











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
El frijol común (Phaseolus vulgaris L.) es ampliamente consumido en todo el mundo. Se considera como la segunda fuente de proteína en África oriental y meridional, y la cuarta en América (Broughton et al., 2003; Lanna et al., 2016). Además, contiene abundante fibra dietética, carbohidratos multifacéticos, vitaminas y minerales (FAO, 2014). En Cuba, la obtención de este grano está a cargo fundamentalmente del sector agrícola no estatal, constituido en su mayoría por fincas y pequeñas parcelas, con condiciones muy diversas y baja disponibilidad de insumos agroquímicos y energéticos (ONEI, 2014). Es por ello, que el desarrollo de investigaciones encaminadas a la obtención de nuevas variedades adaptadas a estas amenazas edafo-climáticas, son esenciales para contribuir a la seguridad alimentaria de una población creciente que requiere incrementos significativos en la producción de alimentos. Por lo que dotar a los productores de nuevas variedades de frijol común mejoradas genéticamente a determinados caracteres de interés agronómico como son la tolerancia a sequía y salinidad, constituye una prioridad para alimentación humana a nivel mundial.
La obtención de cultivares resistentes o tolerantes a través de las técnicas de mejoramiento genético convencional en esta especie resulta difícil, debido al ligamiento genético y a las barreras de hibridación sexual (Veltcheva et al., 2005; McClean et al., 2008).
Como alternativa para complementar los métodos de cruzamiento tradicional, se ha empleado la inducción de mutaciones a través de mutágenos físicos y químicos. Dentro de los químicos se emplean agentes alquilantes, como el etilmetanosulfonato (EMS), bromuro de etidio, colchicina y análogos de bases como el bromouracilo (Mba, 2013). Por su parte, los físicos incluyen las radiaciones gamma, los rayos X, la luz ultravioleta y la radiación de partículas incluyendo neutrones rápidos y térmicos, así como partículas beta y alfa. Dentro de estos últimos la radiación gamma es la más utilizada, debido a su aplicación sencilla, penetración elevada en el tejido, reproducibilidad y alta frecuencia de mutación, de ahí que se han empleado con éxito en los programas de fitomejoramiento de muchas especies de plantas (Hasbullah et al., 2012).
En P. vulgaris los investigadores se han concentrado fundamentalmente en determinar el efecto de los mutágenos físicos sobre semillas y su evaluación en condiciones ex vitro (Ellyfa et al., 2007; Sood et al., 2014). Sin embargo, en la literatura científica se refieren un número reducido de trabajos donde se estudió el efecto de las radiaciones gamma in vitro sobre diferentes tipos de explantes en Phaseolus vulgaris L. (Bajaj et al., 1970; Carneiro et al., 1987). Esto se debe fundamentalmente a que la regeneración de plantas in vitro de P. vulgaris, resulta un proceso complejo debido a los efectos del genotipo en la capacidad regenerativa, así como la baja eficiencia y reproducibilidad de estos métodos (Sabzikar et al., 2010).
Collado et al., (2013) desarrollaron un protocolo eficiente de regeneración de plantas en cinco cultivares de P. vulgaris, dentro de estas se encuentra el cultivar "ICA Pijao". Dicho cultivar está incluido en los programas de mejoramiento genético que se desarrollan en el país, ya que es uno de los más atractivos para los pequeños productores, debido a que presenta un rendimiento potencial de 1.8 t. ha-1 rendimiento elevado. Además posee hábito de crecimiento erecto lo que permite la cosecha mecanizada. Sin embargo, presenta como limitante que es susceptible a la sequía, lo que reduce su extensión a otras regiones cultivables del Oriente y Centro del país (Singh, 1992).
En la literatura científica se plantea que el efecto de las radiaciones gamma es específico para las especies, el tiempo y las dosis (Udensi et al., 2012). De ahí que, además de disponer de un protocolo eficiente de regeneración de plantas in vitro, es necesario el estudio de diferentes factores que influyen en la inducción de variabilidad genética a través del empleo de las radiaciones gamma como son: el tipo de explante, la germinación de las semillas, la edad del explante, la inducción y multiplicación del callo, la regeneración de brotes.
Los estudios referidos en la literatura científica donde se combinan la mutagénesis con el cultivo de tejidos en P. vulgaris, determinan el efecto de las radiaciones gamma sobre diferentes tipos de explantes: callos, semillas, brotes (Bajaj et al., 1970; Carneiro et al., 1987). Sin embargo, estos trabajos no se han desarrollado en función del efecto de este tipo de explantes irradiados en la regeneración de brotes in vitro, sino solamente en la evaluación del crecimiento del callo a partir de la determinación de masa fresca, masa seca, coloración presente en los mismos, y en el caso de semillas y brotes, las evaluaciones solamente se han relizado en condiciones ex vitro. Además estos trabajos no definen un tipo de explante óptimo a emplear en los programas de mejoramiento genético de esta especie. Esto se debe fundamentalmente a que la regeneración de plantas in vitro de P. vulgaris, resulta un proceso complejo debido a los efectos del genotipo en la capacidad regenerativa, así como la baja eficiencia y reproducibilidad de estos métodos (Veltcheva et al., 2005). Es por ello que el objetivo del presente trabajo fue determinar el efecto del tipo de explante irradiado en la regeneración in vitro frijol común (Phaseolus vulgaris L.) cultivar "ICA Pijao" (Veltcheva et al., 2005).
MATERIALES Y MÉTODOS
Se empleó como material inicial semillas de dos meses después de cosechadas de Phaseolus vulgaris L. cultivar "ICA Pijao", desarrollado por el Instituto Colombiano Agropecuario (ICA) en 1974 y cultivadas en Cuba. El material de partida fue donado por el Centro de Investigaciones Agropecuarias de la Universidad Central "Marta Abreu" de Las Villas (CIAP). Las semillas colectadas fueron cosechadas en casa de cultivo.
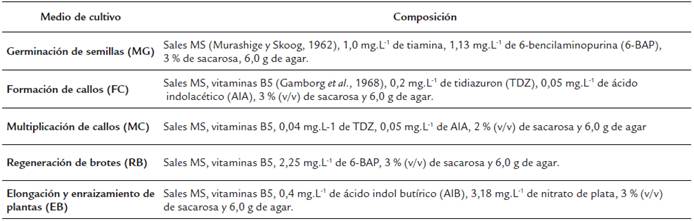
Para el desarrollo de los experimentos in vitro se empleó el esquema de trabajo desarrollado por (Collado et al., 2013), los medios de cultivo empleados en la investigación se describen en la Tabla 1. El pH de los diferentes medios de cultivo fue ajustado a 5,7 con hidróxido de sodio NaOH (0,1N) y cloruro de hidrógeno HCl (0,1N) antes de la esterilización en autoclave a una temperatura de 121 °C y 1,2 Kg.cm-2 de presión, durante 20 minutos. Para los experimentos se utilizaron magentas plásticas de 500 ml de capacidad total y se añadieron 60 ml de medio de cultivo por frasco de cultivo. La manipulación de los explantes se llevó a cabo en cabina de flujo laminar horizontal.
Tabla 1 Medios de cultivo utilizados en la regeneración de plantas vía organogénesis indirecta de P. vulgaris cultivar "ICA Pijao".
El material vegetal en todos los experimentos, se colocó en la cámara de crecimiento a una intensidad luminosa de 45 (μmol m-2 s-1 proveída por lámparas fluorescentes de 45 (μmol m2s-1 lámparas fluorescentes frías a una temperatura de 25 ±2 0C.
La irradiación del material vegetal se realizó con una fuente de 60Co en un equipo PX-Gamma 30 con una potencia de 33, 26 Gy.min-1 33, 26 Gy.min-1, el cual se encuentra situado en el centro de Estudios Aplicados al Desarrollo Nuclear de Ciudad de la Habana, Cuba (CEADEN).
Todos los experimentos fueron desarrollados bajo un diseño completamente al azar. Se aplicaron pruebas no parámetricas H de Kruskall-Wallis y la U-Mann-Whitney, previa comprobación de los supuestos de normalidad de los datos y homogeneidad de varianza. El procesamiento estadístico de los datos experimentales se realizó con la ayuda del paquete computacional Statistic Packaged for Social cience (SPSS) versión 21.0 sobre Windows.
Determinación del tipo de explante para la aplicación de radiaciones gamma en regeneración de plantas vía organogénesis indirecta de P. vulgaris cultivar "ICA Pijao"
Con el objetivo de determinar el efecto de diferentes explantes irradiados en la regeneración de plantas in vitro de P. vulgaris cultivar "ICA Pijao", se evaluaron tres tipos de explantes: callos, nudo cotiledonal con un cotiledón (NC-1), semillas maduras.
Callos
El presente experimento se realizó con el objetivo de determinar el efecto de callos irradiados en la regeneración in vitro de P. vulgaris cultivar "ICA Pijao". Para ello se emplearon callos obtenidos a partir de semillas de dos meses después de cosechadas en casa de cultivo. Se empleó el medio de cultivo de formación de callos (FC). Los callos se colocaron en una placa de Petri y se pesaron en una balanza digital (Daus modelo CS 200), y se colocó una masa fresca inicial de 0,3 g de callo por placa de Petri en medio de cultivo de multiplicación de callos (MC). Posteriormente, callos en un segundo subcultivo de multiplicación fueron tratados con las siguientes dosis de radiaciones gamma (60Co): 0, 10, 20, 30, 40, 50, 60Gy para un total de siete tratamientos. Se emplearon 100 explantes por tratamiento mutagénico y del control sin irradiar.
Se realizaron tres subcultivos en medio de cultivo de multiplicación de callos, para un total de 63 días. Cada 21 días se realizó la transferencia a medio de cultivo fresco y en el tercer subcultivo de multiplicación de callos posterior al tratamiento mutagénico se determinó la masa fresca (g) de los callos con el empleo de una balanza digital (Daus modelo CS 200). Se evaluó visualmente y se registró el número de callos según la coloración que estos presentaron. Posteriormente, a los 21 días en medio de cultivo de regeneración de brotes se registró el número de brotes por callo.
Nudo cotiledonal con un cotiledón (NC-1)
El presente experimento se realizó con el objetivo de determinar el efecto del nudo cotiledonal con un cotiledón (NC-1) irradiado sobre la regeneración in vitro de P. vulgaris cultivar "ICA Pijao". Como explante inicial se emplearon semillas maduras cosechadas en casa de cultivo. A los tres días, después de germinadas las semillas se eliminó la testa y se separaron los cotiledones. Se seleccionó el cotiledón unido al nudo cotiledonal (NC-1) como explante inicial para el tratamiento mutagénico. Los nudos cotiledonales con un cotiledón (NC-1) fueron tratados con las siguientes dosis de radiaciones gamma: 10Gy, 20Gy, 30Gy, 40Gy, 50Gy, 60Gy y un control sin irradiar.
Se emplearon 100 explantes (NC-1) por tratamiento mutagénico y del control sin irradiar a razón de cinco 5 (NC-1) por placa de Petri. La formación, multiplicación de callos y la regeneración de plantas se realizó según lo descrito por Collado et al., (2013).
A los 21 días en medio de cultivo de formación de callos posterior al tratamiento mutagénico se cuantificó el porcentaje de explantes que formaron callos. Se realizaron tres subcultivos en medio de cultivo de multiplicación de callos, para un total de 63 días. Cada 21 días se realizó la transferencia a medio de cultivo fresco y en el tercer subcultivo de multiplicación de callos se determinó la masa fresca (g) de los mismos con el empleo de una balanza digital (Daus modelo CS 200). Para determinar el color de los NC-1 y los callos se utilizó el código hexadecimal de colores. Posteriormente, a los 21 días en medio de cultivo de regeneración de brotes se registró el número de callos con brotes y el número de brotes por callo.
Semillas
El presente experimento se realizó con el objetivo de determinar el efecto de semillas irradiadas sobre la regeneración in vitro de P. vulgaris cultivar "ICA Pijao". Se emplearon semillas maduras cosechadas en casa de cultivo que fueron tratadas con las siguientes dosis de radiaciones gamma (60Co): 100Gy, 200Gy, 300Gy, 400Gy, y un control sin irradiar.
Se emplearon 100 semillas por tratamiento mutagénico y del control sin irradiar y se colocaron en cámaras de crecimiento en las condiciones referidas para la germinación de las semillas durante tres días, posteriormente 21 días en medio de cultivo de formación de callos (siete días en la oscuridad y 14 días a 16-8horas luz/oscuridad), se realizaron tres subcultivos en medio de cultivo de multiplicación de callos, para un total de 63 días a 16-8horas luz/oscuridad. Cada 21 días se realizó la transferencia a medio de cultivo fresco y 21 días en medio de cultivo de regeneración de brotes a 16-8horas luz/oscuridad.
Las evaluaciones se realizaron en la germinación de las semilla, formación y multiplicación de los callos y en la regeneración de plantas. En la germinación se cuantificó: el número de semillas que germinaron, a partir de estos valores se calculó el porcentaje de germinación por tratamiento y se midió la longitud de las raíces (cm) con una regla milimetrada. En la formación de los callos se registró el número de explantes que formaron callos, a partir de estos valores se calculó el porcentaje de explantes con callos y se cuantificó la masa fresca (g) de los mismos a los 21 días de cultivo y para ello se utilizó una balanza digital (Daus modelo CS 200).
Para determinar el color de los cotiledones y los callos se utilizó el código hexadecimal de colores. A los 21 días en medio de cultivo de multiplicación de callos se cuantificó la masa fresca (g) de los mismos. Posteriormente a los 21 días en medio de cultivo de regeneración de brotes se evaluó el número de brotes por callo.
RESULTADOS
Callos
La aplicación de las diferentes dosis de radiaciones gamma afectó la coloración y la consistencia de los callos irradiados. El 100 % de los callos no irradiados eran nodulares, compactos con una coloración verde (código hex: #008000) (Fig. 1a). Mientras que los callos irradiados, solo presentaron zonas verdes (código hex: #008000) y otros de color siena (código hex: #A0522D) (Fig. 1b-1g). Es de señalar que con las dosis de 50 y 60 Gy los callos perdieron la consistencia compacta y se disgregaron en el medio de cultivo.

Figura 1 Coloración presentada por los callos irradiados de P. vulgaris cultivar "ICA Pijao". a) Control, b) 10Gy, c) 20Gy, d) 30Gy e) 40Gy, f) 50Gy, g) 60Gy. Las flechas indican la coloración del callo cuya imagen aparece ampliada en la parte inferior de cada figura. (Barra=0,5cm).
La frecuencia de aparición de callos con zonas de coloración verde (código hex: #008000) varió desde el 70 % hasta el 30 % en las dosis más altas de radiación aplicadas. Con el empleo de 10 Gy, el 66 % de los callos mostraron zonas con coloración verde (código hex: #008000) y el 33,5 % de estos, mostraron una coloración siena (código hex: #A0522D). Por su parte, con la dosis de radiación de 20 Gy, disminuyó la frecuencia de aparición de callos con zonas verdes (código hex: #008000) hasta el 45 %. Con 30 y 40 Gy estos porcentajes estuvieron por debajo del 40 %. Las dosis de radiaciones más altas (50 y 60 Gy) mostraron más del 70 % de los callos con coloración siena (código hex: #A0522D).
Asociado a los cambios en la coloración de los callos se observó una disminución de la masa fresca y del porcentaje de regeneración de brotes de los mismos por frasco de cultivo en comparación con el control con diferencias significativas (Tabla. 2). Los mayores valores de masa fresca los mostró el control con diferencias significativas con las diferentes dosis de radiaciones, con un incremento de 0,24 g por encima de la masa fresca inicial de los callos. Con la dosis de 10 Gy se alcanzaron los mayores valores de masa fresca de los callos con diferencias significativas con las restantes dosis estudiadas. Sin embargo, respecto a la masa fresca inicial utilizada, se produjo una disminución significativa de la masa fresca de 0,25 g, si se tiene en cuenta que 10 Gy fue la dosis de radiación más baja de las estudiadas. Con las dosis de 30 y 40 Gy se alcanzaron valores de masa fresca (g) por frasco de cultivo similares entre ellos y superiores a los alcanzados con los tratamientos de 50 y 60 Gy, que presentaron los valores más bajos de la masa fresca de los callos por frasco de cultivo respecto a la masa fresca inicial (Tabla 2).
Tabla 2 Multiplicación de callos y regeneración de brotes a partir de callos irradiados de P. vulgaris cultivar "ICA Pijao" a los 21 días en el tercer subcultivo de multiplicación de callos y en la regeneración de brotes. Rangos medios con letras desiguales difieren según prueba H de Kruskal Wallis y la U de Mann Whitney p ≤ 0.05. Masa fresca inicial de los callos = 0,3 g.
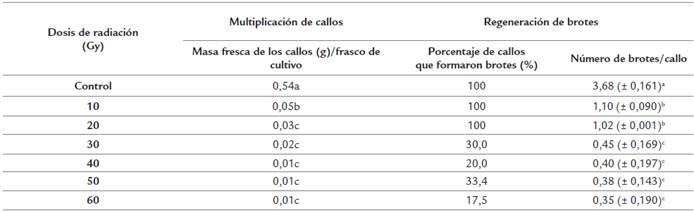
Las dosis de radiaciones gamma afectaron la regeneración de brotes a partir de callos irradiados con diferencias significativas en cuanto al número de brotes por callo entre las dosis de radiaciones estudiadas (Tabla 2). Con las dosis de radiación más bajas (10 y 20 Gy), se logró regenerar un brote por callo con diferencias significativas con las restantes dosis de radiación aplicadas. Con las restantes dosis se lograron valores similares en cuanto al número de brotes por callo y este valor fue inferior al de un brote por callo.
Los brotes formados a partir de los callos no irradiados, presentaron una coloración verde (código hex: #008000), con nervadura definida (Figura 2g). Mientras que los brotes formados con 10, 20 y 30 Gy, presentaron una coloración verde claro (código hex: #90EE90), hojas coriáceas, poca definición de las nervaduras de las hojas, ápices deformados (Figura 2a, b, c). Con las dosis de 40 y 50 Gy se formaron brotes en forma de roseta, con hojas hiperhídricas (Fig. 2d-2e) y con las dosis de radiación más alta (60 Gy), los callos solo presentaron zonas verdes (código hex: #008000), sin el desarrollo de brotes (Fig. 2f).

Fig 2 Morfología presentada por brotes regenerados a partir de callos irradiados de P. vulgaris cultivar "ICA Pijao". a) 10Gy (hojas coriáceas), b) 20Gy (hojas con ápice deformado), c) 30Gy (hojas con nervaduras poco definidas), d) 40Gy (hojas en forma de roseta, e) 50Gy (brotes con hojas hiperhídricas), f) 60Gy (callos con zonas de color verde (código hex: #008000), g) Control (brotes con hojas definidas). En las figuras (a, b, c, d y e), las flechas indican cambios en la morfología de las hojas de los brotes y en la figura f, la flecha indica secciones del callo con coloración verde. (Barra = 0,5cm).
Las radiaciones gamma afectaron considerablemente el callo, el cual fue empleado como explante inicial para la irradiación. Estas afectaciones estuvieron asociadas a la disminución de la masa fresca, disminución en la frecuencia de aparición de zonas de coloración verde (código hex: #008000), así como en el número de brotes por callo. Estos resultados indicaron que el uso de este tipo de explante para irradiar es limitado, ya que se compromete la multiplicación del mismo y la regeneración de brotes de P. vulgaris cultivar "ICA Pijao".
Nudo cotiledonal con un cotiledón (NC-1)
Todas las dosis de radiaciones gamma afectaron la formación del callo a partir del NC-1 irradiado, ya que se alcanzaron valores en el porcentaje de explantes con callo que variaron entre el 27 y el 2 % (Fig. 3).
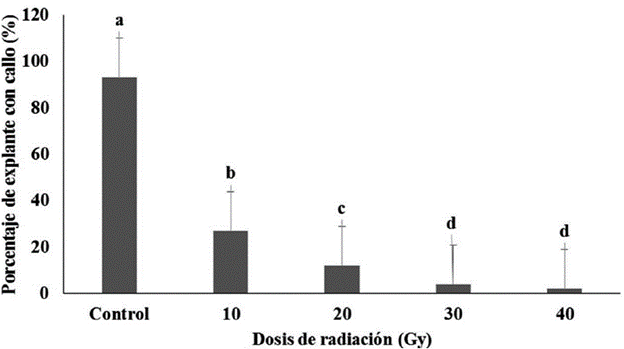
Figura 3 Porcentaje de nudo cotiledonal (NC-1) irradiado que formaron callo de P. vulgaris cultivar "ICA Pijao" a los 21 días en medio de cultivo de formación de callos. Rangos medios con letras desiguales difieren según prueba H de Kruskal Wallis y la U de Mann Whitney p ≤ 0.05
Los NC-1 irradiados con 10 Gy alcanzaron los mayores valores en el porcentaje de explante con callo, con diferencias significativas con las demás dosis de radiación utilizadas. Sin embargo con dicha dosis, los valores se redujeron en un 66 % respecto al control. Por su parte con la dosis de 20 Gy que difirió significativamente con las restantes dosis de radiaciones empleadas, el porcentaje de explante con callo se redujo respecto al control en un 81 %. Con 30 y 40 Gy, estos valores se incrementaron, y fueron de un 89 y 91 % respecto al control. Con las dosis de radiación superiores (50 y 60 Gy), no se logró la formación de callos.
A partir del NC-1 sin irradiar, los callos que se formaron fueron compactos, nodulares y de color verde (código hex: #008000).
Mientras que con las diferentes dosis de radiaciones los cotiledones cambiaron de color verde (código hex: #008000) a siena (código hex: #A0522D). Con las dosis de 10 y 20 Gy se logró la formación del callo nodular verde en el nudo cotiledonal, pero el cotiledón presentó una coloración siena (código hex: #A0522D). Con la dosis de 30 Gy se logró la formación del callo nodular, pero este fue de color siena (código hex: #A0522D) al igual que el cotiledón. Es de señalar que con la dosis de 40 Gy los explantes presentaron una coloración siena tanto en la posición abaxial como en la posición adaxial.
De igual forma, las dosis de radiaciones gamma utilizadas afectaron la multiplicación de callos y la regeneración de brotes formados a partir del NC-1 (Tabla 3). Los callos formados con 10 y 20 Gy presentaron valores similares en la masa fresca (g), pero estos difirieron significativamente con el control. Con estas dosis se redujo la masa fresca de los callos cuatro y seis veces respectivamente en relación con el control. Con las restantes dosis de radiación (30 y 40 Gy), no se logró la multiplicación de los callos, ya que estos mantuvieron el peso inicial y presentaron una coloración siena (código hex: #A0522D). Sin embargo, el control sin irradiar incrementó dos veces los valores de masa fresca (g) por frasco de cultivo.
Tabla 3 Multiplicación de callos y regeneración de brotes a partir de nudo cotiledonal (NC-1) irradiados de P. vulgaris cultivar "ICA Pijao" a los 21 días en el tercer subcultivo de multiplicación de callos y en la regeneración de brotes. Rangos medios con letras desiguales difieren según prueba H de Kruskal Wallis y la U de Mann Whitney p ≤ 0.05. Masa fresca inicial de los callos = 0,3g

De igual forma, las radiaciones gamma afectaron la regeneración de brotes en los callos formados en el NC-1. Solamente se logró la formación de brotes en los nudos cotiledonales con un cotiledón (NC-1) irradiados con 10 y 20 Gy, con valores inferiores respecto al control, ya que menos del ocho por ciento de los callos formaron brotes, a razón de un brote por callo (Tabla 3).
Los brotes formados a partir de NC-1 no irradiados se caracterizaron por presentar hojas definidas de color verde (código hex: #008000). Sin embargo, los brotes obtenidos a partir de NC-1 irradiados fueron hiperhídricos, con hojas estrechas, en forma de roseta.
Las radiaciones gamma afectaron considerablemente el NC-1, el cual fue empleado como explante para la irradiación. Estas afectaciones provocaron la disminución considerable del porcentaje de explante con callo, la masa fresca, cambios en la coloración de los cotiledones y los callos, lo cual incidió directamente en el porcentaje de callos que formaron brotes.
Estos resultados indicaron que el NC-1 es muy sensible a la aplicación de las radiaciones gamma, lo que limita su uso como explante inicial para inducir variabilidad genética en P. vulgaris cultivar "ICA Pijao".
Semillas
La aplicación de radiaciones gamma sobre semillas no afectó el porcentaje de germinación de estas, pero si la longitud de las raíces (cm) y la presencia de raíces secundarias. Se encontraron diferencias significativas entre los tratamientos y el control (Fig. 4). Con 100 Gy se lograron valores inferiores en la longitud de las raíces (cm) en comparación con el control, pero superiores a los valores registrados con las restantes dosis de radiación (Fig. 4). Aunque el control y la dosis de 100 Gy presentaron diferencias significativas en la longitud de las raíces, ambas mostraron desarrollo de raíces secundarias. Los valores de la longitud de las raíces (cm) en las semillas tratadas con 200 Gy fueron inferiores con diferencias significativas con el control y la dosis de 100 Gy pero significativamente superiores a las alcanzadas con 300 y 400 Gy. De igual modo fue la respuesta en esta variable con las dosis de 300 y 400 Gy, los cuales alcanzaron los menores valores en cuanto a la longitud de las raíces con diferencias significativas entre ellas y con las restantes dosis de radiación y el control. Con las dosis de 200, 300 y 400 Gy las semillas presentaron menor longitud de la raíz principal y no mostraron desarrollo de raíces secundarias.
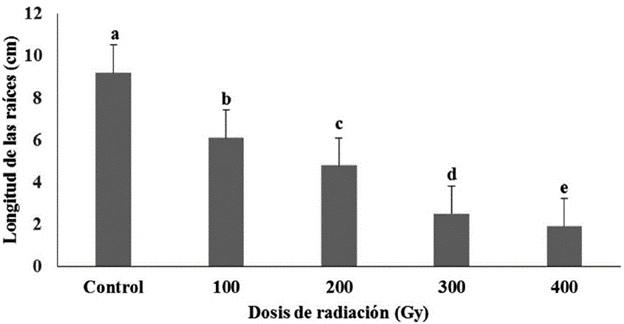
Figura 4 Longitud de las raíces en semillas irradiadas de P. vulgaris cultivar “ICA Pijao” a los tres días en medio de cultivo de germinación. Rangos medios con letras desiguales difieren según prueba H de Kruskal Wallis y la U de Mann Whitney p ≤ 0.05.
Al analizar el efecto de las radiaciones gamma en el porcentaje de formación de callos, se encontró que todas las dosis de radiación afectaron la formación de los callos con respecto al control a los 21 días en medio de cultivo FC (Tabla 4).
Tabla 4 Porcentaje de formación de callos a partir de semillas irradiadas de P. vulgaris cultivar "ICA Pijao" a los 21 días en medio de cultivo de formación de callos. Rangos medios con letras desiguales difieren según prueba H de Kruskal Wallis y la U de Mann Whitney p ≤0.05.

Con la dosis de 100 y 200 Gy se alcanzaron valores similares en la formación de callos a partir de semillas irradiadas y estos valores fueron inferiores al 25 %. Sin embargo con 300 Gy se alcanzaron valores inferiores en el porcentaje de formación de callos con diferencias significativas con 100, 200 Gy y el control. Con 400 Gy no se logró la formación de callos (Tabla 4).
Los callos del control fueron nodulares, verdes (código hex: #008000) y compactos, en comparación con los callos desarrollados a partir de semillas irradiadas con 100, 200 y 300 Gy, los cuales presentaron callos nodulares, compactos con zonas verdes (código hex: #008000) y callos nodulares de coloración siena (código hex: #A0522D) que no mantuvieron la consistencia compacta. Con estas tres dosis de radiación los cotiledones presentaron una coloración verde (código hex: #008000), mientras que con 400 Gy no se logró la formación de callos y el cotiledón presentó una coloración siena (código hex: #A0522D).
Las dosis de radiaciones estudiadas afectaron la multiplicación de callos y regeneración de brotes en las semillas irradiadas con diferencias significativas con el control (Tabla 5). A los 21 días en medio de cultivo MC, se observó que las dosis de radiaciones afectaron la masa fresca de los callos en la multiplicación. Con 100 Gy se lograron valores de masa fresca dos veces menores que las presentadas por el control. Sin embargo, con esta dosis los valores alcanzados fueron superiores a los logrados con las dosis de 200 y 300 Gy con diferencias significativas. Entre estas dos últimas dosis de radiación, los valores de masa fresca fueron similares y tres veces menores que el control (Tabla 5). Con las dosis de 100 y 200 Gy se lograron valores similares en el número de brotes por callo, pero estos fueron inferiores con diferencias significativas respecto al control sin irradiar. Los callos obtenidos a partir de semillas irradiadas con 300 Gy no formaron brotes (Tabla 5).
Tabla 5 Multiplicación de callos y regeneración de brotes a partir de semillas irradiadas de P. vulgaris cultivar "ICA Pijao" a los 21 días en el tercer subcultivo de multiplicación de callos y en la regeneración de brotes. Rangos medios con letras desiguales difieren según prueba H de Kruskal Wallis y la U de Mann Whitney p ≤ 0.05. Masa fresca inicial de los callos = 0,3g.
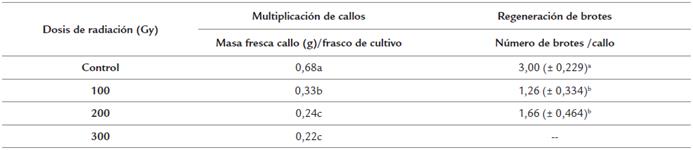
Los callos obtenidos a partir de semillas no irradiadas formaron brotes con hojas definidas de color verde (código hex: #008000) (Fig. 5a). Los brotes obtenidos apartir de semillas irradiadas también formaron brotes definidos, pero con hojas de color verde claro (código hex: #90EE90), hiperhídricas, y en algunos casos estas se tornaron de color amarillo (código hex: #FFFF00) (Fig 5-b, 5-c).

Figura 5 Brotes regenerados a partir de semillas irradiadas de P. vulgaris cultivar "ICA Pijao" con diferentes dosis de radiaciones gamma a los 21 días en medio de cultivo de regeneración de brotes: a) Control, b) 100Gy, c) 200Gy.
La aplicación de radiaciones gamma sobre semillas produjo afectaciones en la longitud y en la emisión de raíces secundarias, el porcentaje de explante con formación de callo fue inferior al 25 % con 100 y 200 Gy. La masa fresca de los callos, así como la regeneración de brotes se redujo dos veces en comparación con el control. Sin embargo, a partir de las semillas irradiadas con 100 Gy, fue el explante con el que se logró la regeneración de brotes con hojas definidas al igual que los brotes regenerados a partir de callos sin irradiar. Estos resultados indicaron que la semilla es el explante inicial para la aplicación de las radiaciones gamma para inducir variabilidad genética en P. vulgaris cultivar "ICA Pijao".
DISCUSIÓN
Es conocido que en el nivel de radiosensibilidad de diferentes tipos de explantes, pueden influir factores como el cultivar, las condiciones fisiológicas de las plantas u órganos a irradiar, el tipo de explante, así como la manipulación del material vegetal antes y después de la irradiación (Predieri, 2001). Por ejemplo, Bajaj et al., (1970); irradiaron callos de P. vulgaris L. y observaron, que con las dosis de radiación más altas (40 Gy), estos presentaron una coloración parda, lo que se correspondió con una disminución de los valores de masa fresca de los mismos. En otros cultivos también se refiere la presencia de cambios en la coloración de callos irradiados, por ejemplo, Yasmin et al., (2011), al irradiar callos de diferentes clones de caña de azúcar con las mismas dosis de radiación, definieron un tipo de callo nodular verde, compacto con capacidad para regenerar brotes, y un segundo tipo de callo, globular, no compacto.
Los resultados en la disminución de los valores de masa fresca de los callos aun con las dosis más bajas (10 y 20 Gy) para el cultivar estudiado y el tipo de explante utilizado, pudo deberse a un incremento en la susceptibilidad de las células a las mutaciones ocasionadas por la radiación acumulada en cada dosis (Predieri, 2001).
Los resultados en la disminución de los valores de masa fresca de los callos aun con las dosis más bajas (10 y 20 Gy) para el cultivar estudiado y el tipo de explante utilizado, pudiera estar relacionado con que el contenido de humedad y de oxígeno presente en los callos incrementa la susceptibilidad de las células a las mutaciones ocasionadas por la radiación acumulada en cada dosis (Predieri, 2001).
Al respecto, Gómez et al., (2001) irradiaron callos de crisantemo (Dendrathema grandiflorum Ramat) con dosis de 5 a 20 Gy, y evaluaron su crecimiento a los 7, 14, 21 y 28 días posteriores a la irradiación. Ellos encontraron que los mayores valores de masa fresca se obtuvieron con la dosis de radiación más baja de las estudiadas, mientras que con la dosis de 20 Gy estos valores disminuyeron en todas las fechas evaluadas. Los resultados obtenidos en la presente investigación están en correspondencia con los resultados obtenidos por dichos autores, ya que con la dosis de 10 Gy que fue la más baja de las estudiadas, se obtuvieron los menores valores en cuanto a la reducción de la masa fresca en comparación con las restantes dosis de radiación aplicadas, aunque no incrementaron su masa fresca como lo registrado para el control. Estas reducciones pudieran estar relacionadas con que el callo es un tejido conformado por células con alta capacidad de diferenciación (morfogénesis) y muy susceptibles a los cambios, principalmente cuando se altera el Ácido desoxirribonucleico ADN (Freire, 2003).
Ha sido descrito en la literatura científica, que la inducción de radiaciones gamma puede influir en la morfología de brotes regenerados en condiciones in vitro, ya que dosis bajas inhiben la síntesis de auxinas, mientras que dosis más altas pueden destruir directamente la actividad de la auxina. Esta se encuentra en la planta en mayores cantidades, en las partes donde se presentan procesos activos de división celular, lo cual se relaciona con sus funciones fisiológicas asociadas con la elongación de tallos, formación de raíces adventicias (McSteen y Zhao, 2008), por lo que la inhibición de la ruta biosintética podría haber influido directamente sobre la disminución del desarrollo de brotes en los callos irradiados en el presente trabajo.
No se han encontrado experiencias de trabajos similares, donde se estudie el nudo cotiledonal con un cotiledón como explante inicial para la aplicación de radiaciones gamma en P. vulgaris en la literatura científica consultada, sin embargo, varios autores refieren la importancia de la presencia del nudo cotiledonal para la formación de callos en los protocolos de regeneración de plantas vía organogénesis indirecta en esta especie (Arellano et al., 2009; Collado etal., 2013) y en la presente investigación con el empleo de las radiaciones gamma, los cotiledones mostraron en la mayoría de las dosis, una coloración parda, lo que pudo incidir en la formación de callos.
Sobre esto, Coelho y Benedito (2008), señalaron que durante el desarrollo de la semilla del frijol común; se sintetizan auxinas, citoquininas y giberelinas que se almacenan en los cotiledones. Estos reguladores del crecimiento, promueven el desarrollo del callo y son una posible causa de la respuesta diferencial observada en la formación de callos en los explantes irradiados en el presente trabajo, de ahí que en los nudos cotiledonales irradiados donde se produjo la caída del cotiledón, la formación de callos se redujo.
Los cotiledones pueden ser una fuente de reguladores de crecimiento (Kamisaka y Shibata, 1977) y en muchas especies, los explantes de los cotiledones o hipocotilos de plantas tienen un alto potencial morfogenético. Estos mismos autores plantearon que esto se debe a que estos tejidos son esencialmente juveniles, o porque los cotiledones son fuentes de sustancias difusibles promotoras de crecimiento.
En relación a esto, Burger y Hackett (1986), observaron en Cítrus sinencis L. "Valencia" que la presencia o ausencia de cotiledones afectó la capacidad para formar brotes. Ellos encontraron que los explantes que mantuvieron el nudo cotiledonal formaron más brotes que aquellos donde se produjo la pérdida del mismo. También sugirieron que estos resultados fueron consistentes con la hipótesis de que un promotor difusible (s) emana de los cotiledones.
Los resultados obtenidos en la presente investigación en semillas como explante inicial difieren con los trabajos donde se ha combinado la mutagénesis con el cultivo de tejidos en P. vulgaris (Bajaj et al., 1970; Carneiro et al., 1987). Estos autores utilizaron dosis de radiación inferiores, a las utilizadas en la presente investigación, así como un cultivar diferente de P. vulgaris ("Milionario 1732"). Las condiciones de cultivo empleadas por dichos autores fueron: semillas con dosis entre 5-400 Gy (Bajaj et al., 1970) y 40-200 Gy (Carneiro et al., 1987). Como resultado, en ambos trabajos se refiere la reducción de los porcentajes de germinación frente a las dosis de radiación más altas. Las diferencias encontradas con ambos trabajos, pudiera deberse a las dosis de radiación aplicadas, el nivel de susceptibilidad de los explantes ante las mismas, así como la respuesta diferencial de las variedades utilizadas en ambos estudios.
Las afectaciones en el porcentaje de germinación de semillas irradiadas de P. vulgaris pero en condiciones ex vitro, también ha sido informado por diferentes autores, por ejemplo, Ellyfa et al., (2007) indicaron que al irradiar semillas de P. vulgaris, con dosis de 300 y 400 Gy, no encontraron diferencias en cuanto al porcentaje de germinación en las semillas irradiadas con respecto al control. Los resultados del presente trabajo coinciden con lo informado por dichos autores, ya que con 400 Gy no se encontraron diferencias significativas con respecto al control en cuanto al porcentaje de germinación, no siendo así para la longitud de las raíces de las semillas germinadas.
En otros cultivos como: lenteja (Lens culinaris Medik) (Kumar y Sinha, 2003), Phaseolus lunatus L. (Kumar et al., 2003), arroz (Oryza sativa L.) (Pons et al., 2001) y trigo (Triticum aestivum L.) (Borzouei et al., 2010), se ha informado, que incrementos en las dosis de radiación gamma, causan reducción en los porcentajes de germinación. Estos autores atribuyeron estos resultados principalmente a la inhibición de procesos biológicos y fisiológicos necesarios para que tenga lugar la germinación de las semillas, como son la actividad enzimática, el balance hormonal, así como la inhibición del proceso de mitosis.
El presente estudio confirmó que las dosis de radiación gamma aplicadas no afectaron el porcentaje de germinación, pero si provocaron una reducción significativa del crecimiento de las raíces. Un resultado similar en la germinación ha sido descrito por Chaudhuri (2002), para el caso de la irradiación de semillas de lenteja (Lens culinaris Medik), donde la longitud de las raíces en las semillas germinadas se redujo a medida que se incrementaron las dosis de radiación. De igual forma, Borzouei et al., (2010) indicaron que al irradiar semillas de Triticum aestivum L. con las mismas dosis de radiación empleadas en el presente estudio (100, 200, 300 y 400 Gy), la longitud de las raíces en las semillas irradiadas disminuyó en la medida en que se incrementaron las dosis de radiación con respecto al control sin irradiar.
La reducción en el porcentaje de formación de callos a partir de semillas irradiadas a partir de los 21 días de cultivo pudo estar dado, por el aumento del efecto de la radiación en la formación de tejido nuevo. Con relación a esto, también se ha descrito en la literatura el efecto del agotamiento del medio de cultivo, lo que trae consigo que en la estructura morfológica del callo se pueden formar con el tiempo de cultivo, masas de células en la periferia externa del mismo, que obstaculizan la absorción de los nutrientes (Pérez, 1998), lo cual pudo haber influido en la disminución del porcentaje de formación de callos a partir de semillas irradiadas obtenidos en la presente investigación.
En relación con esto, Khan et al., (2002) observaron que la aplicación de radiaciones gamma a las semillas disminuye la capacidad de desarrollo de los callos obtenidos a partir de estas. Por otra parte, con 400 Gy no se logró la formación de callos. Ello sugiere que dosis altas de radiación gamma provocan la inhibición del crecimiento de los callos, lo que se pudiera atribuir a la detención del ciclo celular en la fase G2 / M durante la división de las células somáticas y/o diversos daños en la totalidad del genoma (Preuss y Britt, 2003).
Valdéz et al. (2004), por su parte evaluaron el efecto de diferentes dosis de radiaciones gamma (10, 20, 30, 40, 50, 60, 70, 80 Gy), inferiores a las empleadas en el presente trabajo, en callos de caña de azúcar (Saccharum spp.) y obtuvieron como resultado que la masa fresca de los callos, se redujo a medida que aumentó la dosis de radiación hasta la dosis de 30 Gy.
En la presente investigación, los callos obtenidos de semillas irradiadas con 300 Gy solo lograron un incremento en la masa fresca de 0,3 g, lo cual equivale a callos con un crecimiento muy limitado. Los resultados indicaron que para este cultivar de frijol común, dosis superiores a 200 Gy de radiaciones gamma en la semilla, comprometen el crecimiento y desarrollo del callo.
La disminución en cuanto al número de brotes regenerados a partir de semillas irradiadas también se ha informado, pero en trabajos referentes al mejoramiento genético de especies ornamentales como por ejemplo, lo informado por Shukla y Data (1993), quienes encontraron que al irradiar semillas de Chrysanthemum J., con 25 Gy, disminuyó el número de brotes regenerados por explante. Estos resultados coinciden con lo encontrado en el presente trabajo para P. vulgaris cultivar "ICA Pijao", donde se observó también una disminución en el número de brotes regenerados por callo. De igual forma, Bodele (2013), refieren que al irradiar semillas de Andrographispaniculata (Burm.f.) Wall. Ex. Nees con 10Gy, disminuyó el número de brotes regenerados y la mayoría de estos presentaron una reducción en la longitud del tallo y la raíz.
Los resultados del presente ensayo evidenciaron que la semilla irradiada fue el explante que favoreció la formación y multiplicación de callos, así como la formación de brotes con hojas verdaderas en frijol común cultivar "ICA Pijao".
CONCLUSIONES
Los diferentes explantes irradidos afectaron la regeneración in vitro de P. vulgaris cultivar "ICA Pijao" vía organogénesis indirecta. Estas afectaciones provocaron la disminución considerable del porcentaje de explante con callo, la masa fresca, cambios en la coloración de los cotiledones y los callos, y el porcentaje de regeneración de brotes en los tres tipos de explantes evaluados, sin embargo los resultados indicaron que los callos y el NC-1 resultaron los explantes más sensibles a la aplicación de las radiaciones gamma, lo que limita su uso como explante blanco para inducir variabilidad genética en esta cultivar. A partir de las semillas irradiadas con 100 Gy, este fue el explante con el que se logró la regeneración de brotes con hojas definidas al igual que los brotes regenerados a partir de callos sin irradiar, por lo que se recomienda como explante inicial para el uso combinado de la mutagénesis y la regeneración in vitro de plantas de P. vulgaris cultivar "ICA Pijao" vía organogénesis indirecta en la búsqueda de tolerancia a estrés abiótico en los programas de mejoramiento genético de esta leguminosa de gran importancia para la alimentación humana a nivel mundial.


