











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Coral reefs are one of the most important, diverse and productive ecosystems on the planet. They thrive in clear waters of tropical seas and are fundamental in the development of a very complex ecosystem. Coral reefs provide substantial economic and ecological services to society and the environment. The reef structure protects coastal areas, mangroves and seagrasses from erosion (Moberg and Folke, 1999; Ritson-Williams et al., 2009). The reef structure also provides habitat for many organisms including species of commercial interest such as crabs, lobsters and mollusks (Jennings and Polunin, 1996; Birkeland, 1997; Mumby and Steneck, 2008).
Competition determines the structure and composition of coral reefs communities (Birkeland, 1997). In coral reefs, the increasing competition between algae and corals has become an important factor for reef resilience (reviewed in Puyana, 2018). The process of coral reef recovery is difficult because juvenile corals must settle in substrates now dominated by algae and cyanobacteria which are very effective colonizers after disturbances (McCook et al., 2001; Hughes et al., 2007; Diaz-Pulido et al., 2010). The dominance of algae and cyanobacteria can become a serious bottleneck for the settlement and recruitment of corals, depending on the local and dominant characteristics of algal communities (McCook et al., 2001; Diaz-Pulido et al., 2010).
Benthic mats of algae and/or cyanobacteria occupy available substrate but also form physical barriers hampering coral recruitment (McCook et al., 2001; Diaz-Pulido et al., 2010; Ford et al., 2018). Experimental evidence has shown that cyanobacterial mats rely on chemically mediated effects affecting the settlement, survival, and recruitment of coral larvae (Kuffner et al., 2004; Kuffner et al., 2006).
Hence allelopathic mechanisms may play significant roles in the processes of competition between coral reef benthic organisms, impacting reproductive processes, physiology, growth, and survival of reef communities (Thacker et al., 1998; Chaves Fonnegra et al., 2008; Chadwick and Morrow, 2011).
Allelopathy is the process by which an organism produces chemical compounds that negatively affect the growth, development, health, behavior, and reproduction or population biology of another species. Allelopathy has been very difficult to demonstrate in aquatic environments due to the hydrodynamic character and the complexity of communities in those environments. This mechanism may be frequent in sessile organisms that compete for resources such as space or access to light including algae and sponges, among others (De Nys et al., 1991; Littler and Littler, 1997; Steinberg et al., 1998; Engel and Pawlik, 2000; Richelle-Maurer et al., 2003; Titlyanov et al., 2007; Chaves-Fonnegra et al., 2008).
Marine cyanobacteria produce a large number of biologically active compounds, important for their survival (Nagle and Paul, 1998; Tan and Goh, 2009; Engene et al., 2011; Leão et al., 2012), for the establishment and persistence of cyanobacterial blooms (Leão et al., 2012) and as inhibitors of potential competitors such as algae, microalgae, aquatic plants and even other species of cyanobacteria (Jüttner et al., 2001; Berry et al., 2008).
Currently, cyanobacteria are a frequent sight in several reef locations, although reef ecologists still consider algae and cyanobacteria as the same category when performing field surveys. Cyanobacterial mats form multispecies consortia and grow on other organisms such as corals and octocorals causing polyp retraction, bleaching or the death of coral tissue under cyanobacterial mats or in areas where mats come in direct contact with corals (Paul et al., 2005; Smith et al., 2008; Puyana and Prato, 2013; Puyana et al., 2015; Ford et al., 2018) (Figure 1).

Figure 1 a-b Cyanobacterial mats overgrowing soft corals of the genus Antillogorgia. Soft coral tissue under cyanobacterial mats was dead and disintegrated upon contact leaving behind the gorgonin matrix which was rapidly fouled. c. Cyanobacterial mat overgrowing corals and macroalgae. d. Cyanobacterial "lumps" overgrowing corals and coralline algae. All pictures taken in Old Providence Island by Julian Prato, (2013).
MATERIALS AND METHODS
Study Area
Field assays were conducted at Isla Grande, in the archipelago of Islas del Rosario (10°11' N, 75°48' W). The archipelago is a set of low-lying islands surrounded by extensive coral formations, located 54 km to the southwest of the Bay of Cartagena, Colombian Caribbean (Figure 2). The area, comprising approximately 120 km2, is part of the Parque Nacional Natural Corales del Rosario and San Bernardo (PNNCRSB). The area is located within the area of intertropical convergence (ITCZ), under the constant influence of winds from the Northeast and Southeast. The annual average sea surface temperature is 27.5 °C, with an annual rainfall of approximately 1013 mm and salinity close to 35. Tidal variation in the area is low, ranging between 17 and 58 cm (Sánchez et al., 1999).
Sample collection and identification
The Agencia Nacional de Licencias Ambientales (ANLA) and Ministerio de Ambiente y Desarrollo Sostenible granted permission to Universidad Nacional de Colombia to collect samples and perform this research (Permission 4 of 10/02/2010, Anexo 2, Contrato de Acceso a Recurso Genético 108).
Biological material for cyanobacterial identification and organic extract preparation was collected at San Andres. Old Providence Island and Islas del Rosario between 2012 and 2014. Samples of cyanobacteria were manually removed by SCUBA and introduced in bags of very fine mesh. Once at the surface, samples were transferred to ZiplockTM bags and frozen. A small portion of the sample (approximately 15 g), was preserved in a solution of 4 % formalin in seawater in order to perform taxonomical analyses. Cyanobacterial identification was carried out with a Nikon microscope coupled to a computer with the NIS-Elements Br 2.30 Imaging software. The most important characters for taxonomic identification included cell shape, length, and width, the presence of mucilaginous sheath covering the filaments and shape offilament terminals following Komarek and Anagnostidis, (2005).
Extract preparation
Frozen samples of cyanobacteria were lyophilized to then subjected to extraction with a 1:1 mixture of methanol-dichloromethane. Extracts were concentrated under vacuum evaporation, weighed and kept frozen until they were used. Small quantities of each extract were placed in individual vials and weighed. Once in the laboratory and prior to each assay, extracts were fully resuspended in very small volumes of ethanol-acetone (<100 (mL), vigorously shaken and the mixed with seawater to the final test concentration. The ethanol-acetone mixture fully solubilized extracts so no filtration was required.
Lethality assay against
Artemia salina
The lethality assay against Artemia salina is widely used to evaluate the toxicity of organic extracts and for the screening of potentially bioactive extracts and fractions (Sam, 1993). For the assay, cysts of Artemia were incubated, until hatching, in artificial seawater (3.8 % sea salt) at a temperature between 26-30 °C. For the assay, 15 nauplii were placed in each well of 24 well plates filled (1.5 mL) with filtered seawater. Artemia nauplii were then exposed to different concentrations of cyanobacterial extracts resuspended in ethanol-acetone (< 100 (mL) and seawater. For this assay, we evaluated extracts 32, 36, 38 y 42 at concentrations of 1000, 500, 100 and 10 mg mL-1 in triplicate. Negative controls consisted of pure seawater and seawater with the same amount of the ethanol-acetone mixture used to fully resuspend the extracts. A 100 (mg mL-1 potassium dichromate solution was used as positive control. The number of surviving nauplii was assessed 24 hours after addition of the extracts. The LC50 was determined by the Reed-Muench graphical method following Sam, (1993).
Lethality assay against fertilized coral eggs (embryos) of the scleractinian coral Orbicella annularis (Ellis and Solander, 1786)
We evaluated the effect of organic extracts of marine cyanobacteria against fertilized coral eggs (embryos) of the scleractinian coral Orbicella annularis in order to assess the potentially harmful effects of extracts of these microorganisms in the embryos' survival. The coral O. annularis was selected because it is an important reef-building species in the Colombian Caribbean (Reyes et al., 2010). Additionally, the gametogenic cycle and date of the mass spawnings of this species is well known, facilitating the successful collection of their larvae (Sánchez et al., 1999; Alvarado and Acosta, 2009).
Embryos of Orbicella annularis were collected with gamete traps during the mass spawning that took place in September 2014 at Isla Grande, Rosario Islands. The design of gamete traps and collection method is thoroughly described in Alvarado and Acosta, (2009). Right after spawning, gamete collectors were sealed and carefully brought to the surface to be quickly transported to the laboratory at CEINER, Centro de Investigación, Educación y Recreación de San Martin de Pajarales, Rosario Islands. Once in the laboratory, the suspension of gametes in seawater was filtered to remove predators such as copepods and other planktonic invertebrates. Subsequently, a cross-fertilization of eggs was performed. In order to achieve so, gamete packets were shaken to break them and male and female gametes were separated by means of special filters. The male gametes of one colony were used to fertilize the eggs obtained from another colony. Eight hours later, fertilized and unfertilized eggs were manually separated using glass capillaries under a stereoscope. Fertilized eggs had a pink reddish coloration and an oval shape. They were also active. Once dead, the embryos lost their shape and smooth texture. All successful embryos were transferred to plastic containers with fresh, filtered seawater. Throughout this process and during the whole experiment temperature was kept at 28 °C and a salinity of 35.
The embryo lethality assessment started about 14 hours after fertilization. For the assay, ten embryos were placed in each well of 24 well plates filled (1.5 mL) with filtered seawater. Embryos were then exposed to different concentrations of cyanobacterial extracts resuspended in ethanol (< 100 (mL) and seawater. For this assay, we evaluated extracts 32, 36, 38 and 42 at concentrations of 1000, 500, 100 and 10 (g mL-1 in triplicate. Controls consisted of plain seawater and seawater with the same amount of ethanol used to resuspend each extract at the different concentrations tested. Embryo mortality was assessed 6, 18 and 24 hours after exposure to the extracts and only when embryo survival in the seawater control was greater than 75 % (Fearon and Cameron, 1997).
Polyp retraction assay against fragments of the scleractinian coral Madracis mirabilis (=auretenra) (Locke, Weil and Coates, 2007).
Corals are highly sensitive to exogenous compounds and quickly retract their polyps if environmental conditions are not favorable. A rapid and sensitive way to evaluate an indirect potential allelopathic role of organic compounds is the evaluation and extent of coral polyp retraction and subsequent recovery upon exposure to the substance ofinterest (Petritchtcheva et al., 2002; Richelle-Maurer et al., 2003).
The polyp retraction assay was carried out with fragments of the yellow pencil coral Madracis mirabilis (=auretenra). This species has a wide distribution in the Colombian Caribbean , has a great capacity for tissue regeneration, low susceptibility to diseases and easily reproduces asexually by fragmentation (Reyes et al., 2010). Coral fragments were collected manually by SCUBA in the fringing reefs North of Isla Grande, Islas del Rosario in November 2014. They were stored in airtight plastic bags, brought to the surface and immediately transferred to 40 L containers. Once at the CEINER facilities the fragments were placed in 4000 L tanks with filtered seawater, natural lighting, and continuous recirculation. Each fragment was attached to small individual ceramic tiles using an epoxy resin to maintain a vertical position. Corals were acclimatized for three days and fed with freshly hatched Artemia salina nauplii (Petritchtcheva et al., 2002).
For the assay, each coral fragment was introduced in 600 mL glass containers. Once polyps where fully extended, cyanobacterial extracts were resuspended in an ethanol-acetone mixture (< 100 mL), mixed with seawater to reach the desired concentration and then added to each container. Extracts evaluated were 32 and 38 and were selected due to their great local abundance and high toxicity against A. salina.
Each extract was tested at a low concentration (2 mg mL-1) and a high concentration (11 (mg mL-1). These concentrations were selected prior to the experiment in order to detect coral responses in an adequate time frame. Two controls were used. The same volume of the ethanol-acetone mixture used to resuspend the extracts and an 11 |mg mL-1 solution of table sugar (sucrose). The sugar solution was used to verify that the addition of a "harmless" organic compound did not generate adverse effects on the corals. All experiments were run with three to five replicates. Once the extracts were added, individual coral fragments were monitored at 10, 25, 40, 60, 90, 150, 210 and 270 minutes. By the end of the experiments, coral fragments were transferred to containers with clean and circulating sea water to monitor polyp recovery at 340, 470 to 1200 minutes. The number of retracted polyps at different times was counted in five different (1 cm2) subareas of each coral fragment.
Field assessment of the allelopathic potential of cyanobacterial extracts
Field experiments were carried out in April 2012. To assess the allelopathic potential of cyanobacterial extracts, these were incorporated into gels of the polysaccharide PhytagelTM, a commercially available agar substitute gelling agent, and placed in direct contact with healthy corals following Thacker etal, (1998) and Rasher and Hay, (2010). The gel was prepared by mixing the PhytagelTM powder with hot water following the manufacturer's instructions and vigorously mixed. Once the gel had cooled, test extracts resuspended in acetone were blended into the PhytagelTM mixture and rapidly poured into acrylic molds covered with fine plastic mesh. The gels were then allowed to stiffen. The plastic mesh conveys greater strength to the gels allowing easier manipulation during field assays. Controls consisted of plain PhytagelTM and the same amount of acetone (< 100 mL) used to resuspend the extracts. Extracts tested BPVA, LP, and LF was incorporated into the gel at the natural concentration of 6 % (Prato, unpublished data). After hardening, the PhytagelTM covered plastic mesh was cut into strips.
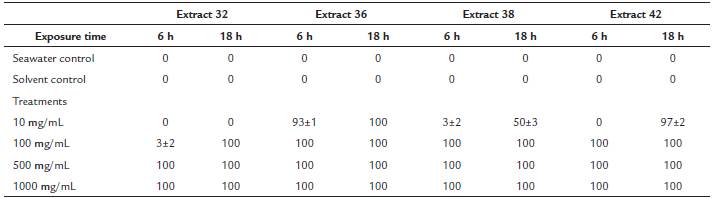
The field experiment was performed close the site Ministerio, north of Isla Grande, were dense stands of Porites porites were found. Treatment and control plastic mesh bands were softly placed over individual coral twigs and secured with plastic cable ties (Figure 3). There were five replicates for each cyanobacterial extract. Controls consisted of a band of PhytagelTM extract, to assess whether there was an effect by direct contact with a solid surface and a band of PhytagelTM with food-coloring (shading control) in order to emulate the coloration of the extract -which tends to be dark green- and discard bleaching responses attributable to darkening. In addition, at the time of removal of the experiment, five coral fragments (from the same colony) were removed to compare with experimental treatments. Treatments were removed after 24, 48 and 72 hours. Once the bands were removed, coral responses such as bleaching, tissue regression or necrosis were registered. Individual coral twigs were fragmented from the colony with pliers, wrapped individually in tin foil and introduced in mini ZiplockTM bags. The bags were immediately carried to the surface and frozen immediately in a bath of dry ice and acetone, which lowered the temperature to -70 °C, thus preventing degradation of photosynthetic pigments. Coral fragments were kept frozen until analysis.
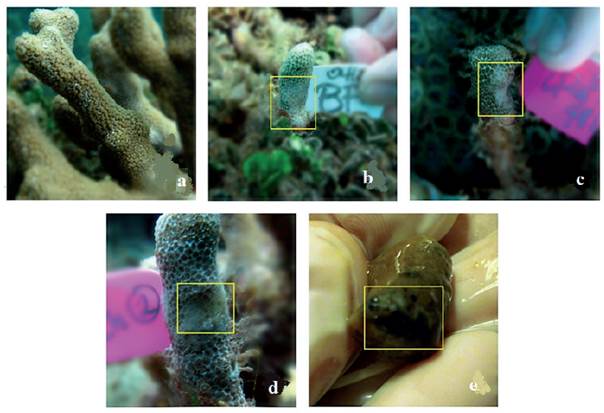
Figure 3 Effect of cyanobacterial extracts upon direct contact with the coral Porites porites. a. Untreated coral. b, c and d. Coral bleaching after 24, 48 and 72 hours respectively. e. Signs of necrosis (at 72 h) evident at the time of polyp removal for chemical analyses.
Gels were gradually removed in order to evaluate bleaching, a clear indicator of stress in corals (Titlyanov et al., 2007). Since the visual assessment of coral bleaching may be subjective, we used chromatographic techniques to assess the abundance and variation of the major photosynthetic pigments after contact with extracts. For this experiment several assumptions were made, (1) each of the coral fragments (treatment or control) was influenced by the same level of light radiation, (2) diversity and proportion of zooxanthellae clades in the coral (Knowlton and Rohwer, 2003) was the same in the fragment and the colony, (3) concentration of photosynthetic pigments between fragments at time zero and without any treatment was the same, (4) polyp removal and subsequent preparation of organic extracts were carried out in a uniform way for all treatments and controls.
Pigment analysis and quantification
The photosynthetic performance of coral-associated zooxanthellae has been evaluated with underwater PAM fluorometry (Rasher and Hay, 2010). However, given the cost of the device, we analyzed pigment content by HPLC following Kleppel et al., (1989) and Venn et al., (2006). For the extraction of photosynthetic pigments from coral fragments, these were thawed at room temperature and polyps covering an area of 1 cm2 were removed with pliers and tweezers. The polyp biomass was then extracted with acetone at 0 °C and sonication for two minutes. All these procedures were carried out in a dark chamber in order to prevent pigment degradation by exposure to light. Extracts were filtered through Pasteur pipettes with cotton plugs, concentrated under rotary evaporation and stored under a nitrogen atmosphere in the freezer. Prior to injection into the HPLC, crude extracts were resuspended in 90 % aqueous acetone and a 2:1 (v/v) ammonium acetate ion pair solution was added in order to increase hydrophobicity and retention time of the molecules of interest (Horvarth et al., 1977). Each extract was filtered using PVDF membranes 0.45 μm in diameter. 100 μL of each extract at a concentration of 1.33 mg mL-1 was injected (2 mg in 1 mL acetone extract plus 0.5 mL solution of the ion pair solution) into a RP-18 PhenomenexTM column, which was maintained at room temperature and eluted with a tertiary mobile phase gradient (0.5 M ammonium acetate in methanol/water 85:15, acetonitrile/ethyl acetate 90:10 and 100 % acetonitrile), at a flow rate of 1 mL min-1 for 50 minutes, with linear changes, following Louda et al., (2000). Spectral reading was carried out at 420 nm (Levy et al., 2003). Preliminary identification of pigments was based on UV absorption spectra and retention times of chromatogram compared to the data reported in the literature (Louda et al., 2000; Venn et al., 2006), since it was not possible to acquire adequate standards of dinoflagellate pigments. Therefore, only xanthophylls and chlorophylls were considered in the analysis, as well as pheophytin, a chlorophyll degradation product. Pigment quantification was carried out based on the areas under the curve of each chromatogram according to integration made by the chromatograph.
RESULTS
The taxonomical identity and toxicity assessment against Artemia salina nauplii of the cyanobacterial extracts evaluated in this study are included in Table 1. Recent studies have shown that the genetic, morphological and chemical diversity of tropical marine cyanobacteria has been largely underestimated (Engene et al., 2011; Engene et al., 2012; Engene et al., 2013a; 2013b; Engene et al., 2018). Accurate species identification involves molecular and chemotaxonomical analyses which can be quite complex. We want to point out that our species identification was performed before the thorough taxonomical revisions of tropical benthic cyanobacteria and was based just on the macroscopical features and microscopical analyses of the most common cyanobacterial mats. Extracts were selected based on their local abundance and available amount of extract in order to run the assays. We also wanted to compare the allelopathic effects of extracts with greater toxicity (extracts 32 and 38) in comparison to others that did not exhibit significant toxicity against A. salina (> 1000 (g mL-1) (extracts 36 and 42).
Table 1 Description, collection details, taxonomical identity and toxicity assessment against Artemia salina nauplii of the cyanobacterial consortia evaluated in this study. Species identification was performed before the thorough taxonomical revisions of tropical benthic cyanobacteria and were based on macroscopical features and microscopical analyses of the most common cyanobacteria in the consortia.

Lethality assay against fertilized coral eggs (embryos) of the scleractinian coral Orbicella annularis (Ellis and Solander, 1786)
All extracts evaluated were found to be toxic at 500 and 1000 mg mL-1 , causing a 100 % mortality of coral embryos in a six-hour period. Controls with sea water and solvent (ethanol) did not show mortality to the embryos during the evaluation period up to 18 hours. After 24 hours, controls showed partial mortality therefor observations for this time frame were not taken into account. The results of the lethality assays against embryos of O. annularis are presented in Table 2. The extract that exhibited the greatest toxicity was number 36 (a mixed assemblage of Lyngbya spp.). This extract caused up to 90 % mortality of embryos at the lowest concentration (10 mg mL-1) after only six hours of exposure.
Polyp retraction assay against fragments of the scleractinian coral Madracis mirabilis (=auretenra) (Locke, Weil and Coates, 2007).
All tested extracts led to the retraction of coral polyps with variable speed and intensity depending on the concentration tested. Solvent and sucrose controls elicited minimum polyp retraction and allowed rapid recovery. The results of this assay are included in Table 3. Extract number 38, caused polyp retraction faster and more severe within minutes of starting the essay at a concentration of11 mg mL-1. Moreover, coral fragments exposed to this extract did not fully recover even after 470 minutes. Only 16 hours after the initiation of the recovery phase did the corals extend all their polyps. Coral fragments exposed to cyanobacterial extracts had a maximum retraction percentage between 80 and 99 %, whereas in controls, polyp retraction did not surpass 25 %. After 20 hours (1200 min) in all cases, corals extended all their polyps displaying full recovery. Controls with the sucrose solution showed a maximum retraction of 23 % after 150 minutes. This value, however, was very low and most of the time polyps were fully extended, suggesting no effects from this organic substance. Solvent (ethanol) controls had a maximum retraction rate of 10 %. The strong retraction shown by coral polyps exposed to cyanobacterial extracts indicates coral stress and could suggest a potential allelopathic effect.
Table 3 Percentage of polyp retraction (mean ± standard error) of the coral Madracis mirabilis after indirect exposure to two organic extracts of benthic marine cyanobacteria (32 and 38) at two concentrations (2 mg L-1 and 11 mg L-1). Controls consisted of a 11 mg L-1 sucrose solution or ethanol. Once the exposure time finished (270 minutes), corals were transferred to containers with clean seawater and their recovery monitored (340, 470 to 1200 minutes). Values of "0" indicate that all polyps were extended and that the coral fragment was not affected by the extracts during exposure time or that they had fully recovered after 270 min.
Field assessment of the allelopathic potential of cyanobacterial extracts
Three to five coral fragments were analyzed for pigment variation upon contact with cyanobacterial extracts.
Unfortunately, quite a few of the treatments and replicates were lost due to excessive water motion. For instance, we lost all treatments of extract LP after 48 and 72 hours. Direct contact of corals with PhytagelTM bands with cyanobacterial extracts had negative effects on these, evidenced by polyp retraction, bleaching or surface erosion on twigs of Porites porites. This effect was apparently greater with exposure time. In some cases black bands, suggestive of colonization or growth of sulfate reducing bacteria were evident (Figure 3).
Pigment profile of zooxanthellae associated to Porites porites
After HPLC analyses, six pigments were selected for comparisons (1) dinoxanthin (a xanthophyll), (2) diadinoxanthin (a xanthophyll), (3) chlorophyll-c, (4) chlorophyll-b, (5) chlorophyll-a and (6) phaeophytin. Figure 4 shows the qualitative comparison of pigment variation over time. In all cases, as expected, extracts of untreated Porites tiwgs had the greatest concentrations of photosynthetic pigments (Figure 5, red columns). When comparing the effect of the different treatments over time, it was clear that in all cases (after 24, 48 and 72 hours of exposure) extract BPVA caused the greatest bleaching evidenced as the utmost pigment reduction in coral twigs. After 24 hours of exposure, coral twigs exposed to extracts LP and BF remained fairly constant in chlorophyll concentrations with respect to controls. After 48 hours of exposure, the trend was similar, where extract BPVA caused the greatest pigment reduction particularly in diadinoxanthin concentrations (Pigment 2). By the end of the observation period (72 hours), it became more evident there was a shift towards an increasing concentration of chlorophyll a and phaeophytin. Pigment reduction in coral twigs upon direct contact with cyanobacterial extracts was greater than in shading (PhytagelTM + food coloring) or PhytagelTM controls. These results suggest that coral bleaching is elicited by the extracts tested, particularly extract BPVA.
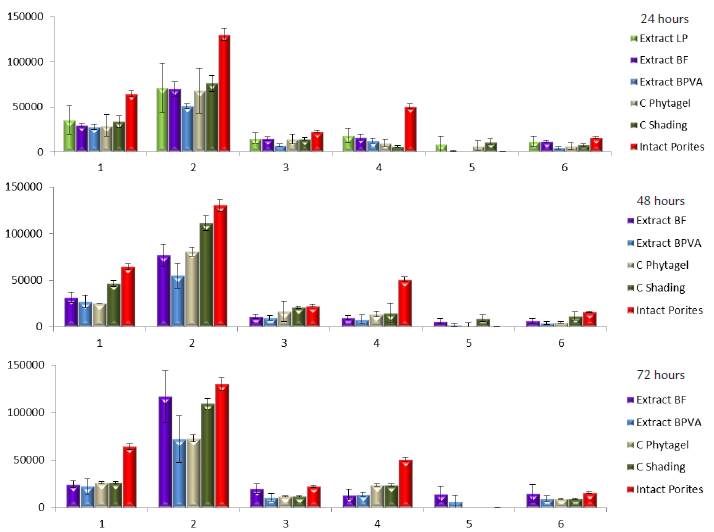
Figure 4 Qualitative variation of photosynthetic pigments (mAU) of zooxanthellae associated with the coral Porites porites over time after coral exposure to cyanobacterial extracts. Numbers refer to the following pigments (1) dinoxanthin (xanthophyll), (2) diadinoxanthin (xanthophyll), (3) chlorophyll-c, (4) chlorophyll-b, (5) chlorophyll-a and (6) phaeophytin.
DISCUSSION
Organic extracts ofmarine benthic cyanobacteria showed deleterious effects against coral larvae and also induced polyp retraction and bleaching in adult coral colonies. These results suggest that cyanobacteria may produce and release substances that prevent the settlement of coral larvae thus affecting their recruitment. In that case, there should be a mechanism by which these substances are actively exuded or passively released (cell breakdown upon death or mechanical friction against the substrate).
Many sessile organisms seem to rely on chemical defenses that protect them against epibiosis and fouling organisms. These defenses may be toxic to larvae of competing organisms or deter larval settlement on their surface (Fearon and Cameron, 1997). Physical barriers may also pose a problem for the settlement of coral larvae. Coral larvae might get entangled on mats of benthic cyanobacteria and die (Kuffner et al., 2004). Also, as we have witnessed in our study sites, algal meadows and cyanobacterial mats trap sediments which may also hamper larval settlement (Birrell et al., 2005). Similarly, lower portions of cyanobacteria mats may present anoxic conditions, beneficial for nitrogen fixation, but potentially harmful to coral larvae (Kuffner et al., 2004).
The polyp retraction assay in fragments of Madracis mirabilis, allowed us to evaluate coral responses to indirect contact with potential allelopathic substances (extracts) from benthic cyanobacteria. Petrichtcheva et al., (2002) and Richelle-Maurer et al., (2003) had previously tested extracts and compounds isolated from the Caribbean sponges Axinyssa ambrosia and Agelas conifera in the same coral as well. In the case of Agelas conifera, the minimum concentration in which the polyps showed retraction was 6 mg L-1 in a period of 270 minutes (Richelle-Maurer et al., 2003). In this work, we registered a very clear polyp retraction at much lower (3X) concentrations. Petrichtcheva et al., (2002) on the other hand, characterized axynissamine hydrochloride as the compound responsible for the significant defensive and allelopathic activity of the sponge Axinyssa ambrosia. That compound caused polyp retraction at a concentration of 0.007 mg mL-1 and had a LC50= 0.009 mg mL-1. In our case, cyanobacterial extracts elicited a clear polyp retraction at lower concentrations (2 mg L-1). Interestingly, all corals recovered and we did not detect coral mortality at the greatest concentration evaluated (11 mg L-1). In all these experiments concentrations tested were below the natural concentrations in the source organisms. Extracts were thus active at very low concentrations. Clearly, for a substance to have a potentially allelopathic effect requires its release into the environment. Although it is difficult to prove how that release actually occurs, in the case of marine benthic cyanobacteria, water movement and subsequent cell abrasion could favor the release of compounds that could affect other organisms as suggested by Titlyanov et al., (2007).
Visual determination of bleaching is obviously very subjective, thus quantification of individual pigments is a better, although difficult, approach to evaluate this response. In the past, only variations in the content of chlorophyll-b were indicative of coral bleaching, it is also important however, to take into account variations in xanthophyll and carotene contents (Venn et al., 2006). Although the assay evaluated the potentially allelopathic properties of cyanobacterial extracts, anoxic effects or the activity of sulfate reducing bacteria cannot be discarded (Baumgartner et al., 2006).
Shading controls and plain PhytagelTM controls, did not appear to elicit bleaching in twigs of Porites porites. Thus bleaching effects were attributed to substances present in the treatment gels. In the assay, a suggestive trend of phaeophytin increase was evident by the end of the observation period. Phaeophytin is a product of degradation of chlorophyll (Cartaxana et al., 2003), thus a mechanism by which cyanobacteria could compete with corals would be affecting the main photosynthetic pigments of coral associated zooxnathellae thus debilitating the coral host. The extract that showed the greatest reduction on the concentration of zooxanthellae photosynthetic pigments was BPVA, which incidentally exhibited the greatest toxicity against Artemia salina. This extract was obtained from a mixed consortium of Lyngbya spp. and Oscillatoria spp. The cyanobacterium once referred to as Lyngbya majuscula is recognized as a prolific source oftoxic and allelopathic compounds. However, recent genetic and chemotaxonomical studies have reassessed the taxonomy of the genus Lyngbya and allowed to reclassify this group in a complex of species within the genera Moorea and Okeania, which explains the great chemical variability in this group of cyanobacteria (Engene et al., 2011; Engene et al., 2012; Engene et al., 2013 a, b).
CONCLUSIONS
Our results show that some consortia of marine benthic cyanobacteria from the Colombian Caribbean have negative effects on corals and that allelopathic mechanisms may play an important role in competitive interactions between corals and cyanobacteria. Negative effects may be either as a result of abrasion by direct physical contact but also due to potentially allelopathic mechanisms affecting coral larvae and indirectly or directly affecting adult corals. These mechanisms along with those that confer protection against herbivory constitute effective adaptations enabling cyanobacteria to effectively colonize the substrate, forming large mats, dominating and surviving in reef environments.


