formación y dispersión, no requieren de alta humedad y su liberación es dada por corrientes de aire y vibraciones en las plantas durante la cosecha (Evans, 1981). La PNF por su parte, es causada por especies del oomicete Phytophthora, ocho de ellas se han aislado en frutos de cacao con las mayores pérdidas en producción mundial causadas por P. palmivora, P. megakarya y P. citrophthora (McMahon y Purwantara, 2004). En Antioquia (Colombia), Ramírez (2016) reporta a P. palmivora y otra especie de Phytophthora sp. no identificada con incidencias en frutos de cacao de 92 y 7,5 % respectivamente, en zona de vida de bosque húmedo tropical. La dispersión de esta enfermedad y el desarrollo de la epidemia ocurre al salpicar agua lluvia sobre los esporangios, encontrados en el suelo o en plantas infectadas, y por insectos vectores (McMahon y Purwantara, 2004).
La expresión y dispersión de las epidemias depende también de las características intrínsecas de susceptibilidad del hospedante. Así, en plantaciones de cacao en Colombia, se ha reportado al clon ICS95 como resistente a varias cepas de M. roreri (Phillips-Mora, 2005; Jaimes et al., 2011) y al clon CCN51 como moderadamente resistente al mismo patógeno en tres regiones del país, incluyendo la Orinoquía colombiana (Arauca) (Jaimes et al., 2011). En contraste, ICS95 y CCN51 se reportan como susceptible y altamente susceptible a Phytophthora spp. respectivamente, donde el uso de este último clon se incrementa cada vez más en el establecimiento de nuevas plantaciones a nivel nacional (Rodríguez y Vera 2015).
En general, una epidemia se define como el progreso de una enfermedad en el tiempo y espacio. Las condiciones ambientales y las características del patosistema determinan la estructura de la epidemia y sus patrones espaciales (van Maanen y Xu, 2003). El estudio de la dinámica de enfermedades en el espacio y tiempo contribuye al desarrollo de estrategias para el manejo de epidemias (Agrios, 2004). Sin embargo, el éxito de estas estrategias depende de la aplicación de métodos y modelos adecuados, que representen el gradiente de enfermedad en el espacio y/o la curva de progreso de la enfermedad en el tiempo, considerando la relación entre patógenos, hospedante, variables ambientales y manejo del cultivo (Campbell y Madden, 1990).
Inicialmente, los estudios de patrones espaciales de enfermedades se centraron en el análisis de la media y la varianza o en la distribución de frecuencias de la incidencia de la enfermedad (Madden y Hughes, 1995; Madden y Hughes, 1999). No obstante, este enfoque analítico no tiene en cuenta información sobre la ubicación de las muestras en el espacio, ni el grado de dependencia entre observaciones cercanas. Además, con el uso de la teoría de variables regionalizadas de la geoestadística, la varianza entre muestras se interpreta como una función de su distancia de separación (Rossi et al., 1992; Pizzato et al., 2014). Su objetivo es medir la dependencia espacial entre mediciones, tomadas a diferentes distancias y empleando esta información para interpolar, extrapolar y simular. En este sentido, el análisis espacial se ha definido como la evaluación cuantitativa de la variación o cambios de una población con orientación espacio-temporal dentro de un área específica (Florez y Corredor, 2000).
El modelamiento teórico y la descripción estadística de la dinámica espacio-temporal de las epidemias tiene un interés creciente, especialmente para datos de incidencia de enfermedad donde plantas individuales o partes de plantas son clasificadas como enfermas o sanas (van Maaden y Xu, 2003). Diferentes métodos han sido empleados para caracterizar la agregación espacial de datos provenientes de incidencia de enfermedad. Análisis de relaciones, varianza-media, ajuste de distribución beta-binomial, autocorrelación espacial y métodos geoestadísticos son métodos empleados para cuantificar la heterogeneidad espacial de datos de incidencia de enfermedad (van Maaden y Xu, 2003).
Así, es fundamental determinar de forma precisa, la dinámica de las enfermedades en campo para poder establecer estrategias de manejo adecuadas. Adicionalmente, la variación en la afectación de las plantas por PNF y MS puede ser originada a partir de la variabilidad espacial de condiciones conductivas a la enfermedad, tales como diferentes niveles de susceptibilidad en el material clonal y su alternabilidad en el establecimiento del cultivo. Por esto, es importante identificar las características de dependencia espacial de la incidencia de enfermedades y su dispersión en plantaciones de cacao.
Análisis por semivariogramas y kriging han sido empleados para detectar patrones de dispersión espacio-temporal de Phytophthora sp. en frutos de cacao. Ten Hoopen et al. (2009) estudiaron la relación espacio-temporal de la enfermedad PNF de cacao empleando análisis por semivariogramas, demostrando una dependencia espacial de la distribución de la enfermedad en el cultivo a partir de plantas monitoreadas durante dos épocas de producción.
En la actualidad, es escasa la información sobre el comportamiento espacial simultáneo de dos enfermedades limitantes en el cultivo de cacao, en condiciones del trópico ácido, en zonas de Suramérica con amplio potencial de producción. Esto puede limitar la aplicación de medidas más precisas de manejo de enfermedades y la implementación de estrategias para su control desde el establecimiento de la plantación. El objetivo de este trabajo fue caracterizar la variabilidad espacial de la incidencia de PNF y MS en una plantación compuesta por tres clones de cacao en El Yopal (Casanare, Colombia) empleando metodologías de análisis espacial.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en la finca "Mate Pantano" de la Universidad de La Salle ubicada en zona rural del municipio de El Yopal (Casanare), a una altura de 256 m s.n.m. y con coordenadas geográficas de 5°19'31"N y 72°17'48"W. Las condiciones de temperatura, precipitación y humedad relativa máximas y mínimas presentadas durante el tiempo de evaluación fueron de 27,6 - 23,3°C, 122,5-50,3 mm y 83 - 91 % respectivamente. Los datos meteorológicos fueron obtenidos a partir de una estación climatológica ubicada en la finca de la Universidad, que colectó y registró datos diarios en línea a través del software WeatherLink 5.9.2. Se delimitó y monitoreó un área de 1,4 hectáreas de una plantación de cacao de 4,5 años de edad, compuesta por tres materiales clonales. El área monitoreada se delimitó empleando la función track manager de un dispositivo GPS Garmin Etrex 30x, con base en criterios de homogeneidad del sistema productivo -sin sombrío en los tres clones presentes- y del terreno -sin cambios significativos en la pendiente, ni anegamientos-.
Datos espaciales y de enfermedad fueron colectados a partir de 135 plantas, correspondiendo a 45 plantas por clon de los materiales CCN51, ICS95 y FSA12, sembrados intercaladamente en el lote cada dos surcos por clon, a una distancia de 3 x 3 m entre plantas. Las plantas monitoreadas fueron seleccionadas a través del lote con base en la presencia de frutos de cacao jóvenes y maduros. Estos fueron evaluados con una frecuencia de siete días durante agosto, septiembre y octubre de 2015, meses correspondientes a la época de lluvias en la zona de estudio.
Variables analizadas
Se analizó la variable incidencia de las enfermedades MS y PNF de cacao y su comportamiento espacial en el lote. La incidencia para ambas enfermedades se calculó según la ecuación 1:
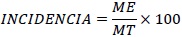
Dónde, ME = número de frutos enfermos por árbol y MT= número total de frutos por árbol. El número acumulado de frutos enfermos para cada planta fue calculado durante las semanas evaluadas.
Geo-referenciación
Se levantó un mapa del perímetro del lote monitoreado por georeferenciación, empleando el dispositivo GPS Garmin Etrex 30x. Se consideró una planta como punto de muestreo y las ubicaciones de las plantas monitoreadas fueron registradas en el mapa mediante coordenadas geográficas espaciales.
Identificación de las enfermedades
El reconocimiento de las enfermedades en campo se realizó siguiendo los síntomas y escalas de severidad reportadas en la literatura. La infección por MS presentó daños internos y externos, incluyendo lesiones acuosas pequeñas, deformación, maduración prematura y manchas de color chocolate -con límite no bien definido- en la parte exterior del fruto de cacao (Phillips-Mora y Wilkinson, 2007). En estados avanzados de infección, los tejidos internos formaron una masa compacta rodeada por una sustancia acuosa que causó la pérdida de las semillas contenidas (Phillips-Mora y Wilkinson, 2007). Los síntomas de PNF por su parte, se presentaron en la superficie del fruto como una mancha incolora en estado inicial, que posteriormente se desarrolló en una lesión de color marrón o negro con un límite bien definido, extendiéndose por todo el fruto en un tiempo de dos semanas aproximadamente (McMahon y Purwantara, 2004). Con frecuencia, las infecciones en frutos maduros iniciaron en la punta o base de estos (puntos distales del fruto), mientras que infecciones ecuatoriales se asociaron con daños en la superficie de los frutos o heridas (McMahon y Purwantara, 2004).
Análisis espacial
Incidencia "pudrición negra del fruto"
La variabilidad espacial (relaciones espaciales) de la incidencia de PNF entre puntos de muestreo (plantas) fue analizada por semivariogramas isotrópicos empíricos y ajustados empleando el procedimiento "variogram" del paquete estadístico SAS Institute Inc. versión 9.4®. Se realizó una transformación logarítmica a los valores de número acumulado de frutos enfermos por planta durante el tiempo de evaluación. Los parámetros descriptivos de los semivariogramas proporcionados por el programa estadístico fueron: 1) La varianza "pepita" (C0) o valor en el cual la curva del modelo calculado intercepta el eje y. 2) El rango (A) para el cual la dependencia espacial (en metros) es aparente. 3) El umbral que corresponde a la asíntota del modelo (C+C0). Los semivariogramas obtenidos permitieron cuantificar el grado y escala de la variación espacial, dados a través de la ecuación 2:
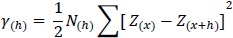
Donde Y(h) corresponde a la semivarianza de la incidencia de plantas de cacao infectadas en el espacio, separadas por los intervalos de las distancias h. N(h) es el número total de pares de plantas de cacao separadas por un intervalo de distancia h. Z(x) es el valor de la incidencia en una planta localizada en la posición x. Z(x+h) es el valor de la incidencia a la distancia del intervalo h desde x.
La dependencia espacial fue estimada a través del índice del grado de dependencia espacial (SDDI -por sus siglas en inglés-) basado en los valores (C) y (C0) de la ecuación 3 y la clasificación adoptada por Alves y Pozza (2010):
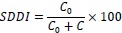
donde cuando SDDI ≤; 25%, existe una fuerte dependencia espacial. Cuando 25% ≤ SDDI ≤ 75%, una dependencia espacial moderada tiene lugar. Y cuando SDDI ≥ 75%, una pobre dependencia espacial es aparente.
El método de interpolación kriging fue empleado a partir de las propiedades estructurales de los semivariogramas obtenidos y los datos de las variables evaluadas. El indicador kriging, a partir del principio de dependencia espacial en el cual las muestras cercanas tienen mayor similitud entre sí que las más distantes, estima los valores de las variables en los puntos no muestreados a partir de los puntos muestreados. Los mapas kriging fueron construidos con el procedimiento "krige2d" del programa SAS Institute Inc. versión 9.4®.
Incidencia "moniliasis"
Se empleó el estadístico de prueba conocido como Join Count para estimar la autocorrelación espacial entre pares de puntos, a partir del número de pares de árboles con síntomas de la enfermedad, separados por una distancia y orientación específica. El Join Count, a nivel nominal, consideró la presencia o ausencia de MS. La distribución y dependencia espacial de esta propiedad se pudo expresar a nivel de una variable binaria, conteniendo solo dos valores asignados a la variable: presencia/ausencia. Join Count permitió caracterizar la naturaleza de la distribución espacial de MS de acuerdo a la similaridad con tres patrones de distribución referentes: agrupada, aleatoria o dispersa, dónde la existencia de una distribución agrupada infiere una fuerte dependencia espacial positiva, una distribución aleatoria no presenta dependencia espacial y una distribución dispersa implica una fuerte distribución espacial negativa (Collet et al., 2016). La distribución espacial de incidencia de MS se presentó mediante mapas asociados a la secuencia temporal del fenómeno estocástico. La relación de dependencia o autocorrelación espacial se evaluó a través del índice de Moran (Dormann et al., 2007), respecto a la aparición de nuevos sitios de infección de MS y los precedentes, para las 13 semanas monitoreadas.
RESULTADOS
El comportamiento espacial de incidencia de PNF se analizó mediante semivariogramas empíricos (o experimentales), obtenidos a partir del conjunto de puntos medidos (Figura 1). Los semivariogramas se usaron como herramientas en la detección de continuidad espacial y de la estructura del proceso aleatorio, ya que la semivarianza empírica proporcionó un estimado de la semivarianza teórica, que caracterizó la estructura espacial del proceso. La variable incidencia de PNF presentó estructura de dependencia espacial de acuerdo a los semivariogramas empíricos obtenidos debido a que la semivarianza no fue constante en función de la distancia en ningún caso (Fig. 1). Posteriormente, con el ajuste de estos semivariogramas a modelos matemáticos teóricos se logró cuantificar el grado y escala de variación espacial, permitiendo dilucidar aspectos biológicos importantes asociados a la epidemia de PNF. Entre estos, se logró inferir la capacidad máxima de dispersión del inoculo de infección en campo a través de los valores de los rangos obtenidos en m (Tabla 1), bajo las condiciones agronómicas dadas en este estudio. Mediante el ajuste se obtuvieron los modelos de "potencia" (Pow), "esférico" (Sph) y "efecto sinehole" (She) para las 13 semanas monitoreadas. Este ajuste permitió extraer parámetros estructurales (Tabla 1) usados posteriormente para la interpolación por kriging.
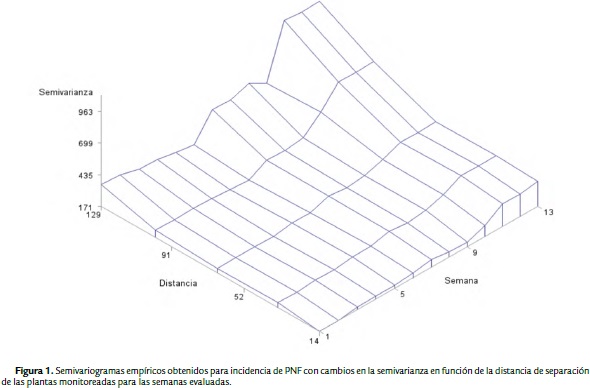
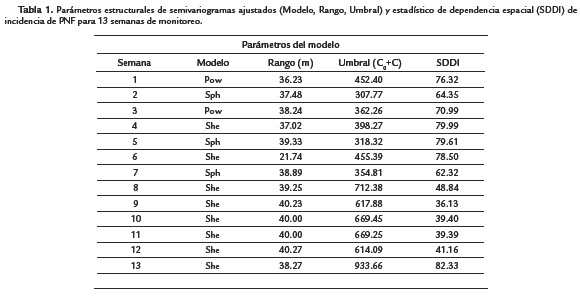
En general, en el modelo Pow la semivarianza del proceso estocástico se incrementó constantemente con la distancia, lo que indicó la existencia de dependencia espacial de incidencia de la enfermedad dentro de los rangos de distancia obtenidos (Tabla 1) y condicionada por los focos de infección presentes. En el modelo Sph la semivarianza alcanzó el valor del umbral en la escala, medida de la variación espacial que correspondió a la varianza de la población, a partir de la cual las medidas de incidencia de PNF son espacialmente independientes unas de otras. En el modelo She la semivarianza se incrementó con la distancia, alcanzando el umbral a una distancia promedio de 37,09 m y oscilando alrededor del valor del umbral con una amplitud que disminuyó a medida que se movió a mayores valores del rango.
Se corroboró la existencia de correlación espacial entre plantas de cacao infectadas para el tiempo de evaluación empleado. Los valores de rango para los cuales existió una dependencia espacial moderada (25 % < SDDI < 75 %) de incidencia de PNF, se encontraron en un radio de distancia entre los 36,23 y 40,27 m para la mayoría de las semanas evaluadas. A distancias menores, alrededor de 21,74 m para el modelo ajustado "She" en la semana seis, la dependencia espacial fue enmascarada debido a la presencia intercalada de clones con características de resistencia a la enfermedad. Por el contrario, para rangos mayores en los semivariogramas obtenidos, la dependencia espacial tendió a ser más fuerte (Tabla 1).
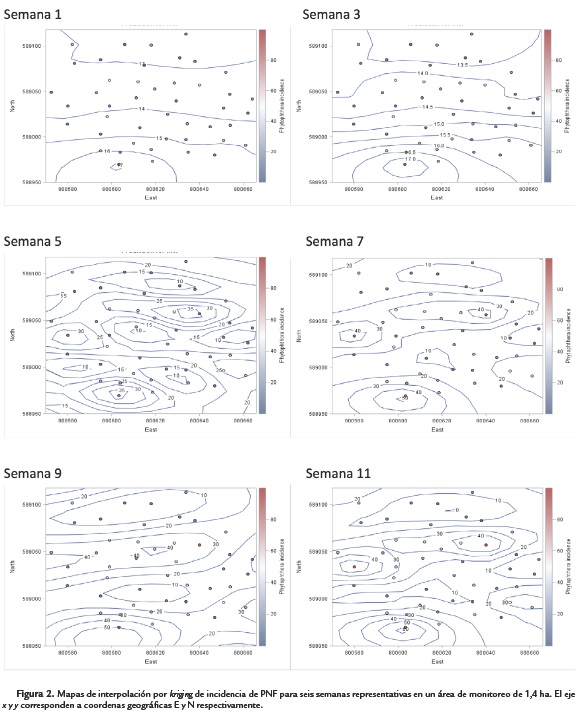
A partir de los parámetros estructurales de los semivariogramas, se construyeron representaciones gráficas bidimensionales de la estimación kriging que identificaron la distribución espacial de la variable incidencia de PNF en campo (Fig. 2). Se corroboró la aparición de múltiples focos de infección y el incremento de los valores de incidencia en sus zonas de influencia en el lote a través de las semanas evaluadas. Valores de incidencia de la enfermedad para diferentes puntos no muestreados, se predijeron a partir de puntos muestreados en el lote. Estos valores se ilustran por medio de diferentes "niveles" de incidencia delimitados por contornos, para seis semanas representativas de evaluación (Fig. 2).
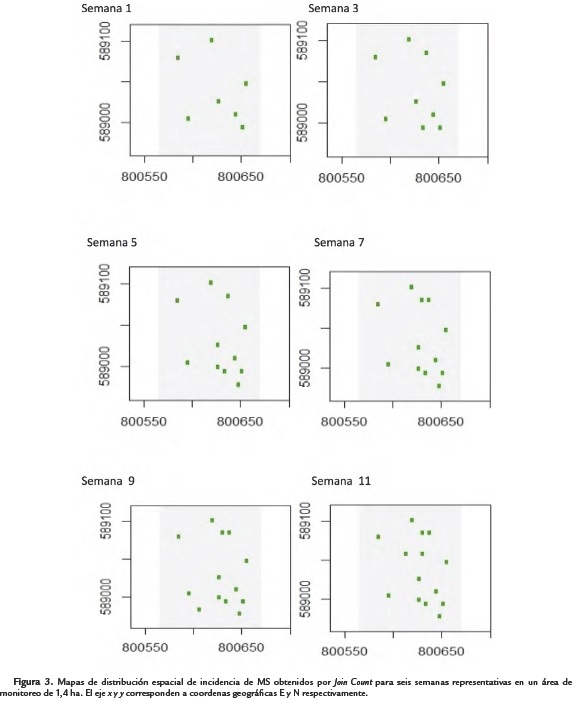
Por otro lado, en el análisis del comportamiento espacial de incidencia de MS, se identificaron sitios en el lote propios a la aparición de focos de infección de la enfermedad. Estos correspondieron con la ubicación de árboles de cacao que reportaron características de susceptibilidad para esta enfermedad. En general, se evidenció la aparición de nuevos sitios de infección en el tiempo (Fig. 3).
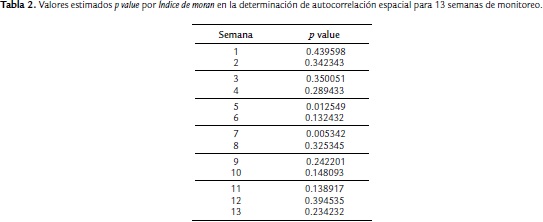
La autocorrelación espacial, a través de los valores estimados de p value de la prueba o Índice de Moran (Tabla 2), se evidenció para las semanas cinco y siete (p < 0,05). Para las semanas restantes no se rechazó la hipótesis asociada al hecho de que la incidencia de MS se distribuyó de forma aleatoria entre las plantas del área de estudio. De acuerdo a esto, los procesos espaciales que promueven el patrón de infección observados constituyen una dispersión aleatoria excepto para las semanas cinco y siete (p < 0,05), dónde existe correlación espacial del proceso de infección de esta enfermedad.
DISCUSIÓN
Se evidenció un progreso de la incidencia de las enfermedades en el tiempo con la aparición de nuevos focos de infección a través de las semanas de evaluación (Fig. 2 y Fig. 3). El estudio de semivariogramas usando un enfoque geoestadístico permitió identificar focos de infección de PNF y MS durante las 13 semanas de monitoreo. Los rangos a los cuales se encontró dependencia espacial de los datos para PNF, indicaron focos de infección distanciados entre 21,74 y 40,27 m para las semanas evaluadas (Tabla 1 y 2); distancias que correspondieron a una mayor amplitud comparadas con las reportadas por Ten Hoopen et al. (2009) para la misma enfermedad en un cultivo monoclonal de cacao en África, las cuales oscilaron entre los 3,3 y 6,6 m. Esto puede explicarse por las características propias de alternabilidad de clones en el cultivo monitoreado, con distintas características de resistencia, que limitó la expresión de nuevos focos de infección y dispersión de la epidemia a manera de barrera viva, en lugares del espacio con presencia de clones resistentes a Phytophthora spp.
Los mapas de interpolación por kriging permitieron predecir la distribución, cuantificación y evolución de zonas de influencia bajo niveles específicos de incidencia de PNF, dados por los diferentes contornos con valores numéricos definidos. (Fig. 2). Se evidenciaron variaciones en los niveles de incidencia y su área de influencia en el tiempo, con una tasa de cambio promedio de 11,03 % mensual en regiones del lote de alta incidencia y de 5,14 % mensual en zonas de menor incidencia de la enfermedad.
En general, la aparición de estos focos de infección de PNF y MS en sitios específicos correspondió a la ubicación de clones con características susceptibles, ilustrando la dinámica epidemiológica de las enfermedades y la relación de especificidad patógeno-hospedante. En plantas del clon CCN51, se corroboraron las características de susceptibilidad a Phytophthora spp. (Arciniegas y Phillips, 2006) y resistencia a Monilophthora roreri (Corpoica, 2004) y de resistencia a Phytophthora spp. del clon ICS95 (Arciniegas y Phillips, 2006). Sin embargo, no se evidenció resistencia a Monilophthora roreri en el material ICS95 (Corpoica, 2004;) en el lote experimental dónde se presentó incidencia de MS.
De acuerdo a esto último, aunque existió presencia de MS en plantas ICS95, ésta enfermedad no se expresó con mayor frecuencia debido a que los valores de incidencia en todas las plantas fueron menores (media 8 %) comparados con los mayores valores de incidencia para PNF (media 35 %). Además, la resistencia reportada en ICS95 frente a Monilophthora roreri se referencia como resistencia "moderada" (Corpoica, 2004) a significativamente alta, ante aislados colombianos del patógeno (Phillips-Mora et al., 2005).
Así mismo, otras características relacionadas con el sistema productivo tales como edad del cultivo y cantidad de frutos por planta, pueden explicar también la ocurrencia de condiciones conductivas para la expresión de las enfermedades evaluadas. Esto debido a que se ha detectado que estas variables se relacionan significativamente con las pérdidas provocadas por PNF y MS en plantaciones de edades entre los 4,4 a 8,7 años (Ramírez, 2016). Rango de edad que correspondió a todos las plantas del lote monitoreado y, en su mayoría, con significativa producción de frutos.
Por otra parte, las condiciones agroclimáticas presentadas en la zona de estudio también pudieron influir en la dinámica de dispersión de las enfermedades. La alta humedad relativa y precipitaciones constantes presentadas, para los meses comprendidos entre agosto y octubre (dentro del régimen de lluvias unimodal en la región) en los cuales se llevó a cabo el experimento, pueden explicar la rápida aparición de focos de infección de PNF durante el tiempo de evaluación. Esto en concordancia con Wood (1974) quien reporta que períodos de alta humedad relativa son requeridos para la rápida dispersión de la enfermedad, y con Ramírez (2016) quién evidenció la mayor relación de ocurrencia de pérdidas en producción de cacao por efecto de incrementos en la precipitación y humedad relativa.
De acuerdo a lo anterior, la humedad relativa se ha visto como el factor climático más determinante para explicar la alta incidencia en regiones de alta humedad relativa como Nigeria y la ausencia casi completa de la enfermedad en zonas de baja humedad relativa en zonas como Malasia (Cabi, 2015). De igual forma, la precipitación y liberación de zoosporas en el agua superficial del suelo, desde donde son esparcidas a los frutos de las plantas por salpicadura de agua lluvia (McMahon y Purwantara, 2004), es un mecanismo de dispersión del inóculo de infección eficaz que puede explicar la mayor incidencia de PNF con respecto a MS. También pudieron existir otros mecanismos de dispersión de la enfermedad a través del movimiento de inóculo desde el suelo hasta el dosel por insectos como la "hormiga negra pequeña" Crematoga sterstriatula (Evans, 1973) que requieren de evaluación detallada. Sin embargo, se ha reportado que el inóculo potencial del suelo disminuye rápidamente con el tiempo fuera de los períodos epidémicos, principalmente por su baja capacidad saprofítica y no patogénica en las raíces de cacao. Aún así, se conoce que puede sobrevivir entre la corteza en los tallos de las plantas por varios meses (Despreaux et al., 1987).
La rápida aparición de focos de infección de PNF en el tiempo, puede explicarse por las condiciones propias de alta humedad relativa en la región, para los meses comprendidos entre abril y noviembre (régimen de lluvias unimodal), período dentro del cual se llevó a cabo el experimento. Esto en concordancia con Wood (1974) quien reporta que períodos de humedad relativa son requeridos para la rápida dispersión de la enfermedad. Deberdt et al. (2008) también asocian la ocurrencia de precipitaciones y alta humedad relativa como condiciones ambientales que favorecen el desarrollo de la epidemia y las especies de Phytophthora implicadas. Es así como, la humedad relativa se ha visto como el factor climático más determinante para explicar la alta incidencia en regiones de alta humedad relativa como Nigeria y la ausencia casi completa de la enfermedad en zonas de baja humedad relativa en zonas como Malasia (Cabi, 2015).
Por otro lado, se ha reportado que la formación, liberación y dispersión de esporas de Moniliophthora roreri no requiere de alta humedad y su dispersión natural ocurre principalmente por viento (Cabi, 2015). Condición que puede explicar, en parte, la menor frecuencia de aparición de focos de infección en el tiempo, con valores más bajos de incidencia de la enfermedad MS con respecto a PNF. Sin embargo, el potencial para la dispersión por viento, es limitado a poca distancia a partir de una sola fuente de inóculo debido al peso de las esporas y su susceptibilidad (Phillips-Mora et al., 2007). Además, Ramírez (2016) menciona otras variables de importancia asociadas a la ocurrencia de MS como edad del cultivo y el total de frutos presentes que deben ser tenidas en cuenta.
Lo anterior también puede ser explicado porque, la alta incidencia de nuevos focos de infección de MS ocurre cuando coinciden las condiciones de aparición de un gran número de frutos, un clima de lluvia cálida y la presencia de inóculo en campo (frutos infectados) (Cabi, 2015). Además, si las esporas son liberadas al suelo, su desplazamiento a largas distancias no es significativo y generalmente los cambios en la turgencia en pocos días y la invasión por otros microorganismos, inmoviliza las esporas (Gonzáles et al., 1981). De tal forma que, el principal vector de transmisión viene dado por la actividad humana y de desplazamientos en las plantaciones (Phillips-Mora et al., 2007).
Los semivariogramas experimentales obtenidos y su ajuste a modelos teóricos demostraron la existencia de autocorrelación espacial de la variable incidencia de PNF, lo que sugiere que los métodos geoestadísticos pueden ser una herramienta útil en la modelación e interpretación de epidemiologías en el sistema productivo de cacao en el trópico húmedo. Los semivariogramas experimentales reflejaron las distancias máximas y la manera en que un punto tiene influencia sobre otro punto en función de la distancia. Esta información base se ajustó, a semivariogramas teóricos para modelar matemáticamente el proceso estocástico debido a la imposibilidad de utilizar de forma directa los semivariogramas experimentales en el método de interpolación kriging. Por otro lado, el Join Count como metodología de análisis espacial, permitió evaluar presencia/ausencia de MS en las plantas monitoreadas y el estadístico de prueba Indice de Moran fue útil para probar autocorrelación espacial entre patrones de puntos o sitios de infección de la enfermedad.
CONCLUSIONES
Las condiciones experimentales dadas en el lote de cultivo y el enfoque de análisis espacial empleado, contribuyó al entendimiento de la dinámica espacial de dos enfermedades limitantes en tres clones de cacao en la Orinoquía Colombiana. Estos análisis prueban su gran importancia en el diseño a futuro, de estrategias de manejo de epidemias de gran incidencia fitopatológica, a través por ejemplo, del diseño espacial de plantaciones en donde materiales vegetales con grados de tolerancia a enfermedades, puedan ser ubicados estratégicamente, sin sacrificar productividad ni calidad.
La alta humedad relativa, característica de la zona donde fue establecido el experimento, parece ser el factor climático más influyente en el desarrollo de las epidemias de PNF y MS. Esta condición de humedad relativa y las características propias de los mecanismos de dispersión de estas enfermedades, se relacionaron con una mayor incidencia de PNF y menor de MS. La dinámica del progreso de las enfermedades PNF y MS a través de la aparición de focos de infección en sitios específicos donde se encontraban clones con características de susceptibilidad definidas, reflejó condiciones conductivas de la aparición de la enfermedad bajo las condiciones experimentales dadas. Sin embargo, es necesario realizar estudios experimentales con tiempos de evaluación más robustos, en condiciones de época lluviosa y seca, para estudiar la dinámica epidemiológica más precisa de estas enfermedades, teniendo en cuenta las características propias del triángulo de enfermedad: hospedante susceptible-patógeno virulento-ambiente propicio.
De acuerdo a los resultados obtenidos en este estudio bajo las condiciones experimentales dadas, y en concordancia con lo reportado con Ramírez (2016) en plantaciones de Antioquia-Colombia, se infiere que la PNF fue la epidemia más predominante en el lote de cultivo ubicado en El Yopal (Casanare)-Colombia comparada con MS, enfermedad que ha sido referenciada previamente como la de mayor impacto negativo en el país.
AGRADECIMIENTOS
A COLCIENCIAS en el marco del proyecto "Análisis de tecnologías novedosas para el cultivo del cacao en el departamento de Casanare-Colombia" por la financiación de este trabajo. Al profesor Edgar Benitez por su importante contribución y aportes en el planteamiento inicial de análisis de este estudio. Al proyecto Utopía de la Universidad de la Salle (Yopal- Casanare) y sus estudiantes de tercer año en el 2015 por su apoyo en campo.
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
REFERENCIAS
Alves MC, Pozza EA, Machado JC, Araújo DV, Talamini V, Oliveira MS. Geostatistics as methodology to study the space-time dynamics of diseases transmitted by seed-borne Colletotrichum spp. Fitopatol Bras. 2006;31:557-563. Doi:10.1590/S0100-41582006000600004. [ Links ]
Alves MC, Pozza EA. Indicator kriging modeling epidemiology of common bean anthracnose. Appl Geomat. 2010;2:65-72. Doi:10.1007/s12518-010-0021-1. [ Links ]
Agrios GN. Plant Pathology. 5 ed. San Diego: Academic Press; 2004. p. 126-138. [ Links ]
Aime MC, Phillips-Mora W. The causal agents of witches' broom and frosty pod rot of cacao (Theobroma cacao L.) form a new lineage of Marasmiaceae. Mycol. 2005;97(5):1012-1022. Doi:10.3852/mycologia.97.5.1012. [ Links ]
Arciniegas A, Phillips MW. Caracterización de genotipos superiores de cacao seleccionados por el programa de mejoramiento genético del CATIE por su rendimiento y/o resistencia a moniliasis. En: Libro de resúmenes, International Cocoa Research Conference: proceedings actes. San José, Costa Rica; 2006. p. 21-25. [ Links ]
Burrough PA, McDonnell RA. Principles of geographical information systems. New York: Oxford University Press; 1998. p. 97-124. [ Links ]
Cabi. Moniliophthora roreri (frosty pod rot). In: Invasive Species Compendium. Cent for Agricul Bioscien Inter. 2015. Available in: http://www.cabi.org/isc/datasheet/40979. [Cited: 13 Jun 2016] [ Links ].
Cabi. Phytophthora megakarya (black pod of cocoa). In: Invasive Species Compendium. Cent for Agricul Bioscien Inter. 2016. Available in: http://www.cabi.org/isc/datasheet/40979. [Cited: 13 Jun 2016] [ Links ].
Campbell CL, Madden LV. Introduction to plant disease epidemiology. New York: Wiley Editorial; 1990. p. 53-90. [ Links ]
Collet C, Schneuwly D, Caloz R. Discrete spatial distributions 2016. Available in: http://www.gitta.info/DiscrSpatVari/en/text/DiscrSpatVari.pdf. [Cited: 03 Ago 2016] [ Links ].
Corpoica. Caracterización de clones de cacao por respuesta a monilia (Moniliophthora roreri) en Santander. Corp Col de Invest Agro. 2004. Available in: http://conectarural.org/sitio/sites/default/files/documentos/CARACTERIZACION%20DE%20CLONES%20DE%20CACAO%20POR%20RESPUESTA%20A%20MONILIA%20%20EN%20SANTANDER. [Cited: 19 Sep 2016] [ Links ].
Correa JS, Castro S, Coy J. Estado de la moniliasis del cacao causada por Moniliophthora roreri en Colombia. Acta Agron. 2014;63(4):388-399. Doi: 10.15446/acag.v63n4.42747. [ Links ]
Deberdt PM, Fegue CV, Tondje PR, Bon MC, Ducamp M, Hurard C, et al. Impact of environmental factors, chemical fungicide and biological control on cacao pod production dynamics and black pod disease (Phytophthora megakarya) in Cameroon. Biological Control. 2008;44:149-159. Doi:10.1016/j.biocontrol.2007.10.026. [ Links ]
Despreaux D, Cambrony D, Clement D, Nyasse S, Partiot M. Study of cocoa black pod in Cameroon: description of new control methods. In: Proceedings of the Tenth International Cocoa Research Conference. Santo Domingo, República Dominicana; 1987. p. 407-412. [ Links ]
Dormann CF, McPherson JM, Araújo MB, Bivand R, Bolliger J, Carl G, Davies RG, et al. Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecograp. 2007;30(5):609-628. Doi:10.1111/j.2007.0906-7590.05171.x. [ Links ]
Evans HC, Commonwealth Mycological Institute (Great Britain). Pod rot of cacao caused by Moniliophthora (Monilia) roreri. Commonwealth Mycological Institute, Kew, Surrey, 1981. 44 p. [ Links ]
Evans HC. New developments in black pod epidemiology. Cocoa Research Institute. Coco Grow Bullet. 1973;20:10-16. [ Links ]
Florez E, Corredor D. Análisis espacial de las poblaciones de Frankliniella occidentalis (Pergande) en un cultivo de fresa bajo cubierta, como soporte en las decisiones de manejo integrado de plagas. Agron Colom. 2000;17:25-42. [ Links ]
González L. Efecto de las fuentes de inóculo sobre las posibilidades de combate de la moniliasis del cacao. En: primeras jornadas de investigación. UCR. San José, Costa Rica; 1981. p. 228-229. [ Links ]
ICA. El ICA apoya Plan Nacional de Renovación de Cacao. Instituto Colombiano Agropecuario, ICA Comunica. 2013. Available in: http://www.ica.gov.co/Periodico-Virtual/Prensa/2013/El-ICA-apoya-Plan-Nacional-de-Renovacion-de-Cacao.aspx. [Cited: 25 Ene 2016] [ Links ].
Jaimes Y, Aránzazu F. Manejo de las enfermedades del cacao (Theobroma cacao L.) en Colombia, con énfasis en Monilia (Moniliophthora roreri). In: Hoyos L.M., editor. Corporación Colombiana de Investigación Agropecuaria, Corpoica, Colombia. 2010; 90 p. [ Links ]
Jaimes Y, Aranzazu F, Rodríguez E, Martínez N. Behavior of introduced regional clones of Theobroma cacao toward the infection Moniliophthora roreri in three different regions of Colombia. Agron Colom. 2011;29(1):361-371. [ Links ]
Madden LV, Hughes G. Plant disease incidence: Distributions, heterogeneity and temporal analysis. Annu Rev ofPhytopath. 1995;33:529-564. Doi:10.1146/annurev.py.33.090195.002525. [ Links ]
Madden LV, Hughes G. Sampling for plant disease incidence. Phytopath. 1999;89(11):1088-1101. Doi:10.1094/PHYTO.1999.89.11.1088. [ Links ]
McMahon P, Purwantara A. Phytophthora on cocoa. In: Drenth A, Guest D, editors. Diversity and Management of Phytophthora in Southeast Asia, 1ra. Ed. Australian Centre for International Agricultural Research. 2004;114:104-115. [ Links ]
Merritt RN, Orum TV, Garcia RJ. Applications of Geographic Information Systems and Geostatistics in Plant Disease Epidemiology and Management. Plan Diseas. 1999;83(4):308-319. Doi:10.1094/PDIS.1999.83.4.308. [ Links ]
Moral F. Aplicación de la geoestadística en las ciencias ambientales. Ecosist. 2004;13:78-86. [ Links ]
Phillips-Mora W, Castillo JK, Rodríguez E, Wilkinson MJ. Evaluation of cacao (Theobroma cacao) clones against seven Colombian isolates of Moniliophthora roreri from four pathogen genetic groups. Plant Pathol. 2005;54(4):483-490. Doi:10.1111/j.1365-3059.2005.01210.x. [ Links ]
Phillips-Mora W, Aime MC, Wilkinson MJ. Biodiversity and biogeography of the cacao (Theobroma cacao) pathogen Moniliopthora roreri in tropical America. Plant Pathol. 2007;56:911-922. Doi:10.1111/j.1365-3059.2007.01646.x. [ Links ]
Phillips-Mora W, Wilkinson MJ. Frosty pod of cacao: A disease with a limited geographic range but unlimited potential of damage. Phytopathol. 2007;97:1644-1647. Doi:10.1094/PHYTO-97-12-1644. [ Links ]
Pizzato JA, Araújo D, Galvanin E, Junior JR, Matos A, Vecchi M, Zavislak F. Geostatistics as a Methodology for Studying the Spatiotemporal Dynamics of Ramularia areola in Cotton Crops. Amer Jour Plant Scien. 2014;5:2472-2479. Doi: 10.4236/ajps.2014.515262. [ Links ]
Plantegenest M, Le May C, Fabre F. Landscape epidemiology of plant diseases. JR Soc Inter. 2007;16(4):963-972. Doi:10.1098/rsif.2007.1114. [ Links ]
Ramírez JG. Pérdidas económicas asociadas a la pudrición de la mazorca del cacao causada por Phytophthora spp. y Moniliophthora roreri (Cif y Par) Evans et al., en la hacienda Theobroma, Colombia. Rev de Prot Veg. 2016;31(1):42-49. [ Links ]
Rodríguez E, Vera A. Identification y manejo de la pudrición parda de la mazorca (Phytophthora sp.) en cacao. Corpoica: Bogotá, Colombia. 2015. 60. p. [ Links ]
Rossi RE, Mulla DJ, Journel AG, Franz EH. Geostatistical Tools for modeling and interpreting ecological spatial dependence. Ecol Monogra. 1992;62(2):277-314. Doi:10.2307/2937096. [ Links ]
Ten Hoopen M, Sounigo O, Babin R, Dikwe G, Cilas C. Spatial and temporal analysis of a Phytophthora megakarya epidemic in a plantation in the centre region of Cameroon. Proceedings of the 16th International Cocoa Research Conference. 2009. 683-687 p. [ Links ]
van Maanen A, Xu X M. Modelling plant disease epidemics. Europ J Plant Pathol. 2003;109(7):669-682. Doi:10.1023/A:1026018005613. [ Links ]

