











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
According to the World Health Organization (WHO, 2018), cancer is the second leading cause of death globally, responsible for 8.8 million deaths in 2015, 70 % of them in low- and middle-income countries. While great advances have been made in the last decades in several types of cancer treatment and prevention, it continues to be necessary to find new approaches and to look for new substances that can be used in the early diagnoses and the treatment of neoplasms.
Several natural and synthetic compounds have been found, which could be candidates for the development of new drugs. In them, it is important to know their structure and biochemical classification, the potential of their biological activity, toxicity, specificity, action pathways, and kinetic parameters. It should be noted that in some cases, compounds of natural origin obtained from poisonous animals, plants or bacteria, provide more often a clearer idea about the molecular structures and the routes of action involved in the anticancer activity (Narang and Desai, 2009). The importance of these at the biological level is that they are better tolerated by the organism and interact optimally with bio-macromolecules. Some natural substances can inhibit angiogenesis; a process that involves the formation of new blood vessels and that is considered essential for the growth and spread of the tumor since it reinforces the supply of oxygen and nutrients in the cells, whose metabolic profile requires large amounts of glucose due to the increase in the activity of glycolytic routes (Sun et al., 2011; Simons et al., 2012). Others block the synthesis of proteins; the increase in protein production is related to the initiation and progression of cancer, since it is a necessary process for the maintenance of cell viability and reproduction due to the role of proteins in the architecture, regulation, and execution of intra and intercellular processes (Novac et al., 2004; Martineau et al., 2014). There are also substances that participate in the induction of cell death, through organized mechanisms such as apoptosis; where macrophages intervene that are responsible for the elimination of cell bodies, or others that involve pro-inflammatory factors, mainly due to the exposure of intracellular content in the extracellular matrix; as is the case of necrosis (Cascales, 2003; Lizarbe, 2007).
Arthropod venoms are precisely the type of substances that have a remarkable potential as antineoplastic and anti-infective agents. We believe that further studies are needed in the identification and isolation of the active antitumoral compounds present in venoms and there should be further attempts to elucidate their mechanism of action. In this review, we present an overview of studies on the antitumoral activity of arthropod venoms.
Several active compounds have been obtained from Arachnid venoms. Proteins such as latarcin 2a obtained from Lachesana tarabaevi (Vorontsova et al., 2011) or gomesin found in Acanthoscurria gomesiana (Soletti et al., 2010), as well as their synthetic analogs, have shown cytotoxicity in vitro against tumor cell lines. Findings included proton gradient imbalance at the mitochondrial membrane, modulation of calcium, potassium and sodium ion channels, pore formation, mitochondrial inactivation, cell necrosis, apoptosis independent of induction of membrane phospholipids symmetry by externalization of phosphatidyl serine, Reactive Oxygen Species (ROS) formation, receptor activation of cell death all of which lead to cell morphological alterations (De Souza et al., 2015).
Other very interesting arachnids, albeit much less studied, are scorpions. Their venom is rich in mucopolysacharides, proteins, and peptides (Possani and Rodríguez, 2006). There are reports that show that some species whole venom or molecules isolated from it inhibit cell growth or induce tumor cell death through mechanisms that include ionic channel blockage (Wang and Ji, 2005), disruption of membrane integrity (Das Gupta et al., 2007), mitochondrial damage, DNA degradation (Gupta et al., 2010), polysaccharides fragmentation (Feng et al., 2008) and immune response modulation (Yang et al., 2000).
The order Scorpionida goes back some 400 million years to the Silurian era. They found they are widely distributed in all continents between 50° N latitude and 50° S latitude. Some 20 families have been described, with approximately 208 genus and 2231 species. However, they are more abundant in the intertropical regions (Brownell and Polis, 2001; Chippaux and Goyffon, 2008; Ortiz et al., 2015; Santibáñez et al., 2016).
The family Buthidae that includes the genera Androctonus, Buthus, Buthotus, Leirus, Mesobuthus y Parabuthus found in Asia and Africa and Centruroides y Tityus found in the Americas has been the subject of great interest in toxicology and medicine due to the pharmacological potential of their venom (Fernandes et al., 2013). The venom contains insect-specific toxins that can interact with a great affinity for vertebrate cells causing a variety of pharmacological responses that make them potentially dangerous for human beings (Brownell and Polis, 2001; Chippaux and Goyffon, 2008). It is believed that only 5 % of the venom's dry weight is made up of peptide compounds (BenNasr et al., 2013). But only 0.02 % may interact with sodium, potassium, calcium, and chloride ionic channel proteins (Possani et al., 2000; Possani and Rodríguez, 2006), particularly those in excitable muscle and nerve cells, accounting for the principal toxic effects of the venom in mammals (BenNasr et al., 2013). It is possible that these toxins represent a survival tool as their injection can cause paralysis both in the scorpion predators and possible preys (Possani and Rodríguez, 2006; Almaaytah et al., 2012). Other proteins found are proteolytic, lipolytic and hydrolases enzymes, particularly hyaluronidase and phospholipase, which even though presenting very low activities, they might contribute with the penetrations and diffusion of the venom to different tissues. The other fraction of the venom's dry weight is made up of a complex mixture of mucopolysaccharides, mucoproteins, free amino acids, nucleotides, biogenic amines, serotonin, histamine, histamine releasers, inorganic salts, carbohydrates, lipids, enzyme inhibitors and other non-identified substances without known biological activity (Heinen and Da Veiga, 2011; BenNasr et al., 2013; Santibáñez et al., 2016).
In the literature it is usual to find the components of the venom classified according to their peptide structure in non disulfide bridges peptides (NDBPs), disulfide bridges peptides (DBPs) and peptides with particular structures such as thio-ether rings and proline or glycine amino acids (Brownell and Polis, 2001; Dai et al., 2002; Nabi et al., 2014; Luna et al., 2016). However, it is also possible to group them following their mechanism of action or molecular target in toxic peptides that represent neurotoxins, cytolytic or host defense peptides (HDPs) and enzymatic or non-toxic peptides (Béchohra et al., 2016; Santibáñez et al., 2016; Romero et al., 2017).
HDPs
Essentially they are broad-spectrum antimicrobials (AMPs) and thanks to this nature they are linked to the innate response of the specimen to pathogenic microorganisms in the absence of an adaptive immune system. They are widely diversified into arthropods, exhibiting different constitution and mechanism of action with low resistance. Parts of this group are peptides rich in cysteines, such as NDBPs, defensins, scorpine-like peptides and recently Waprins molecules frequently found in snake venoms among others (Santibáñez et al., 2016; Luna et al., 2017; Romero et al., 2017).
NDBPs
NDBP are usually low-weight peptides, between 13 to 60 residues prevailing basic amino acids, with more positively charged than neutral or negatively charged, giving the peptide a net positive charge. They have a secondary helical structure which can be arranged in three different ways: one α-helix with two flexible random coil ends, two α-helixes separated by a central random coil and a 100 % coil-coiled structure (Almaaytah and Albalas, 2014). Since they are amphipathic, they can easily interact with negatively charged structures such as membrane phospholipid heads through electrostatic forces changing their conformation and thus penetrating the membrane (Du et al., 2015). In the presence of homologous peptides, they can undergo oligomerization creating pore-like structures that alter membrane stability. The result of these interactions peptide-membrane is cell lysis and induction of other forms of cell death, such as mitochondrial apoptosis (Harrison et al., 2014; Satitmanwiwat et al., 2016). It has been proposed that this is also a means to improve the DBPs action (Almaaytah et al., 2012). It is interesting to note that peptides similar to NDBPs have been found in a wide sample of organisms, including bacteria, fungi, plants and animals where they might play a similar role (Riedl et al., 2011). Since they don't have a target organ, their biological effects are of different kinds: some peptides have cytolytic, antibacterial, antifungal, antimalaric, insecticide, anti-inflammatory, and anti-cancerogenous activity while others can act directly on the endothelium potentiating the action bradykinin or acting as immunomodulators (Almaaytah et al., 2012; Ortiz et al., 2015).
Ramírez et al. to compare the bioactivity of natural and improved AMPs by peptide engineering of the venom of the scorpions Urodacus yaschenkoi and U. Manicatus. During the modifications, positive results were obtained in the account maintain the net positive charge, increase the hydrophobicity of the hydrophobic region and the hydrophilicity of the hydrophilic region. They noted that by concentrating the positive charges at the end of the peptide together with the addition of bulky side chains, they improved the affinity and selectivity towards the bacterial membranes. Likewise, the amino acid substitution of stereochemistry L by D decreased the development of hemolytic activity (Romero et al., 2017).
Defensins
They are antimicrobial peptides structurally like the potassium channel blockers. Defensins are rich in cysteine, in general, have three disulfide bridges, are amphipathic, cationic, with an α-helix and antiparallel β sheets (Zhu et al., 2000; Ganz, 2003; Ghavami et al., 2009). They are the first line of native immune response in plants, fungi, and animals promoting the formation ofmembrane pores in pathological microorganisms. Other effects found are reduction in membrane potential, increased membrane permeability, decrease in ATP levels, increase of ROS (Cociancich et al., 1993). Membrane permeabilization is crucial to cell death and depends on the electrostatic forces generated between the negatively charged membrane and the positively charged defensins (Ganz, 2003).
In his research Meng et al., (2016) suggest the existence of a functional link between defensins and potassium channel blocking toxins, which could suggest a common evolutionary ancestor. Genomic, proteomic and transcriptomic analysis of the BmKDfsin4 defensin of Mesobuthus martensii was carried out, as well as its three-dimensional structure by circular dichroism spectroscopy (CD). Based on the results obtained, it was concluded that the presence of basic residues in the molecule conferred association capacity with the acid residues that make up the potassium channels, especially Kv1.3 (Meng et al., 2016).
Some defensins show a greater affinity for the more negatively charged tumor membranes, which may provide more union sites than normal cell membranes (Guzmán et al., 2015). A preliminary study in which mammary carcinoma MCF-7 cells were exposed to venom from six different scorpion species suggests the presence of defensin-like proteins in the most cytotoxic venoms (Estrada et al., 2017).
Scorpions and Waprins
Postulated as hybrids between cecropins and defensins, scorpions, isolated for the first time from the scorpion Pandinus imperator, have a dual role that can block potassium channels and act as antimicrobials or cytolytic. Due to its high antiparasitic activity, it would be a good candidate for the development of antimalarial therapies (Zhu and Tytgat, 2004; Santibáñez et al., 2016).
DBP PEPTIDES
DBP is more specific when binding to their target structures, mostly voltage-dependent ionic channels. Their secondary structure comprises an α-helix adjacent to a double or triple antiparallel chain β-sheet stabilized by four disulfide bridges for those acting on sodium and chloride channels and three bridges for those acting on calcium and potassium channels. They have two positive basic amino acid poles (Brownell and Polis, 2001; Ding et al., 2014). They have a net positive charge that is related to their toxicity level since several studies have shown that this type of charge improves both recognition and binding affinity of peptide and binding site, the latter usually negatively charged (Brownell and Polis, 2001). They can be long or short chain peptide. Long chain peptides are 58 to 76 amino acid residues, while short chain peptides range between 20 and 43 amino acid residues (Possani and Rodríguez, 2006). Long chain peptides usually modify voltage-dependent sodium channels; a transmembrane proteins family that allows the movement of ions in and out of the cell in response to changes in membrane voltage. They have three subunits, one α or pore formation and two auxiliaries β. The β subunit has four six transmembrane segments domains (Brownell and Polis, 2001; Catterall, 2014).
Long chain peptides can be classified as α or β toxins depending on the electrophysiological effect on the channel (Ortiz et al., 2015). α-toxins delay channel inactivation which is dose-dependent: at high concentrations the action potential generated in the excitable cell is more prolonged, inducing paralysis in muscle cells and arrhythmias in cardiac cells. If the concentration is low, α-toxin causes a strong depolarization followed by a falling out in excitability. They have been found mainly in old world scorpions. β-toxins are found in New World scorpions. They trigger channel opening at more negative potentials, which may unleash myoclonic and spastic muscle response that changes according to tissue type (Nabi et al., 2014; Ortiz et al., 2015).
Short chain peptides, on the other hand, are involved in some potassium channels dysfunction (Heinen and Da Veiga, 2011), which are another type of transmembrane proteins that share structural and functional homologies with sodium channels (Brownell and Polis, 2001). Binding of the short chain peptide to the channels induces channel blockage or reduces the time it remains open, thus causing difficulties in repolarization and neural transmission (Possani and Rodríguez, 2006). Some short chain toxins can also block chloride and even calcium channels through mechanisms that are not presently clear (Heinen and Da Veiga, 2011; Ortiz et al., 2015). Some of the above mechanisms are still under study, and it is possible that some unspecific toxins might not require specific protein receptors (Martineau et al., 2014).
POTENTIAL ANTITUMORAL EFFECT OF SCORPION VENOM
There is some evidence that many scorpion peptides might be useful in the treatment of different types of cancer, given their ability to alter membrane permeability or to selectively bind to certain receptor domains inducing cell death or growth inhibition through several signal cascades (Heinen and Da Veiga, 2011; Baby and Jency, 2012). They can also modify cancer cells local environment turning it more hostile for survival through mechanisms such as angiogenesis inhibition (Sun et al., 2011; Ortiz et al., 2015). Their affinity and specificity can be so great that molecular probes derived from scorpion venom have recently been designed to localize and study ionic channels (Possani and Rodríguez, 2006). Since expression of ionic channel proteins is altered in some types of cancer, molecules affecting those structures become the subject of research studies (Ding et al., 2014; Nabi et al., 2014). Several of these peptides with the capability to seriously alter specific cancer cells growth have been identified (Liu et al, 2012). It seems that sodium channels are overexpressed in several human cancers and seem to correlate cellular function and metastatic processes (BenNasr et al., 2013; Ding et al., 2014).
BMK-AGAP, a peptide isolated from the scorpion M. martensii and other structurally similar peptides are α-toxins which are currently in use bound to fluorescent compounds to delimit gliomas in animal models. They exhibit analgesic activity, as well as inducing growth inhibition and triggering apoptosis processes without affecting normal cells (BenNasr et al., 2013; Ding et al., 2014).
Other peptides isolated from scorpions include BmTx3, from M. martensii, AmmTx3 from Androctonus mauritanicus, Bekm-1 from Mesobuthus eupeus (Liu et al., 2012; BenNasr et al., 2013), Margatoxin (MgTX) from Centruroides margaritatus , Iberiotoxin (IbTx) from Mesobuthus tamulus (Buthidae) and Charybdotoxin (ChTX) from Leiurus quinquestriatus hebraeus, (BenNasr et al., 2013; Ding et al., 2014; Ortiz et al., 2015). All of them have valuable properties. Human lung adenocarcinoma cell lines (A549) exposed to MgTX show a reduction in tumor volume (Jang et al., 2011). IbTx inhibits growth and induces cell death glioma and prostate cancer PC-3 (Weaver et al., 2004; Bloch et al., 2007). ChTX detains murine fibroblasts NIH3T and human melanoma cells migration (Schwab et al., 1999; Ortiz et al., 2015).
Imperatoxin A (IpTxA) from P. imperator, was the first toxin known to bind specifically to ryanodine receptors, proteins involved in intracellular calcium transport. Both IpTxA and its homolog, maurocalcine (MCA) from Scorpio maurus palmatus, have been used as transporters of antineoplastic drugs in human mammary carcinoma MCF-7 and MDA-MB231 cell lines taking advantage of the ease they have to move through membranes. These biomolecules are very useful in the development of new drugs that reduce current resistance levels to antitumoral drugs (Howl et al., 2007; Aroui et al., 2009; Ortiz et al., 2015).
One ofthe most projected toxins in the treatment ofcancer is Chlorotoxin (CTX or CLtx) obtained from L. quiquestraiatus hebraeus venom. This molecule selectively blocks chloride channels in insects. However, it has recently shown that it also acts on voltage-dependent chloride channels, calcium-dependent phospholipid-binding proteins (Annexin-2) and matrix metalloproteinases-2 (MMP-2) in gliomas. These structures are necessary to preserve mobility and cell viability (Ullrich and Sontheimer, 1996; Cohen et al., 2018).
Although the peptide does not have a direct action on the chloride channel, it inhibits the surface of a proteinase intimately ligated to the channel activity, thus blocking chloride ion flow through the channel structure (Deshane et al., 2003; Gomes et al., 2010). CTX and its bioconjugates have undergone phase I and II clinical trials with the hope of differentiating molecular profiles and radiological characteristics of glioblastoma multiforme (GMB), a breakthrough in the generation of theranic agents (Cohen et al., 2018). Its use has also been profiled towards the detection and treatment of colon cancer, skin, lungs, mammary glands, stomach, pancreas, cervix, and colon cancer. One very important feature of CTX treatment is that it has little or no effect on normal cells (Mamelak and Jacoby, 2007; Heinen and Da Veiga, 2011).
BmKCTa, a toxin extracted from M. martensii, shares about 68 % structural homology with CTX and presents similar effects. The recombinant peptide, rBmKCTA has shown activity in reducing human glioma and lymphoma cells proliferation, probably by inducing apoptotic processes (Fu et al., 2007).
There are other scorpion toxins whose activity do not involve ionic channels but that, nevertheless, have good potential as anticancerigenic drugs: Bengalin, obtained from Heterometrus bengalensis is an anti proliferative and pro-apoptotic protein that acts on human U937 and K562 leukemic cells with minimal toxicity on normal lymphocytes. It seems that it provokes changes in mitochondrial membrane potential, inhibits heat shock proteins HSP79 and HSP90 and promotes autophagic processes (Chippaux and Goyffon, 2008).
ANTP is another peptide obtained from M. martensii whose effect is probably due to its union to heat shock proteins. The toxin has been studied in animals inoculated with fibrosarcoma S-180 and Ehrlich ascites cells. A hybrid toxin, ANTP-TPR, was effective against not only the above-mentioned cell lines but other ones such as mammary, lung and prostate carcinoma (Liu et al., 2002).
Other scorpion venom peptides may activate proteins directly involved in cell death. Neopladine 1 and Neopladine 2 isolated from Tityus discrepans, venom are pro apoptotic molecules in human mammary carcinoma SKBR3 cells. Their action involves up-regulation of mitochondrial Bcl-2 and FasL cell death receptor expression. These two toxins have also been involved in necrotic processes (D'Suze et al., 2010).
Hyaluronidase is an evolutionarily conserved enzyme, found in most animal poisons. With a weight between 4550 kDa it catalyzes the degradation of hyaluronic acid (HA), a complex carbohydrate present in the extracellular matrix, making tissues more permeable and up to 20 times the diffusion rate of other toxins (Bordon et al., 2015; Díaz et al., 2015). Different investigations indicate that it is one of the main components of the venom of arthropods including scorpions. Hyaluronidase BmHYA1 found in M. martensii reduced human mammary carcinoma MDA-MB-231 growth. This very aggressive cancer line expresses large amounts of hyaluronic acid (Shuster et al., 2002). Serine proteinases are typically present in some invasive cancer cell lines. An inhibitor of serine proteinases, BmAP1, has been isolated from the same scorpion venom.
Interestingly, serine proteinase BmK-CBP was also found in the venom. This enzyme shows a great capacity to bind specifically to cells from the MCF-7 cell line. This property could be used for transport of anti-cancer drugs to this tumor (Gao et al., 2008).
The above peptides are disulfide bridged peptides (DBP). In contrast to this class of scorpion peptides, the nondisulfide bridged peptides (NDBPs) show little antitumoral activity. Only three of them have been shown to have any activity against tumors. TsAP1 and TsAP2 isolated from Tityus serrulatus have good antimicrobial activity and show some effect against tumor cells.
The design and synthesis of analogous peptides with better anti-tumoral effectiveness have shown promising results. These analog compounds have shown to be more effective against oral epidermoid carcinoma cells H157, lung adenocarcinoma H838, mammary carcinoma MCF-7, prostatic carcinoma PC3 and glioblastoma U251-MG (Guo et al., 2013). Mauroporin, isolated from A.mauritanicus venom, seems to exert cytotoxic and anti proliferative action in different cell lines of prostate cancer (Almaaytah et al., 2013). It seems that these peptides mechanism of action could involve induction of cellular death due to their structure and topological properties, e.g., the high negative surface charge and the presence of microvilli or pseudopods that increase the total cellular area favors increased peptide linkage to membrane proteins and proteolipids, thus affecting membrane integrity (Lohner and Blondelle, 2005; Zargan et al. , 2011a).
Other reports have shown that complete venom from some scorpions is a good cytotoxic agent. It is probable that several of the substances mentioned above play a synergic role potentiating the harmful effects on cancer cells. Exposition of human mammary carcinoma cells to M. tamulus venom significantly decreased their viability. Odontobuthus doriae venom inhibits human neuroblastoma SH-SY5Y and mammary carcinoma MCF-7 cell growth and induces apoptosis, probably to increase of intracellular oxidative stress (Zargan et al., 2011a; b). Androctonus crassicauda's venom has been used against these same cell lines with similar results (Zargan et al., 2011c).
The venom of the Cuban blue scorpion Junceus Rhopalurus has shown selective cytotoxic activity in epithelial cancer cells, likewise, induced apoptosis in the triple-negative human breast cancer cell line MDA-MB-231 through mitochondrial pathways. There was a synergistic effect of this poison combined with low concentrations of cytostatic fluorouracil, doxorubicin, and cisplatin on HeLa cervical cancer cells (Díaz et al., 2013; Díaz et al., 2017; Yglesias et al., 2019). However, Giovannini et al., (2017) when analyzing the effect of a preparation based on this poison in hepatocellular carcinoma (HCC), they found that it induced cell proliferation and improved the expression of the vascular endothelial growth factor receptor (VEGFR) associated with angiogenesis and metastasis capacity. Suggesting good research in molecular biology for these poisons and use them with caution (Giovannini et al., 2017).
NEW INSIGHTS
Thanks to technological advances in high-throughput fractionation methods, molecular biology, bioinformatics and the emergence of omic disciplines, different genes have been studied and analyzed, as well as the product of their expression, using a minimum number of copies of the kind of interest. Table 1 shows the sequences of some components with anticancer activity, isolated from different scorpion species.
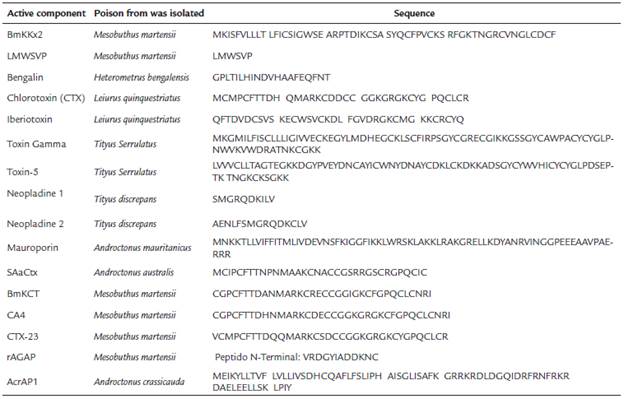
The discernment of the structure of some components found in the poisonous gland of scorpions in association with the effects produced after the bite, has allowed to have a deeper insight into its mechanism of action evaluated in different cellular models, from which designed and tested compounds improved by recombination or biosynthesis (Fernendes et al., 2013).
For example, SAaCtx, CTX and BmKCT, effective in the glioma and coming from different genera of the Buthidae family (Figure 1a), have the same mechanism of action and structural homology, the same happens with the peptides CA4 and CTX-23 (Figure 1b) and between Neopladin 1 and 2 (Figure 1c). Now, CTX and CTX-23, although they have a high structural similarity, have not elucidated that their mechanism of action is the same (Figure 2) (Xu et al., 2016).


The obtaining of transcriptomes from mRNA by techniques such as RNA-seq provides useful information at low cost in the construction and strengthening of cDNA libraries, where genes associated with biological processes, cellular components or molecular functions can be identified, as well as their phylogenetic evolution.
Santibáñez et al., (2016) they highlighted some differences between the venom of Buthidos and non-Buthidos such as Superstitionia donensis when performing a transcriptomic and proteomic analysis for this species by RNA-seq. We found toxins related to ion channels of the scorpines and calcins type, long-chain peptides La1-Like of unknown function, and protease inhibitors, in more abundant amounts than in Buthides (Santibáñez et al., 2016).
Studies conducted by Romero et al. they allowed to know the biodiversity of the NDBPs in the venom of the Mexican scorpion Thorellius atrox. A sequence encoding a peptide with action in the voltage-dependent calcium channels, previously unknown in scorpion venoms, in addition to HDPs, was found (Romero et al., 2017). Another study of the transcriptome of the venom glands of Serradigitus gertschi allowed to know 119 sequences of peptides and proteins with diverse functions; enzymes, ion channel toxins, HDPs, protease inhibitors among others. The findings in both scorpions belonging to the Vaejovidae family demonstrate the existence of anticrobic peptides (Romero et al., 2018). Additionally, transcriptomics allowed Li and his collaborators to identify 238 new peptides in scorpion poisons Androctonus mauritanicus and Androctonus australis, from which they discovered an anticancer peptide called Gonearrestide that inhibits the growth of colon cancer cells (Li et al., 2018). This shows the usefulness of these tools in the discovery, expression, and characterization of peptides and proteins present in scorpion venom, as an initial basis for the modification and synthesis of molecules with anticancer or antimicrobial potential.
CONCLUSIONS
Several toxins found in scorpion venom might be useful to design drugs, tumoral markers, and adjuvants to cancer treatment taking advantage of their high affinity and specificity for certain ionic channels over expressed in many malignant cells. Other substances found in the venom such as peptides, proteins, enzymes, AMP, DBPs, NDBPS, and defensins induce cell death by different mechanisms, including membrane pore formation or activation of proapoptotic signal cascades. It should be taken into account that these same substances can have other applications, e.g., treatment of infectious diseases taking advantage of their wide spectrum and low resistance that they could generate.


