











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
La caracterización de los aspectos auto-ecológicos de las especies es de gran importancia para entender y modelar la dinámica de sus poblaciones (Arismendi etal., 2011; Martins-Camara etal., 2011). Por ejemplo, la relación longitud-peso (RLP) y el factor de condición (K) son dos aspectos que resumen el crecimiento, la capacidad de supervivencia y reproductiva de las especies, y que por tanto, podrían utilizarse en conjunto para aproximarse a su dinámica poblacional (Cifuentes etal., 2012). La RLP ha sido utilizada para describir los aspectos del ciclo de vida de los peces, los aspectos reproductivos, las condiciones de alimentación (Le Cren, 1951), y los cambios energéticos en el tiempo y el espacio (Freitas et al., 2014). Mientras que K ha servido como una medida del bienestar general de las poblaciones y como indicador nutricional de los peces (Baigún et al., 2009), generalmente asumiendo que los cambios están mediados por alteraciones fisiológicas o ecológicas (Sutton et al., 2000). Sin embargo, a pesar de su potencial aplicado, este tipo de aspectos han sido caracterizados sólo para especies de uso comercial que habitan en zonas templadas, siendo escasas las caracterizaciones de especies nativas en ríos tropicales (Froese, 2006).
Bryconops (Kner, 1858) es un género perteneciente a la familia Iguanodectidae, cuenta con 20 especies que se encuentran distribuidas en gran parte de Suramérica, en las cuencas de los ríos Orinoco, Amazonas, Tocatins-Araguaia, Paraná-Paraguay y San Francisco (Lima y Caires, 2011; Wingert y Malabarba 2011; Silva-Oliveira et al., 2015), incluyendo varias cuencas costeras en las Guayanas y Brasil (Froese, 2006). La mayoría de las especies son omnívoras y activas (Perillo-Nogueria et al., 1997), y se encuentran en ambientes lóticos, típicamente ácidos y transparentes (Chernoff y Machado-Allison, 2005). Bryconops giacopinii (Fernández-Yépes, 1950) es un pez de pequeño porte, con cuerpo cilíndrico que puede alcanzar los 18 cm de longitud estándar (Giarrizzo et al., 2015). Se caracteriza por una franja longitudinal plateada desde el opérculo hasta la base de la aleta caudal, el lóbulo superior de la aleta caudal es negro con una mancha redonda de color amarillo - naranja en la base (Gálvis et al., 2007).
En Colombia, a pesar de que B. giacopinii se encuentra ampliamente distribuida en la cuenca del río Orinoco y Amazonas (DoNascimiento et al., 2017) y de que es considerada como especie de importancia en el comercio de peces ornamentales (Ortega-Lara, 2016) a la fecha, no se han realizado estudios auto-ecológicos, los cuales brinden un aporte significativo al conocimiento auto-ecológico de la especie o a la dinámica de sus poblaciones. Considerando el gran número de especies de peces que habitan las cuencas de los ríos Amazonas y Orinoco y las presiones crecientes sobre las mismas (Lasso et al., 2010; 2011), caracterizar los aspectos auto-ecológicos y su variación natural en los sistemas que habitan se vuelve de gran importancia para su conservación.
Teniendo en cuenta lo anterior, el presente estudio buscó caracterizar los siguientes aspectos auto-ecológicos de B. giacopinii: 1) la relación longitud-peso, 2) el factor de condición y 3) la dieta, en sistemas con baja intervención antropogénica como los de la Serranía de La Lindosa.
MATERIALES Y MÉTODOS
Área de estudio
La Serranía de La Lindosa, se encuentra cerca de San José del Guaviare, Colombia, a lo largo de la parte norte de la Amazonía colombiana, en una zona de transición entre las eco regiones del Orinoco y el Amazonas (Cárdenas-López et al., 2008; Figura. 1). El clima es considerado lluvioso tropical, con un promedio anual de precipitación de 2800 mm y un patrón unimodal con temporada lluviosa desde marzo hasta octubre y una temporada seca de noviembre a febrero. Se encuentra conformada en su mayoría por rocas sedimentarias de la formación Araracuara, Sienita Nefelina de San José del Guaviare y rocas sedimentarias del Terciario de la Amazonia, los suelos son areniscas provenientes principalmente de la formación Paleozoica de Araracuara dominados por cuarzos y químicamente pobres (Duivenvoorden y Cleef, 1994; Ortega-Lara, 2016).
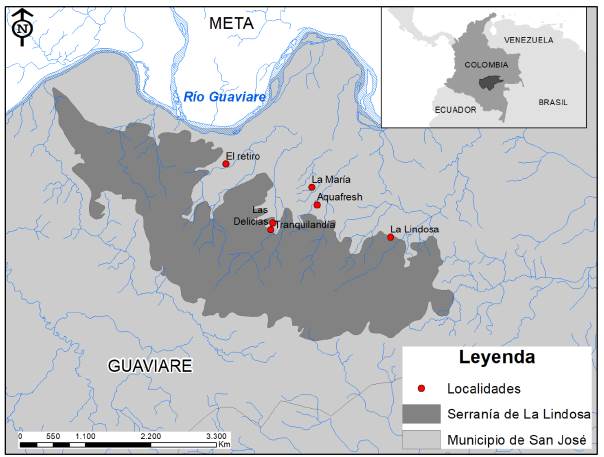
Figura 1 Localización geográfica de los seis tramos de muestreo dentro de la cuenca de la Serranía la Lindosa, municipio de San José del Guaviare (Colombia). Los tramos se indican dentro del mapa en forma de puntos rojos.
Los cuerpos de agua, en su mayoría pequeños cauces que drenan la Serrania de La Lindosa, tienen lechos compuestos por grandes losas relictas del escudo guyanés que conforman secuencias marcadas de rápidos y pozos, cuyas transiciones están dadas frecuentemente por cascadas de altura variable (en ocasiones pueden alcanzar más de 30 m, González-Trujillo obs. per.). Debido a las características de su lecho y a la baja concentración de nutrientes en los suelos de la cuenca, estos ecosistemas pueden ser considerados como ultra-oligotróficos. El agua del cauce tiene una baja concentración y disponibilidad de nutrientes, valores de conductividad cercanos a los 4 uS/cm3, pHs ácidos (entre 5 y 6 unidades), y una concentración relativamente alta de carbono orgánico disuelto (González-Trujillo, s.f.). La vegetación de ribera está compuesta principalmente por especies de bosque de galería, aunque puede cambiar abruptamente al alejarse del cauce principal, siendo las especies de porte bajo como arbustales y rosetales las que aumentan su abundancia (Cárdenas-López et al., 2008).
Muestreo de la población
En cada una de las seis localidades fue escogido un transecto de 50 m de longitud (Figura. 1). La comunidad íctica fue muestreada en cada transecto por medio de tres pases con una malla de angeo plástico (10 x 2 m de área, 2 mm de ojo de malla) durante el inicio (abril de 2013 y 2014) y el final del período lluvioso (octubre 2012 y 2013), para un total de cuatro muestreos en cada uno de los seis sitios (dos eventos de muestreo por temporada). Los individuos fueron colectados durante las salidas de campo del curso de Taxonomía animal del departamento de Biología de la Universidad Nacional de Colombia en Bogotá, por lo tanto, se siguieron todas las directrices de colecta, contándose con el permiso marco de colecta de la Universidad Nacional de Colombia. Los peces fueron fijados en formol al 4 % inmediatamente después de su captura y preservados en etanol al 70 %, para después ser depositados en la colección ictiológica del Instituto de Ciencias Naturales (ICN -Universidad Nacional de Colombia). Se registró el tamaño corporal (longitud estándar - LE; 1 mm), peso (peso total - PT; 0,1 g) y se procedió a retirar el tracto digestivo por medio de una incisión abdominal.
Caracterización del crecimiento y de la condición de los peces
La relación longitud-peso (RLP), se estimó por la expresión Pt = α xLe b , donde Pt es el peso total, Le la longitud estándar, a el intercepto de la curva y b el coeficiente de alometría de la relación longitud-peso; su valor se determina transformando la ecuación en un modelo lineal (Sparre y Venema, 1997). Se realizaron una serie de regresiones lineales con el total de individuos (n= 333) y entre cada localidad de muestreo.
Se evaluó de forma independiente la variabilidad espacial y temporal de la RLP debido a que no se disponía de un tamaño de muestra adecuado para hacer regresiones de las poblaciones de cada una de las localidades en las dos épocas distintas (interacción espacio - temporal). Los valores de longitud y talla fueron transformados con la función del logaritmo en base de 10 para cumplir con los supuestos de un modelo lineal. Un análisis de covarianzas (ANCOVA) fue utilizado para evaluar si existían diferencias de la RLP (resumida por la pendiente de la regresión) entre poblaciones al controlar la localidad y la temporada (seca y lluviosa) en que fueron muestreadas. Se escogió un p-valor menor a 0,05 como umbral. Adicionalmente, la pendiente de las regresiones (b) fue utilizada como una aproximación al tipo de crecimiento de cada una de las poblaciones. De acuerdo con Cifuentes et al., (2012), si una población de peces mantiene la relación peso-longitud (b= 3), entonces presenta un crecimiento isométrico. Mientras que si b>3, el pez ha incrementado su peso en mayor proporción que su longitud y por lo tanto presenta un crecimiento alométrico positivo. En cambio, si b<3, la longitud del pez incrementó en mayor proporción que el peso y por lo tanto se asume que tiene un crecimiento alométrico negativo.
El factor de condición (K) individual fue calculado con el índice de Fulton (Ricker, 1975) siguiendo la ecuación:
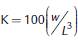 . Donde w es el peso (g) y L es la longitud (cm). Se asume que los peces más pesados de una determinada talla se encuentran en mejor condición que aquellos de la misma talla, pero de menor peso (Badillo et al., 2010).
. Donde w es el peso (g) y L es la longitud (cm). Se asume que los peces más pesados de una determinada talla se encuentran en mejor condición que aquellos de la misma talla, pero de menor peso (Badillo et al., 2010).
Descripción de la dieta
Los contenidos estomacales fueron separados bajo un estereoscopio y analizados siguiendo un método volumétrico (Winemiller, 1989). El volumen de cada ítem alimenticio fue medido por el desplazamiento de agua dentro de un cilindro con una resolución de 0,01 ml. Los ítems alimenticios con un volumen inferior a 0,01 ml fueron medidos sobre papel milimetrado y su volumen fue estimado multiplicando el área ocupada en milímetros por su altura. Cada ítem alimenticio fue identificado al menor nivel taxonómico posible, el cual dependió del grado de digestión en el que se encontraba. Para la identificación se usaron las claves taxonómicas de Domínguez y Fernández, (2009) para los invertebrados acuáticos y Johnson y Tripplehorn, (2005) para los invertebrados terrestres. Para facilitar el análisis estadístico, los ítems alimenticios fueron agrupados en cuatro categorías funcionales: artrópodos terrestres, insectos acuáticos, material vegetal y peces. Para caracterizar la dieta, se calculó la frecuencia de ocurrencia (% FO) de cada categoría alimenticia, y su proporción volumétrica (% V) con respecto al total de la dieta (Hyslop, 1980; Winemiller, 1989).
Se utilizó un análisis de escalamiento multidimensional no métrico ("nMDS") para evaluar si la dieta de las poblaciones tenía alguna estructura o diferenciación de acuerdo con la temporada o localidad muestreados. Aquellos ítems que fueron encontrados en más de cinco estómagos fueron incluidos en el análisis. Esto con el fin de evitar la influencia de ítems de ingesta ocasional en el análisis estadístico. El análisis se llevó a cabo utilizando la distancia de Morisita-Horn (Morisita, 1961; Horn, 1966) ya que ésta les da menos peso a aquellos ítems raros o de baja ocurrencia que se observaron durante el análisis estomacal. Una prueba de Kruskal-Wallis se usó para determinar si el volumen de las categorías era diferente al compararla entre las localidades muestreadas.
Las regresiones, la ordenación por medio del nMDS y las salidas gráficas fueron realizadas en R v.3.4.0 (CRAN, 2017) utilizando los paquetes vegan (Oksanen et al., 2015) y ggplot2 (Wickham, 2009).
RESULTADOS
Aspectos relacionados con el crecimiento y la condición de los peces
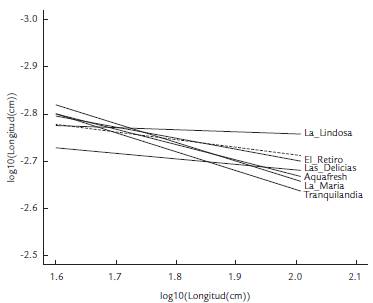
En términos de tipos de crecimiento, tanto a nivel de especie como poblacional, se observó un crecimiento alométrico positivo (b>3, Figura 2). Este tipo de crecimiento se da cuando el peso de los peces incrementa en mayor proporción que su longitud. A nivel poblacional no se observó una diferencia significativa entre poblaciones al controlar la temporada de muestreo (ANCOVA, F(1,329) = 0,256, p = 0,62), pero sí se encontraron diferencias al controlar por la localidad (ANCOVA, F(6,319) = 2,696, p = 0,014). Las poblaciones de La Lindosa y Las Delicias (b = 3,05 y b = 3,11, respectivamente) tuvieron los coeficientes de crecimiento más bajos, asemejándose más a los valores de coeficientes que indican un crecimiento isométrico (b = 3). Este tipo de crecimiento indicaría que en estas poblaciones el peso y la longitud de los peces incrementaron de manera proporcional.
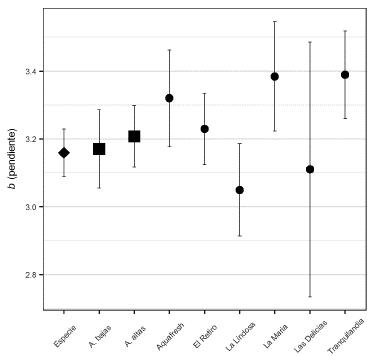
Figura 2 Pendientes (b) e intervalos de confianza (95 %) de las regresiones peso-longitud (RLP). El parámetro para la especie se representa en forma de rombo, para las poblaciones de cada temporada (aguas altas y bajas) en forma de cuadrado, y para las poblaciones de cada río en forma de círculos.
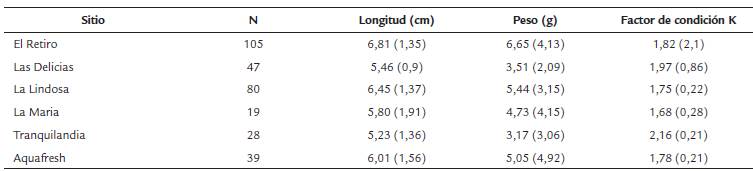
Tabla 1 Localidades de muestreo con el respectivo número de individuos capturados con promedios y desviación estándar para la longitud, peso y factor de condición.
En la Figura 3 se representa la tendencia del factor de condición (K) a lo largo de las diferentes tallas para las poblaciones de cada localidad y para la especie (n = 333). El factor de condición fue similar a lo largo de todas las tallas en las poblaciones de La Lindosa y las Delicias. En las otras localidades, el factor de condición siguió una tendencia similar a la del modelo general para la especie: una disminución con el incremento de la talla. En las poblaciones de Tranquilandia, Aquafresh y La Maria, la tendencia a la disminución fue mucho más marcada que para las otras poblaciones.
Aspectos relacionados con la dieta
Se analizaron los contenidos estomacales de 206 individuos, encontrando 17 tipos de ítems alimenticios: diez órdenes de artrópodos terrestres, tres órdenes de insectos acuáticos, restos de hojas, flores, frutos y semillas de al menos tres familias de plantas terrestres, y algunos restos de escamas y otros peces, en su mayoría de la familia Characidae. Así mismo, se observó la presencia de restos de peces dentro del contenido de 28 estómagos evaluados: en los individuos de tallas inferiores a los 5 cm se encontraron escamas, mientras que en los individuos con tallas superiores a los 6 cm se encontraron restos de tejido muscular y escamas. De acuerdo a la ordenación dada por el nMDS, la ocurrencia de los ítems alimenticios en los estómagos no siguió ningún patrón temporal o espacial (Figura 4).
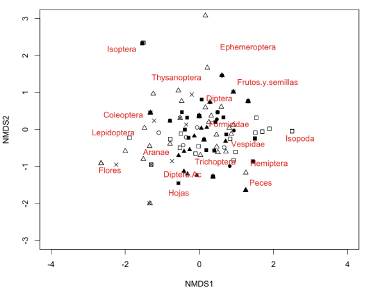
Figura 4 Ordenación de los tramos de muestreo de acuerdo con la dieta de las poblaciones de B. giacopinii por medio de un análisis de escalamiento multidimensional no métrico (nMDS). Distancia utilizada: Horn-Morisita. Estrés total = 0,084. Ajuste no métrico de la representación (R2) = 0,993.
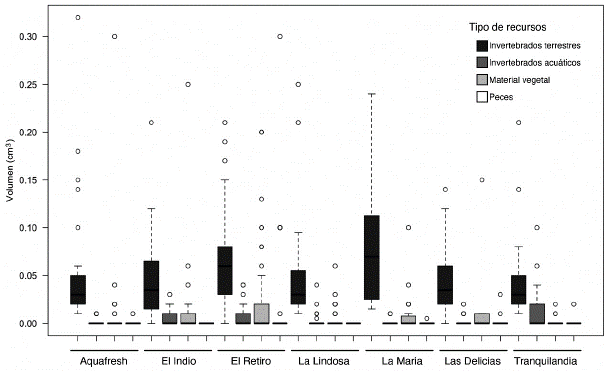
Al agrupar los diferentes ítems en cuatro categorías (artrópodos terrestres, insectos acuáticos, material vegetal, peces) se observó que las poblaciones de B. giacopinii consumen mayoritariamente artrópodos terrestres que caen ocasionalmente al agua (Figura 5). El volumen de este grupo de ítems no varío significativamente entre las diferentes localidades (Kruskal-Wallis test - Chi-cuadrado = 11,84, gl = 6, p-valor = 0,0656). En las localidades de El Indio, El Retiro y Tranquilandia se observó un volumen significativamente mayor de insectos acuáticos (Kruskal-Wallis test - Chi-cuadrado = 16,64, gl = 6, p-valor = 0,0107), y en menor grado, de material vegetal (Kruskal-Wallis test - Chi-cuadrado = 14,613, gl = 6, p-valor = 0,0235). Sin embargo, considerando las diferencias en términos de magnitud con respecto a los artrópodos terrestres, estos ítems podrían considerarse como minoritarios dentro de la dieta de B. giacopinii.
DISCUSIÓN
El presente estudio es el primero para la especie B. giacopinii en el país, por tanto, amplia el conocimiento biológico de la diversidad íctica de Colombia y de áreas poco estudiadas como la Serranía de La Lindosa. Los análisis aplicados en esta investigación han sido utilizados en otras partes del mundo como información biológica básica para la elaboración de planes de manejo y conservación de distintas especies de peces. En Colombia, son pocos los trabajos que han sido desarrollados con estos métodos, la mayoría están limitados a especies de interés comercial como la Cachama (Piaractus brachypomus), el Bocachico (Prochilodus magdaleane) y la mojarra (Oreochromis sp. ). De esta manera, el procedimiento metodológico utilizado en este estudio podría ser considerado como base para el desarrollo de futuras caracterizaciones de aspectos auto ecológicos del gran número de especies de peces que habitan los cuerpos de agua del país.
Aspectos relacionados con el crecimiento y la condición de los peces
La relación longitud-peso fue incluida como una aproximación a la forma de crecimiento de las poblaciones de B. giacopinii en diferentes localidades (ríos) y temporadas de lluvias. La baja variabilidad de los coeficientes de alometría (estimados a partir de las pendientes - "b" -de las regresiones lineales) entre épocas y localidades con respecto al coeficiente obtenido para la especie podrían estar relacionados con la baja intervención de los diferentes puntos de muestreo. En condiciones óptimas y ante la ausencia de factores que impacten directamente sobre la dieta de los peces (p.ej. pérdida del alimento de origen alóctono debido a la pérdida de la vegetación riparia), los peces tienden a presentar un crecimiento alométrico positivo (Abowei, 2010). Aunque estos valores difieren de los reportados para poblaciones brasileras de B. giacopinii por Giarrizzo etal., (2015, b= 2,85) y FishBase, (2018, b= 2,95), cabe resaltar que estos coeficientes son similares al de otras especies dentro del mismo género en la Amazonia brasilera (Freitas et al., 2014; Oliveira et al., 2014; Giarrizzo et al., 2015; Rosa et al., 2016; Nobile et al., 2017).
La variabilidad observada entre localidades podría estar relacionada con diferencias puntuales de los ríos de la Serranía de La Lindosa en términos de temperatura (Beacham, 1990), velocidad de la corriente (Imre et al., 2002) o de composición de microhábitats (Beacham, 1990). Sidlauskas et al., (2006), por ejemplo, encontraron que la temperatura y la velocidad del agua fueron los factores ambientales más correlacionados con la variación morfológica intraespecífica en la forma del cuerpo y la longitud del pedúnculo caudal de Bryconops melanurus. Ante la ausencia de diferencias significativas en la dieta (véase siguiente sección), los coeficientes menores observados en la Lindosa y las Delicias podrían estar relacionados con algún factor ambiental como se ha reportado en otras partes del mundo. Sin embargo, una caracterización sistemática del hábitat en cada uno de los ríos es necesaria para determinar cuáles factores en concreto pueden estar afectando la forma de crecimiento en las poblaciones de la Serranía La Lindosa.
Según Le Cren, (1951), el factor de condición (K) es un parámetro cuantitativo que evalúa el bienestar de los peces, que determina el éxito presente y futuro de las poblaciones por su influencia en el crecimiento, reproducción y supervivencia. El factor de condición de los peces suele depender de diversas variables: los cambios estacionales (Trudel et al., 2005), cambios en la disponibilidad del alimento (Pothoven et al., 2001; DeBruyne et al., 2008) y cambios en la forma del cuerpo a lo largo de la historia de vida (Blackwell et al., 2000). Contrario a lo esperado para las poblaciones de B. giacopinni, no se encontraron diferencias entre las dos épocas muestreadas asociadas con variaciones en la dieta (los insectos terrestres son el recurso energético más importante durante el todo el año). Colectivamente, estos resultados indican que las poblaciones de la Serranía de la Lindosa se encuentran en óptimo estado de bienestar a lo largo del año, por tanto, su dinámica podría no verse afectada en un futuro cercano.
Cabe resaltar que el factor de condición de los individuos de menores tallas tendió a ser mayor en la mayoría de los ríos estudiados, sugiriendo que las planicies de inundación podrían representar un hábitat de guardería para las larvas y juveniles, donde la disponibilidad de alimento y refugio es amplia durante la época de lluvias (Cifuentes et al., 2012). Esto correspondería a su vez con el alto contenido de cuerpos grasos observados durante la disección de los peces, que sugiere una alta disponibilidad de energía para cada individuo (Herbinger y Friars, 1991). La única excepción ocurrió en los ríos La Lindosa y las Delicias, cuyas poblaciones también tuvieron las menores pendientes en las RLP. En estos ríos, el factor de condición se mantuvo estable a lo largo de las tallas, lo que sugiere un umbral de crecimiento en las poblaciones que allí habitan. Como se mencionó anteriormente, ante la ausencia de una diferencia en las dietas de los individuos, este umbral podría estar determinado por la acción de uno o más factores ambientales restringiendo el crecimiento.
Aspectos relacionados con la dieta
La entrada de materia orgánica desde la vegetación de ribera conecta los ecosistemas acuáticos y terrestres de tal manera que los insectos terrestres, las frutas y semillas, son generalmente la fuente principal de alimento para los peces (Vannote et al., 1980; Lorion y Kennedy, 2009). Esto concuerda con la importancia que presentó el volumen de esta categoría en la dieta de B.giacopinii. El alimento de origen alóctono es por tanto la fuente de energía más importante para el mantenimiento de las poblaciones, como se ha registrado previamente para esta especie (Sabino y Zuanon, 1998) para otras especies del género como B. caudamaculatus y B. alburnoides (da Silva et al., 2008). De acuerdo con Winemiller, (1990) el alimento de origen alóctono presenta una mayor disponibilidad durante los periodos de aguas altas debido a la inundaciones y la acción de los vientos y lluvias. Por tanto, se esperaba que B. giacopinii consumiera mayoritariamente alimento de origen alóctono durante la época lluviosa; sin embargo, no se observó una variación estacional en la dieta en las poblaciones de la especie. Los insectos terrestres fueron abundantes en los estómagos de los individuos durante las dos épocas de estudio.
La dominancia del material alóctono junto a la posición de la boca de B. giacopinii sugiere que esta especie tiene un hábito alimenticio generalista y que obtiene el recurso mayoritariamente al capturar los ítems que caen al agua, saltando fuera de esta o consumiendo presas a la deriva en la columna de agua (Costa-Pereira y Severo-Neto, 2012). Considerando su abundancia en estos ríos y su relación con el material alóctono, B. giacopinii puede ser un modelo de estudio adecuado para entender cómo funciona la interfaz terrestre-acuática en ecosistemas fluviales del escudo guyanés. Futuros estudios podrían enfocarse a determinar los factores que regulan el crecimiento de los individuos y la dinámica poblacional, y cómo esto se relaciona con la transferencia de energía que ingresa del componente alóctono al ecosistema fluvial. Estos estudios podrían contribuir a entender el rol de los bosques de ribera en el funcionamiento de estos ecosistemas fluviales tan amenazados por la creciente deforestación.
CONCLUSIONES
Este es el primer estudio en evaluar estos aspectos auto-ecológicos de B. giacopinii en Colombia. Por medio de técnicas no invasivas como la relación longitud-peso y índice de condición se describieron los parámetros de crecimiento y bienestar de seis poblaciones de B. giacopinii en la Serranía de La Lindosa. Las poblaciones presentes en esta parte de Colombia presentan un crecimiento alométrico positivo, con coeficientes más altos que los presentados por la misma especie y especies congénero en la Amazonía brasilera. El factor de condición de B. giacopinii es óptimo, sin embargo, estos resultados deben manejarse con cuidado ya que son los primeros que se obtienen de esta manera para esta especie en esta región de Colombia con baja intervención en los ecosistemas acuáticos. B. giacopinii es una especie generalista ya que se alimenta de un gran espectro de ítems alimenticio alóctonos y autóctonos. Por otra parte, el alto consumo de artrópodos terrestres por parte de B. giacopinii durante las dos épocas de muestreo evidencia la estrecha relación que hay entre esta especie y el bosque de ribera. Por lo tanto, la conservación y mantenimiento de estos corredores vegetales son necesarios para el bienestar de las poblaciones de esta especie. Estudios futuros son necesarios para poder establecer comparaciones entre poblaciones que están asentadas en otras regiones del país y habitan ríos con diferentes grados de intervención.


