











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
En respuesta a patógenos, las plantas han desarrollado un sofisticado mecanismo de acción, el cual depende en primera medida de la capacidad de reconocer moléculas propias del patógeno y extrañas para la planta, tanto en la resistencia cuantitativa como cualitativa (Boller y He, 2009; Vásquez et al., 2018). A nivel de la membrana plasmática, el reconocimiento de los patógenos depende de los receptores de reconocimiento de patrones (PRRs) los cuales reconocen patrones moleculares asociados a microbios (MAMPs). Por otro lado, a nivel intracelular el reconocimiento depende de las proteínas codificadas por los genes de resistencia R, las cuales reconocen proteínas efectoras inyectadas por los patógenos (Monaghan y Zipfel, 2012; Jones et al., 2016). Una vez ocurre el reconocimiento del patógeno por parte de la planta se desencadena una serie de respuestas de defensa. Estas defensas incluyen el fortalecimiento de la pared celular a través de la síntesis de calosa y lignina (Hauck et al., 2003), la producción de metabolitos secundarios antimicrobianos como fitoalexinas (Almagro et al., 2009) y la activación de las cascadas de señalización por proteínas kinasas activadas por mitógenos (MAPK) (Meng y Zhang, 2013), entre otras. Todas estas respuestas culminan con la inducción de expresión de genes dentro de los cuales se encuentran aquellos que codifican para proteínas relacionas con patogenicidad (PR) (van Loon, et al., 2006).
Las proteínas PR están asociadas con la defensa de la planta, especialmente en interacciones incompatibles, en las cuales se impide el progreso del patógeno. Estas proteínas no poseen un alto grado de especificidad en la respuesta a patógenos y son coordinadas a nivel transcripcional (van Loon et al., 1999). Clásicamente, los genes PR se han definido como aquellos que son inducidos únicamente en respuesta a patógenos y no bajo ningún otro tipo de estímulo o respuesta, sin embargo, esta definición es problemática porque un PR en un patosistema puede no serlo en otro (van Loon et al., 2006). Aunque muchas de las proteínas PR poseen una actividad antimicrobiana in vitro y su acumulación está asociada con una respuesta de resistencia de la planta, la función de estas proteínas in vivo es poco conocida (Sels et al., 2008). Basada en su similitud de secuencia y actividad, las proteínas PR se han clasificado en 17 familias denominadas PR1 a PR17 (van Loon et al., 2006). Estas proteínas poseen una masa molecular relativamente baja e incluyen, por ejemplo, proteínas con actividad tipo quitinasa (PR3, 4, 8 y 11), glucanasa (PR2) y ribonucleasa (PR10). Una de las PR más empleadas como marcador de activación de la resistencia es PR1 cuya función o actividad aún se desconoce (van Loon et al., 2006). Adicionalmente, se conoce que las proteínas PR son activadas por varias hormonas tales como el ácido salicílico, ácido jasmónico y etileno (Sels et al., 2008).
La yuca es una de las principales fuentes de alimentación en África, Asia y Latinoamérica (FAO, 2017). Sin embargo, su producción se ve fuertemente afectada por la bacteriosis vascular, causada por Xanthomonas axonopodis pv. manihotis (Xam). En los últimos años se han empleado diferentes estrategias para comprender mejor los mecanismos celulares y moleculares de la interacción yuca-Xam. Gracias a estudios histológicos (Boher et al., 1995; Kpémoua et al., 1996) y al mapeo de loci de rasgos cuantitativos (QTL) (Jorge et al., 2000; Soto et al., 2016), se ha podido establecer que la resistencia a Xam es de tipo cuantitativa. El nivel de resistencia entre las diferentes variedades es muy variable, en tanto que la velocidad de la respuesta es genotipo-dependiente. Se ha observado que las respuestas son más rápidas e intensas en variedades resistentes comparadas con las susceptibles (Kpémoua et al., 1996; López et al., 2005; Sandino et al., 2015). Adicionalmente, se han realizado varios estudios con el fin de identificar proteínas involucradas en el reconocimiento del patógeno, así como el estudio del cambio de la expresión génica global durante la interacción yuca-Xam, tanto en variedades resistentes como susceptibles (Santaella et al., 2004; López et al., 2005; Muñoz et al., 2015). Estos estudios han evidenciado el aumento en los niveles de expresión de posibles genes de defensa presentes en yuca y que han sido previamente descritos en otros patosistemas. Estos incluyen genes involucrados en el estallido oxidativo (ascorbato peroxidasa, glutatión-S-transferasa), reconocimiento de patógenos potenciales (proteínas con dominios LRR) y reguladores de transcripción (factores de transcripción de tipo WRKY). También se han identificado como diferencialmente expresados genes que codifican para una quitinasa y osmotina, los cuales basados en su similitud y en su potencial función se acuñaron como putativos genes PR (López et al., 2005). Sin embargo, hasta hace poco tiempo no se había establecido con certeza cuáles son los genes de tipo PR que se inducen en respuesta a Xam y si esta expresión es diferencial entre variedades resistentes y susceptibles. Un primer estudio que aborda esta problemática fue reportado por Yoodee et al. (2018) quienes evaluaron genes que codifican para proteínas R, RLKs, WRKY y PRs en dos variedades durante la infección con una cepa de Xam empleando plantas in vitro sin considerar la evaluación sobre plantas provenientes de estacas maduras. Tres genes de tipo WRKY, PR1 y PDF2.2 fueron inducidos y su expresión se incrementó con un previo tratamiento con hormonas vegetales (Yoodee et al., 2018). Este tipo de conocimiento es fundamental no sólo para evaluar si existen respuestas moleculares diferenciales entre variedades resistentes y susceptibles sino también para contar con genes marcadores que permitan determinar rápidamente si se están activando procesos moleculares inducidos en respuesta a una infección. Con este tipo de conocimiento se podría realizar una mejor caracterización de variedades resistentes y compararlas con susceptibles, y determinar molecularmente las respuestas que se establecen en yuca cuando se emplean aproximaciones en las que se busca generar resistencia a diversos patógenos.
Con el fin de identificar genes PR en yuca, en este estudio se empleó en primera instancia una aproximación bioinformática para identificar putativos genes PR en el genoma de yuca, posteriormente un set de ellos fue seleccionado para evaluar su expresión mediante RT-PCR y qRT-PCR en plantas de yuca adultas de variedades resistentes y susceptibles tras la inoculación con Xam en condiciones de invernadero.
MATERIALES Y MÉTODOS
Búsqueda de PR candidatos y diseño de cebadores
Para identificar el repertorio de proteínas PR en yuca, en primera instancia, se llevó a cabo una búsqueda en la base de datos Uniprot empleando como motor de búsqueda palabras conteniendo los nombres de cada uno de los PR: "PR1", "PR2", hasta "PR17". También se emplearon palabras que abarcaran su función molecular tales como: proteínas relacionadas con patogénesis (pathogenesis-related protein) y la actividad asociada al tipo de familia.
A partir de las secuencias seleccionadas, se realizó un alineamiento tipo BLASTP de cada una contra el proteoma de yuca y se seleccionaron los cinco mejores resultados a condición de que tuvieran un porcentaje de similitud mayor al 60 % para cada uno de los PR de referencia de Uniprot. A partir de las secuencias codificantes (CDS, coding sequence) de los PR candidatos obtenidos de yuca, se diseñaron los cebadores empleando el programa Primer3 (Untergasse et al., 2012).
Material vegetal e inoculación
Plantas de las variedades cv.60444 y SG107-35 se crecieron durante dos meses en el invernadero de Biología de la Universidad Nacional de Colombia, bajo las siguientes condiciones: temperatura de 28°C/21°C (día/noche), humedad relativa de 80 %, con un fotoperiodo de 12 horas luz y 12 horas oscuridad.
Las bacterias se crecieron por agotamiento a partir del stock almacenado a -80 °C del cepario del laboratorio del grupo Manihot Biotec de la Universidad Nacional, en medio LPGA (5 g extracto de levadura, 5 g peptona, 5 g glucosa y 15 g agar por 1 L de agua destilada). Las bacterias se crecieron a 28 °C durante dos días. Tras obtener colonias aisladas, se tomó una de ellas y se realizó el cultivo en 3 mL de medio LPG a 28 °C a 250 rpm durante 12 h. Se centrifugó 1 mL del cultivo a 6000 rpm durante 4 min, se descartó el sobrenadante, y posteriormente se resuspendió el precipitado en 1 mL de MgCl. Se ajustó a una DO de 0,02 que corresponde a 107 UFC/mL (unidades formadoras de colonia). La concentración de bacterias empleada se verificó por diluciones seriadas.
La inoculación se realizó en el tallo mediante punción entre la segunda y tercera hoja totalmente extendida con un palillo de madera estéril. El palillo fue removido y se reemplazó por una punta pequeña estéril con 10 uL de la solución bacteriana de Xam686. Para las plantas control se realizó el mismo procedimiento, pero con una solución de MgCl2 10 mM. Se tomó la región del tallo que comprendía 1.5 cm hacia arriba (ápice) y 1.5 cm hacia abajo desde el punto de inoculación. Para cada tratamiento se colectó el tallo en nitrógeno líquido. Los tiempos de toma de muestra fueron: 0 y 24 horas post-inoculación (hpi) y 3, 5 y 7 días post-inoculación (dpi). Se empleó un diseño completamente aleatorizado para la distribución de las plantas en el invernadero.
Extracción de ARN, síntesis de ADNc y RT-PCR
El material vegetal se maceró en un mortero utilizando nitrógeno líquido. El buffer de extracción utilizado contenía acetato de sodio 100 mM, EDTA pH 7.5 1 mM y SDS 4 % (pH 5,2). Posteriormente se purificó con fenol ácido (pH 4,5): cloroformo: alcohol isoamílico 24:24:1. La fenolización se repitió una vez y se realizó la extracción con cloroformo:alcohol isoamílico 24:1, dos veces. Finalmente, se precipitó con isopropanol. El ARN se lavó con etanol al 70 % y se resuspendió en agua ultra-pura (libre de nucleasas). A partir del ARN extraído manualmente, se realizó una purificación empleando el Invitrap ® Spin plant RNA minikit (STRATEC, Berlín, GER). La integridad del ARN se verificó en gel de agarosa al 1% y se cuantificó con el espectrofotómetro Nanodrop 2000c (ThermoScientific, Waltham, MA, USA) mediante la medición de absorbancia a 260 y 280 nm. Una vez obtenido el ARN, se trató con DNase I (ThermoScientific, Waltham, MA, USA) sobre 600 ng-1000 ng de ARN total, con el fin de remover el ADN genómico y así poder proceder con la síntesis de ADNc. Posteriormente se realizó un PCR del gen tubulina sobre el ARN tratado previamente con DNase I, como control. La síntesis de primera cadena de ADN se realizó con el kit First Strand cDNA Synthesis (ThermoScientific, Waltham, MA, USA). Para verificar la síntesis correcta de ADNc se realizó un RT-PCR con cebadores diseñados a partir del gen tubulina, el cual se expresa constitutivamente (Tabla suplementaria 1).
qRT-PCR
Todas las reacciones de qRT-PCR se realizaron con el kit SsoAdvanced Universal Inhibitor SYBR Green Supermix (BioRad) en un termociclador LightCycler® 96 System (Roche Diagnostics GmbH, Indianapolis, IN, USA). Las reacciones se realizaron en un volumen final de 10 uL (0,1 mM de cada primer, 0,1 mM de dNTPs) conteniendo 1 uL de ADNc. Las condiciones de amplificación fueron: denaturación inicial a 95 °C por 10 min, 45 ciclos de 95 °C por 20 s, 60 °C por 20 s y 72 °C por 20 s. Se realizaron las curvas de calibración de los cebadores utilizando cinco diluciones seriadas usando un factor de dilución de 1:10. Para la evaluación de los genes se utilizaron 3 réplicas biológicas por cada una de las dos variedades y tres tiempos evaluados para plantas con tratamiento de inoculación y mock. Los resultados se analizaron con el software LightCycler 96 SW 1.1 (Roche). Para el cálculo de la expresión relativa de los genes de interés se utilizó el gen HK2 (Manes06G114200.1 como gen de referencia (normalizador). Se realizaron dos análisis de expresión relativa empleando el método 2-M Ct method (Livak and Schmittgen, 2001). Para el análisis de expresión en relación al tiempo 0 hpi, se empleó el modelo matemático de la siguiente manera: ACt(60444/SG10735 inoculadas a 3 y 7 dpi) = C t (target gene)-C(reference gene); ACt(60444/ SG10735 inoculadas a 0 hpi) = C(target gene)-C (reference gene); AACt(60444/SG10735)= ' AC(60444/Sg10735 inoculadas a 3 y 7 dpi)-ACt(60444/SG10735 inoculadas a 0 hpi). Por otro lado, para el análisis de expresión en relación a la herida con MgCl2, se empleó el modelo matemático de la siguiente manera: ACt(60444/SG10735 inoculadas a 0, 3 y 7 dpi) = C(target gene)-C t (reference gene); ACt(60444/ SG10735 herida con MgCl2 a 0, 3 y 7 dpi) = Ct( target gene)-C t (reference gene); AACt(60444/SG10735 de la inoculación en relación a la herida)= AC (60444/SG10735 inoculadas a 0, 3 y 7 dpi)-ACt(60444/SG10735 herida con MgCl2 a 0, 3 y 7 dpi).
RESULTADOS
Búsqueda de proteínas de tipo PR y diseño de cebadores para homólogos de PRs en yuca
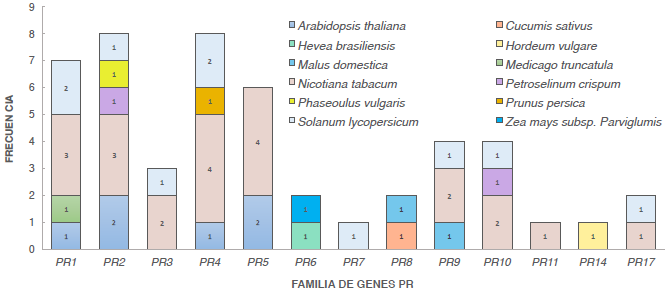
Al realizar la búsqueda en la base de datos de Uniprot empleando como motor de búsqueda el término "PR", su nombre y la función molecular asociada a PRs, se seleccionaron 50 proteínas de este tipo (Tabla suplementaria 1). La mayor cantidad de proteínas PR reportadas provienen de tabaco (Nicotiana tabacum) con 20 secuencias, seguido de tomate (Solanum lycopersicum) con 11 y Arabidopsis thaliana con siete proteínas. Las demás especies tan sólo presentan una proteína reportada, como el caso de cebada (Hordeum vulgare) con el PR14 y la proteína de maíz (Zea mays) de la familia PR6. La familia con mayor número de proteínas reportadas es la PR1, con nueve proteínas, seguido de la familia PR5 con ocho proteínas reportadas y PR2 y PR4 con seis proteínas cada una. Las familias PR3 y PR10 presentaron cuatro proteínas seguido de PR-9 y PR-6. Las demás familias (PR7, 11, 14 y 17) presentaron tan sólo un reporte (Fig. 1).
Las secuencias de estas proteínas PR se descargaron y se procedió a realizar un BLASTP contra el proteoma de yuca v4.1 y así identificar los genes similares en el genoma de yuca. A partir de estos resultados se construyó una lista de los genes PRs por especie y su similar en yuca (Tabla suplementaria 1). Una vez creada la matriz de los diferentes PR reportados y los candidatos similares en yuca, se evaluó si las secuencias de dichos PR candidatos identificados estaban presentes en el microarreglo reportado previamente por López et al. (2005). Sin embargo, no se encontraron en esta colección de marcadores de secuencias expresadas (ESTs)). Se evaluó el nivel de expresión según los datos de RNAseq disponibles (Gomez et al. sometido) de las secuencias similares en yuca para cada una de las 50 proteínas PR reportadas. Para la selección de los genes se tuvo en cuenta aquellos que mostraran un mayor incremento en la expresión, fuese o no este significativo, con respecto al control (plantas no inoculadas) y que representaran miembros de diferentes familias de PRs.
Evaluación de la expresión mediante RT-PCR semicuantitativa
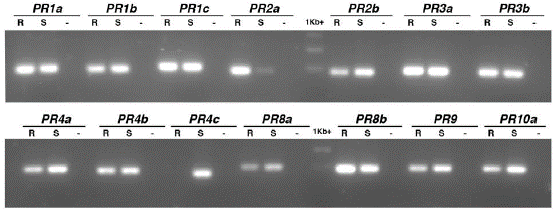
En total, se sintetizaron 15 juegos de cebadores en dirección sentido-antisentido, tres para PR1 y PR4, dos cebadores para PR2, PR3, PR8 y PR10 y uno para PR9 (Tabla 1). Para verificar el correcto diseño de los cebadores y amplificación se realizó un PCR sobre ADN genómico de las variedades cv.60444 y SG107-35 y empleando una temperatura de anillamiento de 52 °C para todos ellos. De los 15 candidatos, 14 amplificaron en las dos variedades (Fig. 2). El único juego de cebadores que no amplificó fue el correspondiente a PR10b.
Tabla 1 Familias de proteínas PR seleccionadas para el estudio en yuca.
| PR | Familia | ID del gen en el genoma de yuca (v.6) |
| PR1a | Pfam:00188 Cysteine-rich secretory protein family | Manes.07G050700 |
| PR1b | Pfam:00188 Cysteine-rich secretory protein family | Manes.07G050400 |
| PR1c | Pfam:00188 Cysteine-rich secretory protein family | Manes.07G050300 |
| MePR1 | Pfam:00188 Cysteine-rich secretory protein family | Manes.06G026900.1 |
| PR2a | Pfam:00332 Glycosyl hydrolases family 17 | Manes.03G202900 |
| PR2b | Pfam:00407 Pathogenesis-related protein Bet v I family | Manes.15G007900 |
| PR3a | Pfam:00182 Chitinase class I | Manes.04G067900 |
| PR3b | Pfam:00182 Chitinase class I | Manes.11G106100 |
| PR4a | Pfam:00187 Chitin recognition protein | Manes.01G064200 |
| PR4b | Pfam:00187 Chitin recognition protein | Manes.08G135100 |
| PR4c | Pfam:00967 Barwin family | Manes.08G135200 |
| PR8a | Pfam:00704 Glycosyl hydrolases family 18 | Manes.06G155500 |
| PR8b | Pfam:00704 Glycosyl hydrolases family 18 | Manes.06G155400 |
| PR9 | Pfam:00141 Peroxidase | Manes.15G104200 |
| PR10a | Pfam:00407 Pathogenesis-related protein Bet v I family | cassava4.1_022277m* |
| PR10b | Pfam:00407 Pathogenesis-related protein Bet v I family | Manes.15G008700 |
* Los genes se identificaron en la versión 4.1 del genoma de yuca y al buscar su correspondencia en la versión actual (v. 6.1) se observó que no lista como anotado (www.phytozome.net).
Una vez verificado que los cebadores podrían amplificar los genes respectivos sobre el ADN genómico de las dos variedades, se evaluó su expresión empleando el ADNc obtenido a partir de las plantas de yuca de las dos variedades y colectado a diferentes tiempos post-inoculación. A partir del ADNc sintetizado se realizó la amplificación del gen constitutivo tubulina (Manes.08G061700) con el fin de verificar la calidad del ADNc. En la figura 3, en el panel superior se observa la amplificación de dicho gen. Cada carril representa el ADNc obtenido de las dos variedades SG107-35 y cv.60444, tratadas con la solución control (MgCl2 10 mM) o con la bacteria (Xam686) y colectado a 0, 1, 3, 5 y 7 dpi. Después de realizar una curva de calibración modificando número de ciclos y cantidades de ADNc en cada muestra se logró obtener un amplicón para tubulina visualmente homogéneo en la mayoría de las muestras, aunque con una pequeña diferencia en algunos tiempos para la variedad susceptible (Fig. 3). Con esta misma cantidad de ADNc se llevaron a cabo las amplificaciones de los genes correspondientes a los PR seleccionados. Dado el gran volumen de trabajo se decidió evaluar inicialmente los genes PR1a, PR1b, PR8a y PR8b en las dos variedades. Además, se incluyó el gen que codifica para la Glutation-S- transferasa el cual ha sido previamente reportado como inducido en una variedad resistente en respuesta a Xam (Lopez et al., 2005). El gen PR8b no mostró expresión en ninguna de las dos variedades, mientras que el gen GST se expresó de manera constitutiva en las dos variedades, incluyendo tanto las plantas no inoculadas (NI) como en las que se trataron con solución de MgCl2 (Fig. 3). Para los genes PR1a y PR1b se observa una leve inducción de la expresión a lo largo del tiempo, particularmente el PR1b muestra una inducción en la variedad resistente y al día 5 post inoculación. El gen PR8a (glicosil-hidrolasa) mostró una expresión desde el tiempo cero tanto en las plantas tratadas con la solución control como las infectadas con Xam en la variedad resistente SG107-35 (Fig. 3). En la variedad susceptible, cv.60444, el gen PR8a se expresa en todos los tratamientos, con una fuerte inducción en la planta inoculada con la solución control y colectada a los 7 dpi. De igual manera, se observó un incremento de la expresión en las plantas inoculadas con Xam y colectadas a los 5 y 7 dpi, siendo mayor en este último tiempo (Fig. 3). Sin embargo, al comparar el perfil de expresión obtenido en las plantas tratadas con MgCl2 se observa también una expresión y/o inducción, sugiriendo que solo el proceso de punción y herida induce la expresión de estos genes, haciendo difícil determinar la contribución cuantitativa real de la inducción provocada por la bacteria misma.
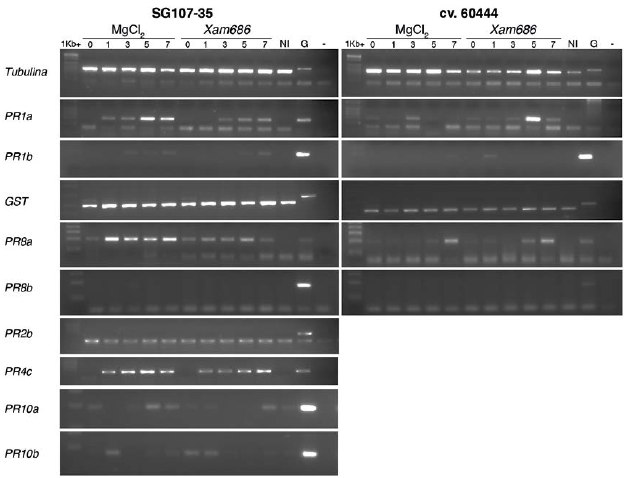
Figura 3 Cinética de expresión de varios genes PR en las variedades resistentes (SG107-35) y susceptible (cv.60444) a lo largo del tiempo tras la inoculación con la bacteria Xam686. NI: No inoculado; G: control de ADN genómico.
La expresión de los genes PR2b, PR4c, PR10a y PR10b se evaluó solamente en la variedad resistente. El gen PR2b no mostró expresión a ningún tiempo ni en ninguna de las condiciones evaluadas. El gen PR4c se indujo tanto para las plantas tratadas con la solución control, como en las infectadas con Xam en todos los tiempos a partir de 1 dpi. Los genes PR10a y PR10b presentaron una amplificación sin una aparente tendencia, incluso hay una leve amplificación en el control absoluto (plantas de la misma edad a las que no se les realizó la inoculación ni con la solución control ni con Xam) (Fig. 3). De esta manera el perfil de los genes PR4c, PR10a y PR10b fue similar al descrito para los genes PR1a, PR1b y PR8, razón por la cual no se pudo determinar la contribución real de la inducción ocasionada por la bacteria misma.
Evaluación de la expresión de genes PR mediante qRT-PCR
Con el fin de lograr una determinación cuantitativa y no basada en la inspección visual de la intensidad de las bandas, se evaluó la expresión de algunos genes PR mediante qRT-PCR (quantitative reverse transcription-PCR). Para ello se determinó el nivel de expresión relativo de cada gen utilizando como gen de referencia Manes06G114200.1 tanto para las muestras provenientes de las plantas inoculadas con Xam como de las plantas tratadas con la solución de MgCl2. De esta manera la relación entre estos dos valores permite determinar el efecto de la inoculación bacteriana sobre la expresión de los genes PR. Para este ensayo se realizó en primera instancia las curvas de eficiencia y la eficacia de los mismos cebadores para los ensayos de QRT. Los cebadores para los genes PR1c, PR3a, PR8b y PR10b, presentaron problemas, baja amplificación (valores de Ct muy altos), presencia de productos inespecíficos y no permitieron ser evaluados en las dos variedades (datos no mostrados). Los genes PR1b y PR2b no mostraron amplificación en ninguna de las dos variedades y en ninguno de los dos tiempos evaluados, confirmando los datos obtenidos mediante RT-PCR semicuantitativa. Los valores de expresión más relevantes se obtuvieron empleando los cebadores para PR1a y PR9. Adicionalmente se incluyó el gen candidato MePR1 (Manes.06G026900.1) recientemente reportado en la literatura (Yoodee et al., 2018). Si bien este gen fue identificado en el primer análisis utilizando la base de datos de Uniprot, no fue considerado inicialmente por no presentar uno de los mejores E- value (Tabla suplementaria 1). Dado el gran número de muestras se decidió evaluar la expresión de estos tres genes en las dos variedades a los tiempos 0, 3 y 7 dpi, tomando de esta forma un tiempo relativamente temprano (3 dpi) y otro tardío (7 dpi) con el objetivo de tener un amplio espectro del perfil de expresión.
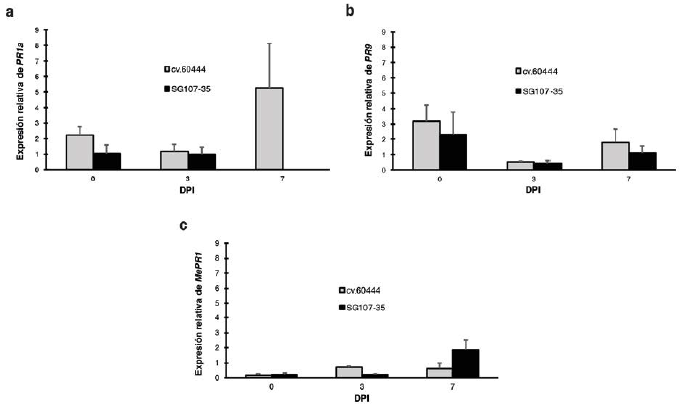
Con el fin de evaluar la expresión relativa de PR1a, PR9 y MePR1 en respuesta a la inoculación con Xam, se comparó la expresión de cada gen a lo largo del tiempo en ambas variedades con respecto a 0 dpi. Los resultados indican que la expresión de PR1a mostró un perfil interesante donde se observa claramente una inducción a los 3 dpi exclusivamente en la variedad resistente, incrementando su expresión a los 7 dpi (Fig. 4a).
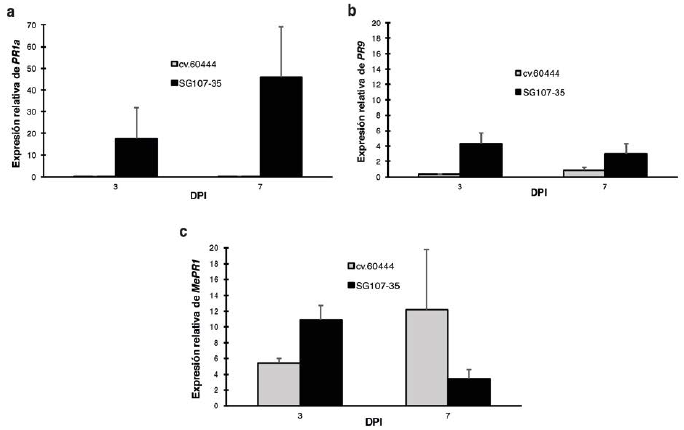
Correspondientemente, el perfil de expresión de PR9 es similar al de PR1a, sin embargo, los valores de expresión de PR9 en la variedad resistente son mucho más bajos comparados con la expresión de PR1a en esa misma variedad (Fig. 4b). Por otro lado, la expresión de MePR1 es inducida por la bacteria en la variedad resistente al compararla con la susceptible a los 3 dpi y no a los 7 dpi (Fig. 4c). De manera notable, estudios previos han mostrado que la herida que se genera al momento de la inoculación puede producir la inducción de genes de manera diferencial (Gomez et al. sometido). Por esta razón, se decidió evaluar la expresión relativa de PR1a, PR9 y MePR1 con respecto a la herida. Para esto, se realizó un nuevo cálculo de la expresión relativa de cada gen en las dos variedades inoculadas con Xam con respecto a la inoculación con MgCl2. Los resultados de este análisis muestran que la expresión relativa de PR1a y PR9 es muy similar en la variedad resistente comparada con la susceptible para los tres tiempos evaluados (Fig. 5a, 5b). En particular, el gen PR1a mostró una baja expresión en las dos variedades a los 0 y 3 dpi. Aunque a los 7 dpi hay una inducción en la variedad susceptible, por problemas técnicos no se pudo evaluar la expresión en este tiempo para la variedad resistente (Fig. 5a). Por otro lado, y de manera interesante se encontró una baja expresión del gen MePR1 a 0 y 3 dpi en ambas variedades y una clara inducción a 7 dpi exclusivamente en la variedad resistente (Fig. 5c). En conjunto estos resultados sugieren que, si bien parece presentarse una inducción de los genes PR1a, PR9 y MePR1 en la variedad resistente inoculada con Xam a lo largo del tiempo (Fig. 4), esta inducción puede estar explicada en parte por la herida que se realiza durante el proceso de inoculación.
DISCUSIÓN
Las proteínas PR tienen una actividad muy importante durante las respuestas inmunes (van Loon et al., 2006). La producción de tales proteínas es inducida por la activación de la señalización que se genera ante el reconocimiento del patógeno. Las proteínas PR tienen actividad antimicrobiana y afectan directamente la viabilidad y crecimiento del patógeno. Además, su inducción suele ser diferencial durante reacciones de resistencia y susceptibilidad (van Loon et al., 1999). La determinación de la expresión de las proteínas PR se convierte en un excelente marcador de la activación de la respuesta inmune (van Loon et al., 2006). A pesar de la importancia de este tipo de proteínas, son muy escasos los estudios de PRs en yuca durante la interacción con Xam (López et al., 2005; Yoodee et al., 2018). Para llenar este vacío en primera instancia se realizó una aproximación bioinformática basada en la similitud de estas proteínas entre especies para identificar un grupo de genes PR candidatos en el genoma de yuca. A partir de 50 secuencias de PR de otras plantas se logró obtener un grupo de 15 posibles genes PR en yuca. Las secuencias de los genes que codifican para proteínas PR hacen parte de familias multigénicas (Sudisha et al., 2012). Así por ejemplo para el caso de PR1 existe el PR1a, PR1b y PR1c. Algo similar sucede con el PR2, PR4, PR8 y PR10. Estas proteínas presentan un alto grado de especificidad en respuesta a un patógeno y son reguladas a nivel transcripcional (van Loon et al., 1999). Por esta razón resulta difícil determinar cuáles de los diferentes miembros identificados en yuca están implicados en la respuesta a Xam. Para cada uno de los genes PR se identificaron en general más de cuatro posibles genes en yuca. Esto llevaría a evaluar más de 50 posibles juegos de cebadores; por esta razón en este estudio se seleccionó sólo un subgrupo de ellos para evaluar experimentalmente el nivel de expresión en plantas de yuca de dos variedades, una resistente y otra susceptible durante la infección con Xam. Para definir respuestas diferenciales, clásicamente se ha empleado el valor de AUDPC (Area Under Disease Progress Curve) que está determinado por el desarrollo de síntomas en el tiempo. A mayores valores de AUDPC las plantas son más susceptibles. Las variedades SG107-35 y cv.60444 presentan una respuesta contrastante ante la cepa Xam686 con valores de AUDPC de 1,44 y 1,90 respectivamente, por lo cual se catalogan como resistente y susceptible respectivamente para esta cepa de Xam (Trujillo et al., 2014).
Los genes que codifican para proteínas PR por definición son aquellos que se inducen en respuesta a patógenos. En general, el análisis de expresión de los 14 genes PR evaluados mediante RT-PCR semicuantitativa demostraron en varios casos que la herida que se hace para llevar a cabo la inoculación genera por sí misma una respuesta de la planta evidenciada por una expresión tenue en plantas control absoluto o una expresión inducida y en algunos casos fuerte en las plantas tratadas con la solución de MgCl2, por lo tanto con esta información no es posible determinar si la inducción de los genes se da en respuesta a la herida o al patógeno (Fig. 3). Previamente se ha reportado la expresión de genes PR en plantas sanas, sin ningún tipo de estrés (Tuzun, 2001). En el caso de yuca quizás la herida en el tallo este generando una respuesta que puede ser similar a la que ocasionan patógenos necrótrofos o herbívoros, y esto puede llevar a la inducción de genes PR. Para lograr estimar la diferencia en la expresión de los genes entre la respuesta ocasionada por la herida y la infección ocasionada por Xam se realizó un estudio mediante QRT-PCR para algunos genes PR, el cual toma en consideración la expresión relativa de cada gen en las plantas inoculadas con Xam con respecto a las plantas tratadas con MgCl2. Mientras que el patrón de expresión del gen PR1 no mostró una inducción en respuesta a Xam, el gen MePR1 mostró ser claramente inducido a 7 dpi solamente en la variedad resistente (Fig. 5). Estos resultados apoyan la idea presentada anteriormente respecto a la presencia de varios miembros dentro de la misma familia de genes PR. Si bien uno de los miembros de PR1a (Manes.07G050700) no mostró un perfil correspondiente a la definición de gen PR, otro de ellos sí lo mostró (MePR1, Manes.06G026900.1). Estos resultados ponen en evidencia la necesidad de evaluar cada miembro de la familia frente a diferentes patógenos, pues uno de ellos puede ser considerado como un real PR frente a algunos patógenos y otros no. En arroz se ha reportado la inducción de dos PR1 (a y b) por la infección causada por el Magnaporthe grisea (Agrawal et al., 2001) y por Xanthomonas oryzae pv. oryzae (Mitsuhara et al., 2008). En Arabidopsis se demostró que de 22 genes PR1 predichos similares a PR1a de tabaco, sólo uno respondía a este patógeno (van Loon et al., 2006). Una situación similar se esta presentando en yuca, en donde no todos los miembros de la familia PR1 parecen estar implicados en la inmunidad.
Los genes de la familia PR1 son inducidos por ácido salicílico y por la presencia de patógenos, razón por la cual se ha empleado particularmente como marcador de resistencia. Sin embargo, Mitsuhara et al. (2008) reportan la inducción de genes de esta familia ante interacciones compatibles e incompatibles frente al añublo, aunque también se da frente a otros estímulos. A pesar de ser empleados extensamente en el estudio de interacciones planta patógeno, aún se desconoce la función de los genes PR1 . El gen MePR1 (Manes.06G026900.1) ha sido recientemente identificado como inducido en hojas de plantas de yuca in vitro durante la respuesta a Xam (Yoodee et al., 2018). El hecho de haber encontrado en este estudio una inducción diferencial de este mismo gen entre variedades resistentes y susceptibles, a diferentes tiempos y en el tallo de plantas de yuca provenientes de estacas corrobora su papel en la respuesta a Xam, su alta reproducibilidad y robustez erigiéndose como un excelente gen PR marcador de inmunidad.
CONCLUSIONES
Este estudio permitió demostrar la naturaleza inducida de algunos genes en respuesta a la infección por Xam. Bajos estas consideraciones se recomienda en particular el uso del gen MePR1(Manes.06G026900.1) como marcador de la resistencia, el cual podrá ser empelado en posteriores estudios. Por ejemplo, se podría esperar que en variedades silenciadas para genes de resistencia candidatos la expresión de este gen sea baja, tardía o nula debido a que no se activan las vías de respuesta. De manera similar, en plantas transgénicas con genes de resistencia heterólogos o complementadas con genes de resistencia endógenos se esperaría que presentasen una mayor inducción de este gen o que su expresión se vea inducida a tiempos mas tempranos.

