











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCCIÓN
Los dinoflagelados son el segundo grupo más abundante del fitoplancton, responsable de la producción de energía en las redes tróficas del océano (Lalli y Parson, 1997). Su adaptación a gran variedad de ambientes se ve reflejada en la gran diversidad de formas, tipos de nutrición y un enorme registro fósil que data de millones de años (Rizzo, 1987). Poseen dos flagelos móviles y en su mayoría son unicelulares, aunque algunos forman colonias (Gómez y López-García, 2011) . Existen especies autotróficas, otras heterotróficas y un grupo muy reducido con condiciones mixotróficas (Lalli y Parson, 1997). Se estima que aproximadamente en los océanos 2294 son especies de vida libre marina (Gómez, 2012) , en el gran Caribe 404 (Wood, 1968) y 171 en el Caribe colombiano (Lozano-Duque et al., 2011).
Los dinoflagelados según su morfología se dividen en dos grupos; los tecados (cubiertos) por la presencia de placas celulósicas en el anfiesma; y los atecados (desnudos) por el poco desarrollo o la ausencia de estas placas (Balech, 1988; Larsen, 2002). Los atecados presentan una pared frágil y placas finas, lo que generan dificultades en la observación por microscopía óptica (Maciel-Baltazar y Hernández-Becerril, 2013), suelen pertenecer a los órdenes Oxyrrhiales, Actiniscales, Gymnodiniales y Noctilucales y los caracteres morfológicos que los distinguen son: tamaño, forma general, posición y morfología del cingulum (desplazamiento), el sulcus, la presencia o ausencia de cloroplastos y pirenoides, forma, posición del núcleo y posibles estructuras superficiales (Larsen y Sournia, 1991). Caracteres de gran relevancia como la hendidura apical, los apéndices y las manchas oculares, solo se pueden observar en muestras frescas sin fijación (Larsen, 2002).
Con el propósito de mitigar las presiones sobre los ecosistemas estratégicos regionales y en particular los ecosistemas de coral, Colombia declaró en el año 2013 Área Marina Protegida-AMP al Parque Nacional Natural Corales de Profundidad-PNNCPR (Hincapié, 2015), constituyendo cerca del 40 % de la biodiversidad del borde de la plataforma continental del Caribe colombiano (Alonso et al., 2015). Los corales de profundidad son considerados ecosistemas estratégicos de biodiversidad en los océanos, se cree que albergan una mayor diversidad de especies que los arrecifes de corales duros tropicales (Echeverry, 2013). Esto representa gran relevancia entre los ecosistemas marinos al brindar un importante valor económico, ecológico y social, mediante la oferta de bienes y servicios ecosistémicos (Maldonado y Cuervo, 2016).
Colombia como país megadiverso debe continuar adelantando acciones para extender el conocimiento de su diversidad biológica, en especial de sus AMP soportado en la comunidad científica nacional e internacional, con el fin de generar información como insumo para acciones de manejo y la toma de decisiones a nivel ejecutivo basadas en datos científicos. Por tal razón, en función a su relevancia ecológica y los pocos reportes de dinoflagelados atecados en el Caribe colombiano, derivado en la dificultad de preservar y observar los especímenes, el presente trabajo tiene el objetivo de describir la presencia del dinoflagelado atecado P. spinifera, algunos aspectos morfológicos y las condiciones ambientales de las aguas del PNNCPR en donde se hallaron estos ejemplares.
MATERIALES Y MÉTODOS
Área de estudio
El PNNCPR se encuentra ubicado en el Caribe colombiano a una distancia aproximada de 12 km del PNN Corales del Rosario y San Bernardo y a 32 km de la Península de Barú (Fig. 1) (Hincapié, 2015). Cuenta con una extensión aproximada de 1421 km2, con profundidades que oscilan entre 24 y 800 m y temperatura ambiental entre 27 a 30 °C. Constituido por tres hábitats importantes: fondos blandos profundos, corales profundos y corales mesofóticos (Alonso et al., 2015; Parques Corales, 2016).
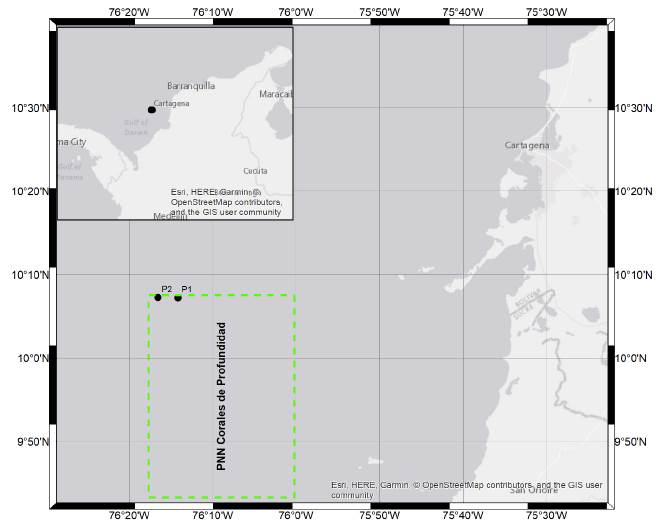
La cuenca del Caribe colombiano esta modulada por el desplazamiento de la zona de convergencia intertropical (ZCIT), la cual consiste en una cinta longitudinal formada por masas de aire que provienen del hemisferio norte y sur (Sarmiento-Devia et al., 2013). El muestreo fue realizado en el mes de mayo, cuando la ZCIT se encuentra localizada hacia el sur, correspondiente a la época seca (Lonin et al., 2010), manifestándose con vientos moderados y en ocasiones vientos fuertes provenientes del norte (N) y noreste (NE), (Andrade, 1993).
Fase de campo
En una campaña oceanográfica desarrollada en mayo del 2016, fueron tomadas muestras de agua en roseta oceanográfica Carousel SBE 32 equipada con botellas tipo Niskin de 8 L y perfi lador CTD SeaBird Electronics 19 Plus, en el buque oceanográfico A.R.C "Providencia"; las muestras fueron colectadas en dos puntos del PNNCPR; el punto uno (P1) muestreado a un metro de profundidad y el punto dos (P2) a 40 m. Asimismo fueron tomadas mediciones in situ de salinidad y temperatura con conductímetro WTW pH/ Cond 3320 y el perfilador antes descrito. Para las muestras fitoplanctónicas fueron colectados 500 mL de agua marina, los cuales fueron depositados en botellas plásticas ámbar y preservadas con formaldehido al 4 % neutralizado con tetraborato de sodio (Throndsen, 1978; Boltvskoy, 1995; Edler y Elbráchter, 2010).
Fase de laboratorio
El montaje y conteo de las células fue realizado mediante el método de cámaras de sedimentación Utermöhl, (1958); las muestras fueron sedimentadas por un periodo de tiempo de 48 horas, utilizando un cilindro de 100 mL debido a la baja concentración de organismos fitoplanctónicos previamente observados con montajes de volúmenes inferiores. La observación de las muestras fue realizada en el laboratorio de biología de DIMAR-CIOH Caribe mediante el microscopio invertido Olympus IX73 a magnificaciones de 200X y 600X. Los registros y mediciones se realizaron con la cámara Olympus XC50 con el Software Olympus Cellsens Standard 1.13.
Identificación
La identificación taxonómica de P. spinifera fue realizada tomando como referencia los trabajos de Schiller, (1933), Balech, (1971), Taylor, (1976), Balech, (1988), Steidinger y Tangen, (1996), Ojeda, (1998), Gómez, (2013b), Maciel-Baltazar y Hernández-Becerril, (2013), Gárate-Lizárraga, (2014) y Escobar-Morales y Hernández-Becerril, (2015).
RESULTADOS
Pronoctiluca Spinifera (Lohman), (Schiller, 1933) (Figs. 2A-E).
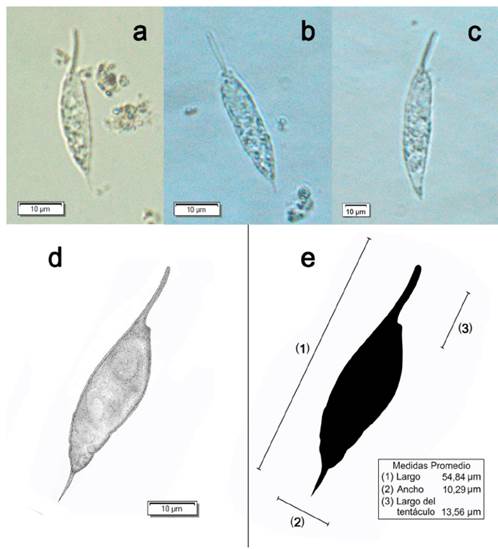
Figura 2 Resultados de las observaciones de Pronoctiluca spinifera en microscopio óptico con magnificación 60x. a-b) Ejemplares colectados a un metro de profundidad (superficial), c) ejemplar 3, corresponde a 40 m de profundidad, las dimensiones de los ejemplares se muestran en la Tabla 1. d) Descripción morfológica de Pronoctiluca spinifera, a partir de las figuras a-c, e) Medidas promedio de Pronoctiluca spinifera.
Sinonimia: Protodinifer tentaculatum Kofoid y Swezy 1921, p. 112, pl. 7, fig. 74, figura en el texto R, 2 Cystodinium sp:Balech, 1971, p. 168, lám. 38, fig. 747.
Referencias: Schiller (1933, p. 270, fig. 259 a-d) Taylor (1976, p. 188, pl. 37. fig. 429); Balech (1988, p. 200, lám. 87, fig. 5) Steidinger and Tangen (1996, p. 465, lám. 23), Ojeda (1998, p. 160, lám. 12), Gómez (2013b, p. 73, fig. 2 a-j), Maciel-Baltazar y Hernández-Becerril (2013, p. 253, fig. 3G), Gárate-Lizárraga (2014, p. 580, fig. 3R) Escobar-Morales and Hernández-Becerril (2015, p. 18, fig. 32).
Se logró realizar la descripción morfológica de P. spinifera (Fig. 2d) con registros fotográficos que se obtuvieron en las observaciones de organismos fitoplanctónicos (Fig. 2 a-c). Se observaron células solitarias con longitud aproximadamente cuatro o cinco veces el diámetro, contorno fusiforme; presencia de dos extensiones, una proyección anterior en forma de tentáculo y otra puntiaguda mucho más corta en el extremo opuesto. Núcleo circular situado cerca del extremo anterior y pequeños residuos de sustancias de reserva en su interior, evidenciando características heterotróficas.
Las células de P. spinifera fueron encontradas en dos puntos, el primero a un metro de profundidad (10°9' N, y 76°17' W) y en el segundo a 40 m de profundidad (10°7' N, y 76°16' W) en el PNNCPR (Fig. 1). Los especímenes de P. spinifera encontrados corresponden a longitudes entre 50,07 y 59,33 μι׳η (Tabla 1). La morfología de los ejemplares fue variable, observando formas de tentáculos, longitud y ancho distintos. Las medidas promedio de los tres especímenes de P. spinifera descritos en las aguas del Caribe colombiano pueden visualizarse en la Fig. 2e.
Tabla 1 Dimensiones de Pronoctiluca spinifera en mieras de diferentes localidades. N/R no se reporta la información.
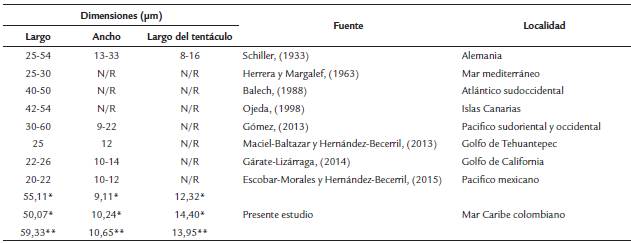
* Ejemplar encontrado en P1 (1 m)
** Ejemplar encontrado en P2 (40 m)
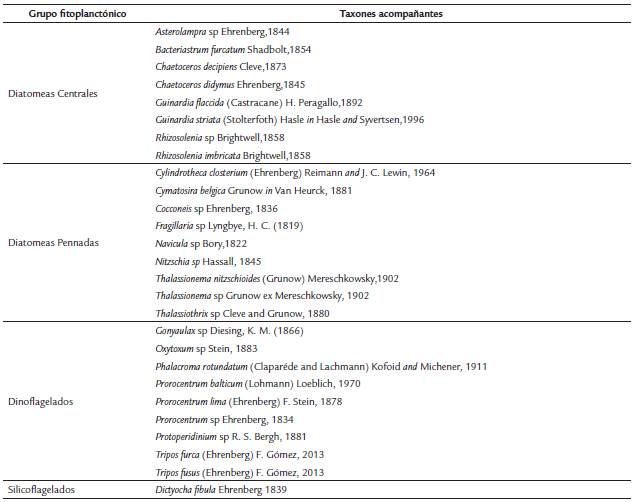
El elenco sistemático fitoplanctónico acompañante de P. spinifera fue compuesto por 27 taxa (Tabla 2), las diatomeas fueron las de mayor riqueza de taxa (nueve diatomeas pennadas y ocho diatomeas centrales), asimismo se encontraron nueve taxa de dinoflagelados y un taxón de silicoflagelados.
Variables fisicoquímicas: P. spinifera fue encontrada en aguas de superficie y profundidad con temperaturas de 30,34 °C y 27,69 °C, respectivamente; salinidades (PSU) de 35,6 y 36,38 y concentraciones de clorofi la a de 0,174 mg/m3 y 0,08 mg/m3.
DISCUSIÓN
P. spinifera es un dinoflagelado atecado capaz de alimentarse de otros organismos como dinoflagelados y zooplancton pequeños (Steidinger y Tangen, 1997). Los dinoflagelados atecados como los del género Pronoctiluca no poseen una pared celular funcional, por lo que la conservación de la morfología celular no es realizable, dado que los métodos de preservación de los especímenes no son adecuados y la mejor opción para la observación de estos son las muestras vivas (Larsen, 2002). Varias descripciones de dinoflagelados desnudos no conservables se basan en material preseleccionado en donde es frecuente que la taxonomía sea inadecuada; muchos de ellos son poco descritos y probablemente nunca puedan identificarse de manera adecuada (Larsen y Sournia, 1991; Larsen, 2002).
Sin embargo, se han desarrollado métodos de colecta y fijación con Tetraóxido de osmio (OsO4), el cual permite la observación de estructuras celulares de estos grupos, pero es poco utilizado por el riesgo que representa a la salud de los observadores, sus vapores altamente irritantes y tóxicos (Boltvskoy, 1995). Por estas razones, son poco estudiados, con escasos reportes en los listados de especies fitoplanctónicas (Larsen, 2002). En el caso del Mar Caribe se han reportado 331 especies de dinoflagelados tecados y 67 atecados (Wood, 1968), en el Caribe colombiano se han encontrado 183 y cinco respectivamente (Lozano-Duque et al., 2011; Malagón y Perdomo, 2013; Mancera-Pineda et al., 2014).
Los individuos encontrados mostraron acumulación de sustancias en el velo alimenticio, lo cual concuerda con descripciones morfológicas del género realizadas por Gómez, (2013b) quien evidencia la acumulación de alimento en el interior de P. spinifera.
En relación con el tamaño de P. spinifera, se determinó que las dimensiones del presente estudio están dentro del rango que caracterizó por primera vez la especie (Schiller, 1933), también coinciden con las medidas reportadas en las Islas Canarias, Pacifico sudoriental y occidental (Tabla 1). Por el contrario, los especímenes en el Caribe colombiano fueron de mayor tamaño que los registrados en el Mar Mediterráneo, Atlántico sudoccidental, Pacifico mexicano y Golfo de California (Tabla 1).
El registro de las medidas morfológicas son insumos clave para la identificación de las especies, el tentáculo es un carácter relevante del género Pronoctiluca; permite identificar a P. rostrata por la doble sección en el tentáculo, a P. pelagica por la disposición en diferentes ángulos del tentáculo, a P. acuta por la presencia de un tentáculo mediano o durante la división celular siendo ausente y P. spinifera por diferentes formas, tamaños y disposición del tentáculo (Gómez, 2013b). Schiller, (1933) describe la especie con medidas de largo, ancho y longitud del tentáculo; no obstante, trabajos como el de Herrera y Margalef, (1963), Balech, (1988) y Ojeda, (1998) se limitan a registrar solo una medida (largo del espécimen); en trabajos más recientes (Gómez, 2013b; Gárate-Lizárraga, 2014; Maciel-Baltazar y Hernández-Becerril, 2015) solo se reporta largo y ancho de las células.
El ejemplar de P. spinifera hallado a 40 m de profundidad presentó un mayor tamaño en contraste con los especímenes identificados en la superficie; esto permite plantear la existencia de condiciones favorables para la especie a esta profundidad. Desafortunadamente no existen estudios acerca de la dieta de P. spinifera; no obstante, en general los dinoflagelados heterotróficos poseen gran capacidad de adaptación trófica, pueden consumir aparte de dinoflagelados y zooplancton, diatomeas, bacterias y detritos (Jacobson y Anderson, 1986; Shanks y Walters, 1996; Hansen y Calado, 1999); además están asociados con el consumo de nieve marina (Tiselius y Kuylenstierna, 1996). En el Caribe colombiano, aunque son poco estudiados estos tópicos; Lozano et al., (2010b) en un estudio taxonómico del fitoplancton en el Caribe colombiano, mencionan que es probable que las diatomeas como grupo más abundante, mejore la disponibilidad alimenticia de las comunidades profundas, a esto se le agrega que los dinoflagelados se constituyeron en el grupo de mayor riqueza de especies, lo que permite plantear también un posible pastoreo sobre las diatomeas, favoreciendo la presencia de los dinoflagelados; hipótesis a comprobar en estudios posteriores.
Con relación a la distribución de P. spinifera, habita distintos mares y océanos, tanto cálidos como templados (Margalef, 1961; Gómez, 2005). Se ha encontrado en el Océano Índico (Wood, 1963; Taylor, 1964; Taylor, 1967; Sournia, 1972; Taylor, 1976); Mar Mediterráneo (Herrera y Margalef, 1963; Palau etal., 1991; Ignatiades, 1994; Gómez, 2003; Gómez y Gorsky, 2003; Gómez y Boicenco, 2004; Nassar y Khairy, 2014); Pacífico Occidental (Sur de China, Filipinas, Célebes y Sulu); Pacífico sudoriental Perú-Chile (Gómez, 2013b); Pacífico mexicano (Escobar-Morales y Hernández-Becerril, 2015); Golfo de Tehuantepec (Maciel-Baltazar y Hernández-Becerril, 2013); Baja California Sur (Gárate-Lizárraga et al., 2007); Golfo de California (Gárate-Lizárraga, 2014); Océano Atlántico nororiental (Wood, 1968); Atlántico sudoccidental (Balech, 1988); Islas Canarias (Ojeda, 1996; 1998; Gil-Rodríguez et al., 2003); Golfo de México (Zamudio et al., 2013) y en la Bahía de Guanabara, en Brasil (Vieira de Rezende et al., 2015). Para las aguas territoriales colombianas, anterior a esta publicación, no existe registro de esta especie; en el Pacífico colombiano no se ha reportado la presencia de P. spinifera; mientras que en el Caribe colombiano se amplía su distribución, hallándose en el PNNCPR.
El género Pronoctiluca Fabre-Domergue, (1889) está integrado por dinoflagelados heterotróficos, con morfología piriforme o fusiforme. Las especies de este género poseen un tentáculo bien desarrollado en la parte anterior, no estriado, móvil, pero no retráctil, con dos flagelos largos insertados; uno dirigido hacia la parte posterior de la célula y otro en dirección transversal, el núcleo suele ser un dinocarion típico (Balech, 1988; Sournia, 1986; Gómez, 2013b). El género se caracteriza por la locomoción y los cambios rápidos en su forma celular (Gómez et al., 2010), mostrando afinidad con Noctiluca scintillans (Macartney) Kofoid, con quien comparte similitud en la movilidad del tentáculo en todas las direcciones, la falta de cíngulo y las numerosas vacuolas (Gómez, 2013b). Sin embargo, Noctiluca posee un flagelo, mientras que Pronoctiluca exhibe dos (Fensome et al., 1993).
Pronoctiluca es considerado un género primitivo de posición sistemática incierta (Patterson et al., 1991). Fue ubicado inicialmente en la familia Nocticulaceae (Kent 1881), y posteriormente excluido por no tener los caracteres definidos para la familia (Cachon y Cachon, 1967). Schiller, (1933) lo incluyó en la familia Pronoctilucaceae (M. Lebour. 1925) del orden Gymnodiniales; lo que evidencia que el género presenta problemas al determinar su posición sistemática. En la actualidad está ubicado en el orden Noctilucales, familia Protodiniferaceae (Kofoid y Swezy 1921), (Gómez, 2013b; Worms, 2017; Guiry y Guiry, 2018;), junto con otras cuatro especies y una variedad: P. acuta (J. Schiller, 1933; Lohmann, 1912;), P. pelagica (Fabre-Dom, 1889), P. rostrata (F.J.R. Taylor, 1976), P. Phaeocysticola (Scherffel Pavillad, 1922) y P. Acuta var. Curvata (Lohmann, 1912; J. Schiller,1933).
P. spinifera presenta caracteres similares con otras especies de Pronoctiluca (Gómez, 2013b) durante observaciones en la división celular de P. spinifera, caracterizó un extremo posterior puntiagudo y alargado, indicando similitud con la morfología de P. acuta; generando una hipótesis en la que P. acuta puede constituir una variación morfológica de P. spinifera. Además P. pelagica posee un contorno piriforme, carece de flagelos y está cubierto con capas hialinas, con gran parecido a la forma intermedia de P. spinifera, pero la capacidad de encriptación es la que determina a P. pelagica (Takayama, 1998) utilizando microscopia electrónica de barrido, logró describir dos secciones del tentáculo cilíndrico de P. rostrata, diferenciando el tentáculo de P. spinifera por ser reducido. En ese orden de ideas, es incierta la legitimidad de P. rostrata como especie separada, dada la posibilidad de ser un morfotipo de P. spinifera.
Del elenco sistemático acompañante a P. spinifera (Tabla 2), son taxa regularmente reportadas en el Caribe colombiano (Arias y Duran, 1984; Franco-Herrera et al., 2006; Lozano et al., 2010a; Lozano et al., 2010b; Lozano et al., 2010c; Lozano et al., 2011; Salon-Barros y Cañón-Páez, 2013). Dentro de las diatomeas, las destacables por su abundancia fueron Navicula sp, asociada comúnmente a zonas costeras con influencia de descargas continentales (Lange-Bertalot, 2001) y Thalassionema nitzchioides, al encontrarse generalmente con alta abundancia en sistemas estuarinos (Cassis y Avaria, 2002). Por los dinoflagelados, Prorocentrum balticum y P. lima, quienes han sido responsables de recientes florecimientos algales en la Bahía de Cartagena (Salon-Barros et al., 2016); asimismo, las especies Tripos furca y T. fusus, en donde es importante resaltar el reemplazo del género Ceratium Schrank, 1973 por Tripos Bory, 1823 (Gómez, 2013a); han sido asociadas a florecimientos algales nocivos (Vargas-Montero y Freer, 2004), ellas poseen características psicrófilas que le permiten mayor influencia en aguas oceánicas que en costeras (Carbonell, 1979).
La temperatura y salinidad del agua marina estructuran físicamente la columna de agua y regulan gran parte de sus procesos biogeoquímicos (Millero, 2013), de allí la relevancia en el registro de las condiciones ambientales en PNNCPR al encontrar a P. spinifera. Al momento de revisar los reportes de esta especie a escala global, algunos simplemente no reportan condiciones oceanográficas (Wood, 1968; Palau et al., 1991; Ojeda, 1996; Gil-Rodríguez et al., 2003; Gómez, 2003; Gárate-Lizárraga, 2007; Maciel-Baltazar y Hernández-Becerril, 2013; Nassar y Khairy, 2014; Escobar-Morales y Hernández- Becerril, 2015), por su parte Margalef, (1961) evidencia un amplio rango de temperatura de adaptación de esta especie y la describe como cosmopolita de aguas templadas (10 - 20 °C) y aguas cálidas (20 - 25 °C).
Adicional a eso, en el Océano Pacífico (Gómez, 2013b) y en el Golfo de México (Zamudio et al., 2013) esta especie fue hallada en aguas con temperaturas de 26 - 30°C y 28,8 °C respectivamente; temperaturas símiles a las encontradas en PNNCPR. Paralelamente, los valores de salinidad en los puntos estudiados en PNNCPR son propios de aguas oceánicas (Lalli y Parson, 1997), condición con la cual P. spinifera es habitualmente reportada (Gárate-Lizárraga, 2014), hasta llegar a encontrarse en salinidades de 38 en el Mediterráneo, lo que muestra que puede establecerse en gran variedad de ambientes oceánicos templados y tropicales, validando su condición cosmopolita.
La concentración clorofila-a como indicador de biomasa fitoplanctónica analizada bajo el criterio de Barocio-León, (2006) quien categorizó las aguas marinas en oligotróficas (<0,2 mg/m3), mesotróficas (>0,2 mg/ m3 y 2 mg/m3) y eutróficas (>2,0 mg/m3), indican que las características tróficas de las aguas donde se encontró P. spinifera corresponden a un estado oligotrófico, siendo muy distinto a los valores de biomasa fitoplanctónica encontrados por Franco-Herrera et al., (2006) en el Golfo de Salamanca frente a las costas del Magdalena, quienes hallaron concentraciones altas de clorofila a en distintas épocas del año debido a la alternancia entre la influencia de la pluma del Rio Magdalena en época lluviosa (septiembre-noviembre) y la surgencia costera en épocas de vientos alisios (enero-marzo). Las concentraciones de clorofila-a encontrados durante el presente estudio son también más bajas que las encontradas en la Bahía de Santa Marta a diferentes profundidades (Mancera-Pineda et al., 2013).
Muller-Karger et al., (1989) por medio de análisis de series de tiempo con el sensor remoto Coastal Zone Color Scanner (CZCS) establecieron para las aguas del Caribe la condición oligotrófica, coherente con los resultados in situ del presente estudio. No obstante, de acuerdo con la propuesta de regionalización dinámica del Caribe, Cañón-Páez y Santamaría, (2010) empleando sensores remotos (SeaWiFS, MODIS-AQUA y MERIS) establecieron que para la zona de PNNCPR confluyen la región mesotrófica del Golfo del Darién y la región oligotrófica Giro Panamá-Colombia, lo que permite plantear en función a los datos encontrados de clorofila-a, la posible influencia del Giro Panamá-Colombia por la advección de aguas superficiales sobre PNNCPR. Estos resultados estarían de acuerdo con los de un reciente estudio en el que utilizando imágenes mensuales del sensor MODIS entre 2003 y 2013, encontraron que solo tres áreas del Mar Caribe colombiano presentan valores relativamente altos de clorofila, el Golfo de Urabá, la desembocadura del Río Magdalena y la península de la Guajira (Coronado-Franco et al., 2018).
CONCLUSION
Dada la dificultad del estudio de los dinoflagelados desnudos y los pocos reportes para el Caribe colombiano, el registro de P. spinifera en aguas superficiales y subsuperficiales, toma relevancia. Se han descrito aspectos morfológicos, encontrándose que su tamaño es mayor en aguas subsuperficiales del PNNCPR y que las condiciones ambientales de las aguas en donde fue encontrada son de carácter oceánico y oligotrófico. Este primer reporte de P. spinifera en el Caribe colombiano, no solo amplía el conocimiento sobre la distribución geográfica de la especie, sino que a la vez muestra la necesidad de desarrollar estudios sobre dinoflagelados desnudos en aguas colombianas. Para esto se requiere el desarrollo de nuevos métodos de preservación y observación apropiados para este tipo de organismos. Así mismo es necesario estudiar la dinámica trófica de los dinoflagelados heterotróficos, con el propósito de mejorar el conocimiento de la diversidad biológica y funcional de las aguas territoriales colombianas y a su vez de ecosistemas estratégicos como PNNCPR, en especial de los organismos planctónicos que son la principal base alimenticia de los sistemas oceánicos.


