











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkINTRODUCTION
Human health, economy, and marine ecosystems have been affected in the last decades by toxic events and harmful algal blooms (HABs) caused by microalgae. In Colombia, these events have generated consequences ranging from the death of fish and birds to human poisonings (Mancera-Pineda et al., 2009). These events are not yet recognized as a serious problem, and there are no regulations, monitoring programs, or extensive studies about these organisms (Lozano et al., 2011; Mancera-Pineda et al., 2014).
Studies carried out on San Andrés Island have found dinoflagellates in seagrass beds belonging to the genera Prorocentrum and Ostreopsis, which have been associated with the production of toxins causing diarrhea and ciguatera on the island (Rodríguez et al., 2010). These and other toxins generated by microalgae have become a serious problem, because they can accumulate as trophic levels increase, creating a hazard for humans and other organisms.
The incidence of ciguatera in the South Pacific has increased by 60 % over the last 30 years (Skinner et al., 2011) and could increase further due to changes in global weather patterns, degradation of the coastal marine environment and overexploitation of marine resources (Tester et al., 2010; Gingold et al., 2014; Celis and Mancera-Pineda, 2015). A study carried out in the Caribbean indicates that between 1980 and 2010 there have been 10710 registered cases of ciguatera, with an average annual incidence of 42/100 000 inhabitants. This study also shows an increase in the incidence of ciguatera in the Caribbean during the last 31 years, mainly in the Eastern Caribbean (Celis and Mancera-Pineda, 2015).
The increase in ciguatera is worrisome considering that more than 116 million people live on the Caribbean coast and that more than 25 million tourists visit this region every year. Tourism income amounts to $ 25 billion per year for the region (Burke and Maidens, 2005; Celis and Mancera-Pineda, 2015).
The city of Cartagena de Indias declared as "Heritage of Humanity" by UNESCO in 1984, along with its neighbor Islas del Rosario National Natural Park represent the most important tourist and historic district of Colombia. Recent blooms of dinoflagellates have made evident the need to increase the local understanding of the dynamics of algal blooms in the region to improve the ability to cope with its impacts, which could be highly negative for the population and industry (Salon-Barros et al., 2016). The objectives of this research were to determine the composition, abundance, and seasonality of potentially toxic dinoflagellates associated to seagrass on Isla de Barú, aimed to the understanding of distribution patterns and ecological dynamics of toxic microalgae.
MATERIALS AND METHODS
Study area
The Barú Island or "Isla de Barú" is located 18 km from the city of Cartagena in the Department of Bolívar (Colombia) (10° 18' 03" - 10° 08' 02", N and 75° 42' 28"-75° 31' 07", W.) and is approximately 35 km long ranges from 1.5 km to 10 km wide. It has a warm climate with temperatures ranging from 27 °C to 30 °C, which presents a bimonthly unimodal regime, with a dry period that covers the months of December to April and a rainy period from May to November (Berdugo et al., 2004). However, the El Niño event of 2015 decreased the extension and frequency of historical rainfall patterns of Isla de Barú.
Isla de Barú belongs to the Islas del Rosario National Park, where activities such as fishing, conventional tourism, cultivation of small plots and the extraction of mangrove resources among others are carried out.
Sampling sites
The samples were collected during April 11 and September 25 of 2015 at three sites on the Isla de Barú: Isla Arena (IA) (10° 8' 40' N, 75° 43' 37'' W), Playa del Muerto (PM) (10° 8' 22" N, 75° 40' 42" W) and Ciénaga Cholón (cC) (10° 9' 53" N, 75° 40' 10" W) (Fig. 1). The type of sediment is highly variable between Isla Arena and Playa del Muerto, the former mainly consists of medium and coarse sands, while the second is made up of fine sands and silts (Table 1).
Table 1 Characteristics of the sampling sites in 2014 and 2015.
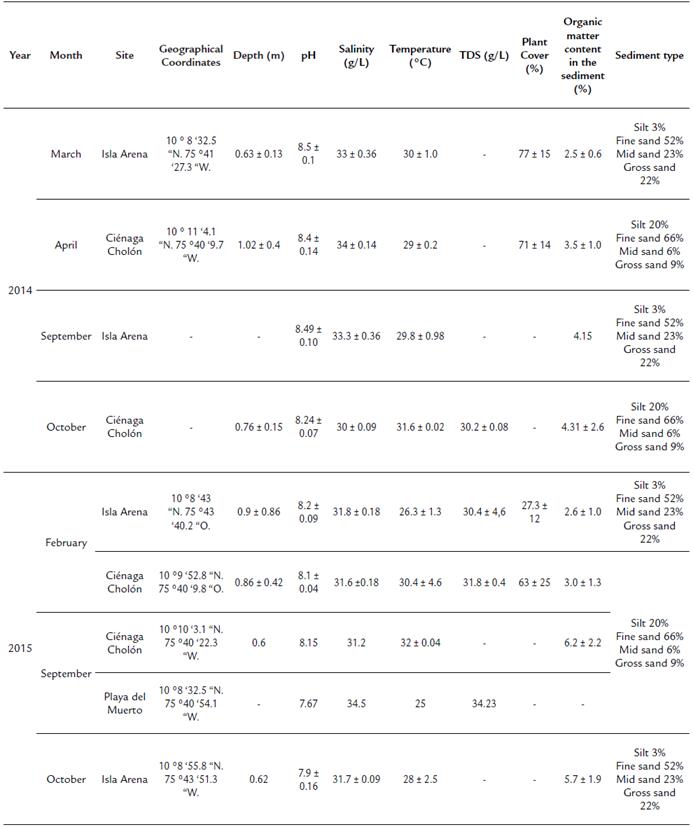
(-) No Data. TDS: Total dissolved solids.
Isla Arena is located offshore 5.2 km to the west of the coast of the peninsula. It is a small private island located in the open ocean, so human intrusion into this place is limited compared to other islands of the archipelago.
Playa del Muerto is located on the eastern side at 0.8 km from the town center of Barú bordering Barbacoas Bay. It is a place with a variety of seagrasses; it brings the waters of the Magdalena River that flow into the Barbacoas Bay through the Caño Lequerica.
Ciénaga Cholón is located 3.1 km north of the town of Barú. It is a tourist place that is affected by garbage problems caused by a large number of people who visit it and by the trade generated in the area. Similarly, both seagrasses and macroalgae are also affected by the motorboat rotors.
Sampling
Representative samples of seagrasses were collected by snorkeling. The sites selected for sampling had seagrass beds consisting mainly of Thalassia testudinum. Three quadrants of 25 cm x 25 cm were marked between 10 to 15 m from the coast at a water depth of approximately 1.5 m. All material within each quadrant was collected using labeled plastic bags (date, collection site, station). The sample bags contained previously filtered seawater according to the methodology described by Rodríguez et al., (2010).
At each sampling site, seawater samples (500 mL) were collected at a depth of 1 m. The water was filtered the same day of the collection, using Sartorius™ glass fiber filters with pore size of 0.7 pm. Ammonium (NH3-N), nitrates (NO 3 ) and phosphorus (PO 4 3- ) concentrations of each sample were measured using the HACH DR/200 spectrophotometer and polyvinyl alcohol reagents, mineral stabilizer, Nessler reagent (for ammonium), NitriVer 3 and NtriVer 6 (for nitrates), and PhosVer 3 (for phosphorus), according to the manufacturer's protocols and recommendations (HACH ®).
Sample processing
Each substratum (seagrass or macroalgae) was sorted by species and stored in plastic bags with a known volume of a fixative/preservative solution composed by 50 % filtered seawater and 50 % Transeau (six parts of water, three parts of alcohol, one part of formalin). Subsequently, agitation of each sample was carried out for five continuous minutes to guarantee a greater detachment of the epiphytic dinoflagellates. The contents of each subsample were filtered through a filter of 6 pm pore size; then the filtrate was re-suspended in a 15 mL volume of fixative solution. The microalgae-free substrata were weighed and dried at a temperature of 60 °C to determine the number of cells / g dry weight.
Identification and quantification
The dinoflagellates obtained from each sample were identified by light microscopy SCIENTIFIC MODEL 85-230V50 using the objective of (40X) and scanning electron microscopy (when possible). Electron microscopy was carried out using a FEI QUANTA 200-r equipment; the samples were dehydrated using a series of ascending concentrations of absolute ethanol. Ethanol concentrations of 30 %, 50 %, 70 %, 90 %, 95 % (10 minutes each) and 100 % (15 minutes) were used. Subsequently, a metallic gold coating was carried out using a Quorum Q150R ES sputter coater before observation.
The preliminary characterization of dinoflagellates was carried out based on dichotomous keys (Balech, 1988; Faust, 1990; Hernández and Almazán, 2004; Parra et al., 2011; Vidal and Lozano, 2011; Hoppenrath et al., 2014).
Epiphytic dinoflagellates were quantified by triplicate using a Sedgewick-Rafter chamber. The densities of epiphytic dinoflagellates are expressed regarding cells/g substrate wet weight, establishing the number of individuals present in the volume of the sample and dividing by the weight of the substrate.
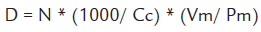
Where:
D = Cellular density in cells/g wet weight or cells/g dry weight of substrate
N = Number of counted cells of a given species
Cc= Number of frames scanned in the chamber
Vm = Sample volume (ml)
Pm: Wet or dry weight of the substrate (g)
Statistical analyses
Given the nature of the biological data collected on Isla de Barú, determined by a Shapiro-Wilk test, we proceeded as follows. The differences in the number of microalgae present at different sampling sites of Isla de Barú (Playa del Muerto, Isla Arena and Ciénaga Cholón) were determined by a one-way ANOVA test. The difference in the number of microalgae between months/seasons (April and September) for each of the sampling sites was determined by a T-test for related samples. All analyses were carried out using SPSS Statistics®.
ONI (Oceanic Niño Index)
The ONI calculated by the National Oceanic and Atmospheric Administration (NOAA) is based on sea surface temperature anomalies that exceed a threshold of +/- 0.5 °C in the three-month moving average in the Niño 3.4 region (5°N - 5°S, 12°O - 170°W) (Coronado, 2015; NOAA, 2015).
RESULTS
Three species of seagrasses, Thalassia testudinum, Syringodium filiforme and Halodule wrightii, were found on Isla de Barú, and T. testudinum was the most abundant and dominant. Although the cell densities of epiphytic dinoflagellates found on Isla de Barú in April and September of 2015 did not show statistically significant differences between sites or months of sampling (p>0.05), some trends were detected. The mean number of cells/g substrate dry weight (± standard deviation) was lower in April (142 ± 109.67, 164 ± 27.30 and 132 ± 32) compared to September (342 ± 231.3, 127 ± 29.8, 1032 ± 1023.5) for Ciénaga Cholón, Isla Arena and Playa del Muerto, respectively (Table 2).
Table 2 Averages and standard deviation of the density of dinoflagellate species in different months, sampling sites and substrates, on Isla de Barú during 2015.
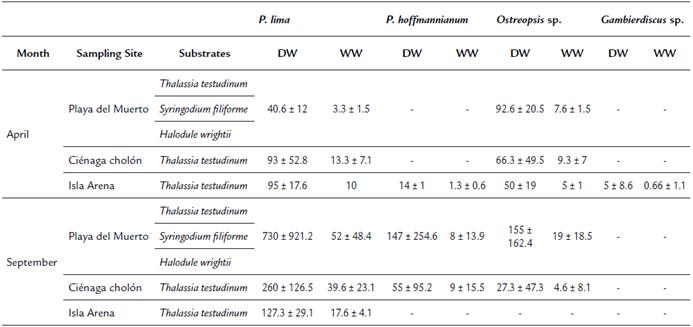
(DW) cells / g dry weight, (WW) cells / g wet weight and (-) absence.
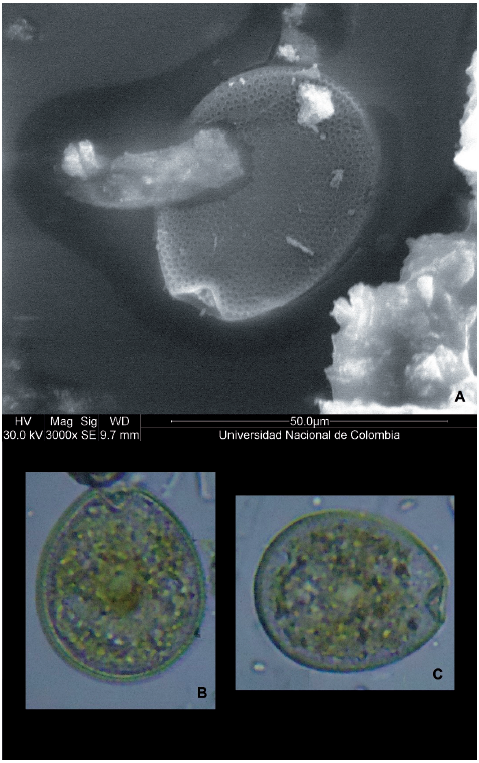
Four epiphyte dinoflagellate species were found: Prorocentrum lima, Prorocentrum hoffmannianum (Fig. 2), Ostreopsis sp., and Gambierdiscus sp.
The number of epiphytic diatoms found in Ciénaga Cholón were statistically significantly higher in September than in April (p<0.05) (Fig. 3). Diatom cell densities were higher on Isla Arena in April and Ciénaga Cholón in September compared with dinoflagellates.
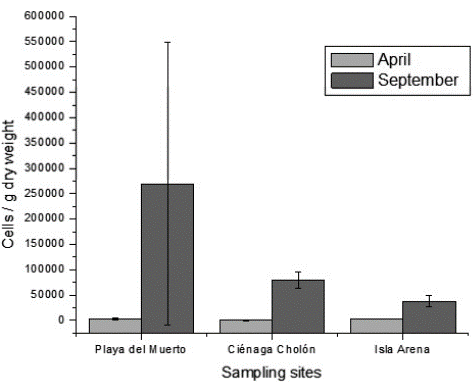
Figure 3 Total cell densities of epiphytic diatoms in Playa del Muerto, Ciénaga Cholón and Isla Arena (cells / g dry weight) in both months. The error bars represent the 95% confidence interval.
Eleven diatom species were found. Entomoneis sp., Cocconeis sp., Actinocyclus normanii, Nitzschia longissima, Diploneis crabro, Licmophora ehrenbergii, L. remulus, Plagiotropis lepidoptera, Mastogloia corsicana, Rhabdonema adriaticum, and Thalassionema nitzschioides.
This is one of the few records of Mastogloia corsicana in the Colombian Caribbean (Fig. 4), it is important to highlight that this species has been reported for the Caribbean by Navarro (1983); Navarro and Hernández-Becerril (1997); as a new record for Puerto Rico and the Caribbean (Navarro et al., 1989); and, in the East Caribbean by Loir and Novarino (2014). To date, none of these diatom species have been reported as toxic.

Prorocentrum lima (52±48 cells/g substrate wet weight) was the epiphyte dinoflagellate with greater presence, followed by Ostreopsis sp. (19±18 cells/g) and Prorocentrum hoffmannianum (27±24 cells/g).
The N:P ratios were considerably low for all sampling sites, ranging between 0.08 to 1.71, indicating that nitrogen was limiting biological production. Likewise, phosphorus was more abundant, followed by ammonium. According to the typical levels of nutrients in seagrass habitats described by Burkholder et al. (2007), the nitrate values at the three sampling sites are within the expected range of 2 to 10 μM/L (PM: 0.48±0 μM/L, IA: 0.16±0.16 pM/L, CC: 2.58±0.96 μM/L). Ammonium concentrations are below the expected range of 1 to 180 μM/L (PM: 26.05±16.07 μM/L, IA: 221.17±19.95 μM/L, CC: 27.76±4.98 μM/L) and phosphorus within the range between 0.3 to 20 μM/L (PM: 9.79±8.32 pM/L, IA: 46.15±4.32 μM/L, CC: 10.64±11.59 μM/L). Sampling site values were obtained in the present study.
The ONI data show that in 2014 there was a weak El Niño, which increased to very intense ONI category in the second half of 2015 up to the first quarter of 2016. This was the second strongest El Niño in the last 65 years. El Niño usually causes a decrease in precipitation on Isla de Barú; the 2015 event decreased rainfall 31 % compared to the historical average.
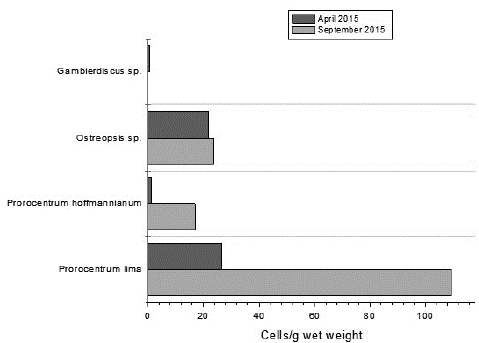
The epiphytic dinoflagellate species found on Isla de Barú showed a tendency to increase with the ONI category increase between April (ONI: 0.7 El Niño) and September (ONI: 1.8 El Niño) 2015. The largest increase (cell abundance) occurred in species belonging to the genus Prorocentrum (Fig. 5).
DISCUSSION
Three genera of epiphytes dinoflagellates occurred on Isla de Barú, Gambierdiscus, Ostreopsis and Prorocentrum. These genera contain toxin-producing species, some of which are related to toxic events as ciguatera.
Prorocentrum lima was the only dinoflagellate present at all sampling sites during both seasons followed by Ostreopsis sp. that only was absent on Isla Arena in September. P. lima was also found in each examined substrate (T. testudinum, S. filiforme and H. wrightii).
Other studies have found much higher quantities than those found on Isla de Barú. For example, a study conducted in Cuba by Delgado (2005) on different yielded of 37778 ± 2399 cells/g substrate wet weight, for P. lima, 6716 ± 1124 cells/g for Gambierdiscus sp. (reported as G. toxicus), and 471 ± 42 cells/g for O. lenticularis. The author also mentions that P. lima was always found in higher densities than Gambierdiscus sp., without observing any preference for the substrate. Ballantine et al. (1985) recorded highly variable abundances of Ostreopsis species with peaks up to 43000 cells/g substrate wet weight in the coastal waters of southwestern Puerto Rico. Other studies conducted at the coast of the Yucatan Peninsula have shown values of 1600 (approx.) and 9988 for P. lima and Gambierdiscus sp., respectively (Okolodkov et al., 2014). However, some authors argue that toxic events generated by Prorocentrum and Gambierdiscus species can be generated even if the concentrations of the organism that cause them are very low (Masó and Garcés, 2006).
Mancera-Pineda and colleagues (2014) have reported values of 6622 ± 8864 cells/g substrate dry weight for P. lima, 3608 ± 2789 cells / g dry weight for P. hoffmannianum and 1190 ± 1650 cells/g substrate dry weight for Gambierdiscus sp. (reported as G. toxicus), in floating organic material on San Andrés Isla. In addition, abundances of 59 ± 39 cells/g substrate dry weight for P. lima, 157 ± 110 cells/g substrate dry weight for P. hoffmannianum and 110 ± 177 cells/g substrate dry weight for Ostreopsis sp. on T. testudinum have been found on San Andrés Isla (Rodriguez et al., 2010).
Morton and Faust (1997) have mentioned that the low quantities of toxic dinoflagellates have been associated with the dominant seagrass T. testudinum. These authors mention that the low incidence of ciguatera in the barrier reef ecosystem of southern Belize could be triggered by this seagrass. This low incidence of ciguatera is because Thalassia testudinum is produced tannins under wound conditions (Arnold et al., 2008), and these tannins act as algaecides against dinoflagellates (Nagayama et al., 2003).
Working in Chengue Bay (Colombian Caribbean) during 2014, Arbeláez-Merizalde and Mancera-Pineda (2016) found that P. lima was the most abundant and frequent epiphytic dinoflagellates species. Cell densities ranged from 232 to 15867 cells/g substrate wet weight, with an average of 3081 and maximum values during the rainy season (August and September). They report that a slight decrease in salinity (from 37 to 36), together with an increase of the temperature (>29.60 °C) and low concentration of N (1.3 to 2.4 μg N/L), favor the growth of P. lima.
EL NIÑO EVENT
Dinoflagellate dynamics in the Caribbean is poorly understood. Therefore, the identification of climatic and oceanographic signals might serve as early warning of proliferations and useful to development predictive simulations. El Niño and La Niña events affect a variety of factors. During El Niño, environmental changes are generated such as sea level rise, changes in circulation patterns, weaker winds, decrease in primary production (phytoplankton), subsidence of the thermocline, decrease in water enrichment (Gajardo et al., 2013; Calvo et al., 2014). La Niña, conversely, causes an increase in precipitation, generates the outcropping of subsurface waters with lower temperatures than normal in the central and eastern equatorial Pacific Ocean. This outcropping causes nutrient-rich, cold Humboldt Current flows along the South American coast increasing primary (phytoplankton) production.
The outbreaks of certain species of dinoflagellates and diatoms (e. g., Gymnodinium breve, Nitzschia pungens, Pseudonitzschia pseudodelicatissima) have coincided with El Niño events. These outbreaks suggest that the warming trends generated by global climate change in conjunction with eutrophication of anthropogenic origin can stimulate the growth of these species and broaden or change their range. However, El Niño is not the only factor contributing to rising sea temperatures; global warming has probably contributed to variations in sea surface temperature (Burkholder, 1998).
A study carried out by Coronado (2015) using satellite images, found that La Niña events could favor the presence of algal blooms in the Colombian Caribbean (the Gulf of Urabá, the mouth of the Magdalena River and the Guajira Peninsula) in greater proportion than the El Niño. The author mentions that during the rainy season a greater influx of nutrients can occur. Another study in the Colombian Pacific between 1995 and 1997 found that the decrease in the abundance of diatoms and the increase of dinoflagellates, due to an increase in water temperature, is related to the development of a warm event such as El Niño (Medina, 1997).
Calvo and colleagues (2014) evaluated the abundance of dinoflagellate and diatom species during two La Niña events (2008 and 2010) and one El Niño event (2009) in the Gulf of Nicoya (Costa Rica). The study showed, in general, an increase in the abundance of dinoflagellates and diatoms during La Niña events and a decrease of these groups during El Niño event, however, some species ofdinoflagellates like Prorocentrum micans and Tripos furca showed an opposite behavior, reaching its maximum peaks during the El Niño event.
A similar study on the abundance of marine phytoplankton in the Colombian Pacific during El Niño and La Niña events was led by Rojas and Ortiz (2007). The study found that the abundance of dinoflagellates has a different behavior compared to diatoms because it tends to be higher under El Niño conditions than under La Niña conditions in both coastal and oceanic regions. Another research conducted by Gingold and colleagues (2014) showed that both the increase in the frequency of tropical storms and in sea surface temperature generated by climate change are positively related to the poisonings caused by ciguatera, which is generated by an increase in the abundance of toxic dinoflagellates, mainly those belonging to the genus Gambierdiscus. Likewise, Lavaniegos et al. (2003) and Tapia (2007) observed increases in diatom concentrations during El Niño events in 1997-1998 and 1999-2000, respectively.
The results found on Isla de Barú suggest that large-scale atmospheric and oceanographic processes such as El Niño, may modulate dinoflagellate populations, increasing abundance compared to neutral periods, especially in Prorocentrum species.
CONCLUSIONS
Three genera of epiphytic dinoflagellates and ten genera of epiphytic diatoms were found on Isla de Barú. Prorocentrum, Ostreopsis, and Gambierdiscus contain species that are related to toxic events as ciguatera and diarrhetic shellfish poisoning. Prorocentrum lima was the most abundant dinoflagellate found in this study.
The results found on Isla de Barú suggest that El Niño may modulate dinoflagellate populations, increasing its abundance compared to neutral periods, especially in species of the genus Prorocentrum. However, further study is necessary to understand better how events such as El Niño affect these populations


