











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Los escarabajos coprófagos constituyen uno de los grupos de insectos mejor conocidos a nivel mundial tanto en su taxonomía como en la conformación e importancia ecológica de sus ensamblajes de especies, los cuales responden como unidad funcional a características específicas de los paisajes naturales y de aquellos alterados por la acción antrópica, lo que los constituye como uno de los grupos más utilizados en estudios de conservación y recuperación ecosistémica por su carácter bioindicador (Halffter y Matthews, 1966; Hanski y Cambefort, 1991; Halffter y Favila, 1993; Escobar et al., 2005; Fuentes y Camero, 2006; Nichols et al., 2008; Simmons y Ridsidill, 2011). Las principales funciones ecológicas de este grupo de insectos son la contribución al reciclaje de nutrientes, la fertilización del suelo y la contribución a procesos edafogenéticos, la dispersión de semillas y su relación ecológica directa con vertebrados y otros grupos de artrópodos e invertebrados (Hanski y Cambefort, 1991; Nichols et al., 2008). La fauna de escarabajos coprófagos en Colombia está representada por 40 géneros y cerca de 300 especies (Noriega et al., 2015), lo cual representa alrededor del 60 % de la diversidad de géneros y el 30 % de especies de la fauna Neotropical (Philips, 2011).
La Guyana colombiana hace parte de la provincia biogeográfica occidental del escudo guyanés, subdividida en cinco distritos y 12 áreas de endemismos y conformada por una serie de sierras, serranías, cerros y sabanas que estuvieron unidas en el Terciario a los escudos guyanés y brasileño (Huber, 1994; Etter, 1997). Estas serranías están configuradas por afloramientos rocosos, bosques y sabanas con elementos vegetales comunes a todas ellas y una gran cantidad de especies endémicas como el caso de las bromeliáceas del género Navia, árboles del género Roupala y las emblemáticas Vellozia lithophila y Paepalanthus formosus (Hernández et al., 1992; Berry et al., 1995). Entre las sierras y serranías de origen guyanés y con elementos florísticos y faunísticos compartidos, sobresalen las de Chiribiquete, La Macarena, El Tuparro, Araracuara, Taraira, Naquén, Lajas de Guainía y La Lindosa (Gentry, 1982; Cortés y Franco, 1997).
La vegetación típica de la Serranía de La Lindosa está incluida en una serie de bosques bajos, arbustales y sabanas con mayores afinidades florísticas con las Serranías de La Macarena y El Tuparro que con las Serranías de Araracuara, Chiribiquete y las Lajas de Guainía (Giraldo-Cañas, 2001). Los bosques bajos están conformados por comunidades vegetales de pocos individuos, con alturas de hasta 8 m y de dosel irregular, dando la apariencia de bosques ralos con poca cobertura del dosel. Los arbustales se localizan sobre los afloramientos rocosos y están compuestos por pocas especies arbustivas en general pertenecientes a las familias Bonnetiaceae, Clusiaceae, Rubiaceae, Annonaceae, Asteraceae, Melastomataceae, Myrtaceae y Tepuianthaceae y especies herbáceas de los géneros Vellozia, Axonopus, Schizachyrium, Navia, Aechmea, Anemia, Selaginella y Diacidia.
Las sabanas pueden ser de varios tipos, pero en general con presencia de algunas especies de Trachypogon, Andropogon, Panicum, Paspalum, Axonopus, Otachyrium, Xyris, Utricularia, Bulbostylis, Calea, Aeschynomene, Clitoria, Desmodium, Nautilocalyx, Hyptis, Siphantera, Sipanea, Sipaneopsis y Drosera entre otras (Giraldo-Cañas, 2001).
La composición florística de sierras y serranías de la Guyana colombiana y sus nexos biogeográficos son parte de la mayoría de las investigaciones realizadas en esta zona del país, siendo muy escasos los trabajos en grupos animales y particularmente en los grupos de insectos. Para el caso de los escarabajos estercoleros son pocas las investigaciones en biodiversidad y los trabajos más puntuales abarcan las serranías de El Tuparro (Quintero et al., 2007) y Chiribiquete (Martín-Piera y Fernández-Torres, 1996; Pulido et al., 2003). La ausencia de trabajos referentes a la composición de la fauna de insectos y en especial aquellos que contemplen los escarabajos estercoleros en la diversidad de ecosistemas de la Serranía de La Lindosa, animan al conocimiento de la biodiversidad regional más allá de los aspectos descriptivos, por lo que el presente trabajo, busca determinar la conformación de los ensamblajes de especies de escarabajos coprófagos en cuatro estaciones de muestreo de la Serranía de La Lindosa bajo diferentes coberturas vegetales y establecer sus afinidades faunísticas con otras áreas ya estudiadas de la Guyana Colombiana..
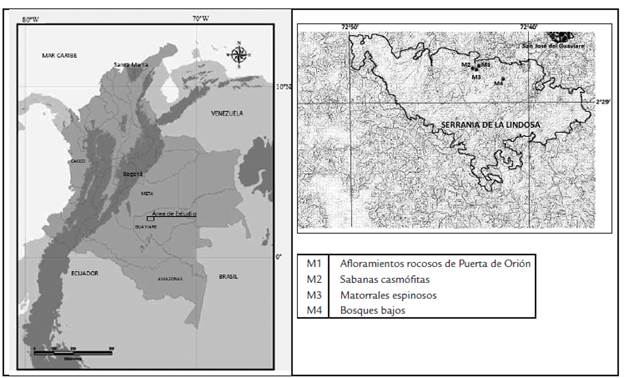
MATERIALES Y MÉTODOS
El trabajo de campo se realizó en la zona norte de la Serranía de La Lindosa (Departamento del Guaviare) en cuatro puntos de muestreo: afloramientos rocosos de la Puerta de Orión (M1), ubicados en los 2o30'53"N y 72o42'33"W a 290 m de altura; sabanas casmófitas (M2) ubicadas en los 2o30'45"N y 72o42'52"W a 320 m; los matorrales espinosos (M3) ubicados en los 2o30'42"N y 72o42'41"W a 345 m y bosques bajos (M4) ubicados en los 2o30'12"N y 72o41'17"W a 315 m.s.n.m (Fig. 1). En cada una de las cuatro estaciones se utilizaron trampas de caída cebadas con coprocebo humano durante 48 horas siguiendo las recomendaciones de Howden y Nealis, (1975) y Howden y Young, (1981). Se instalaron diez trampas de caída separadas 10 m una de otra en transectos de muestreo aleatorio para cubrir áreas de 100 m x 100 m al interior de matrices homogéneas de cada cobertura vegetal para evitar los efectos de influencia por coberturas vegetales adyacentes, manteniendo las condiciones similares del mantillo y de la capa superficial del suelo como carácter edáfico determinante en la conformación de los gremios de escarabajos. Los especímenes colectados se etiquetaron y transportaron al Departamento de Biología de la Universidad Nacional de Colombia para ser depositados en la colección de referencia del Departamento de Biología luego de ser determinados a nivel de especie siguiendo las claves taxonómicas de Arrow, (1933); Halffter y Martínez, (1977); Howden y Young, (1981); Edmonds, (1994; 2000); Medina y Lopera-Toro, (2000); Vitolo, (2000); Kohlmann y Solís, (1997; 2001); Solís y Kohlmann, (2002; 2004; 2013); Edmonds y Zidek, (2004; 2010); González et al., (2009); Camero, (2010); Vaz de Mello et al., (2011); Sarmiento y Amat, (2014) y Cupello, (2018) y por confrontación con especímenes de referencia confirmando los nombres válidos en Species 2000 y Catálogo ITIS (Roskov et al., 2014). Para el análisis de los datos se evaluó la eficiencia del muestreo mediante los estimadores de abundancia ACE, CHAO 1 y COLE en las curvas de acumulación de especies de los puntos de muestreo con el uso del programa EstimateS v. 9.0 (Colwell, 2013). La comparación entre localidades se efectúo mediante análisis de Box plot y dendrogramas de similaridad de Bray-Curtis (Brower y Zar, 1984) a partir de los datos de presencia/ausencia de especies siguiendo el método de single-linkage y por la comparación entre los valores de diversidad medida por los índices descriptivos de Brillouin (B) de dominancia (D) y equitatividad de Simpson (1-D) y de Diversidad Beta de Wittaker (aW) a partir de los valores de abundancia de las especies mediante el uso del programa PAST v. 2.15 (Hammer et al., 2001; Magurran, 2004) que se validaron mediante pruebas de Kruskal-Wallis (Zar, 1999). Finalmente, se evaluó la afinidad de la fauna colectada en los sitios de muestreo mediante un Análisis de Correspondencia (DCA) y se realizó el modelamiento geográfico para otras áreas del país con condiciones climáticas similares a las condiciones climáticas promedio del área de estudio, seleccionando las variables más relevantes mediante el Ecological Niche Factor Analysis (ENFA), (Hirzel et al., 2002) a partir de las variables bioclimáticas libremente disponibles en Worldclim (Hijmans et al., 2005).
RESULTADOS
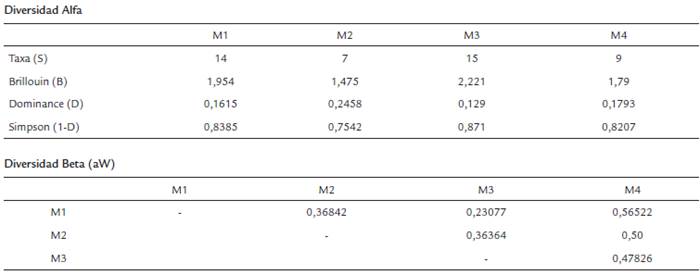
Se colectaron 1025 ejemplares de Scarabaeinae pertenecientes a ocho géneros, 22 especies. Las localidades con mayor número de especies colectadas fueron las ubicadas en los afloramientos rocosos de Puerta de Orión (M1) y los matorrales espinosos (M3) con 13 especies, seguidas por los bosques bajos (M4) con diez y las sabanas casmófitas (M2) con seis especies (Tabla 1). Existe diferencias en los valores de diversidad entre los cuatro puntos de muestreo (Kruskal-Wallis, H=7,05, p=0,05); la mayor diversidad faunística se obtuvo en la estación de muestreo localizada en los matorrales espinosos (M3) (B=2,221) y la menor en las sabanas casmófitas (M2) (B=1,475), la cual también presenta los mayores valores de dominancia de especies (D=0,2458) y la menor equitatividad (1-D=0,7542) (Tabla 2). El mayor recambio de especies se encontró entre los puntos de muestreo de los bosques bajos (M4) y los afloramientos rocosos de Puerta de Orión (M1) (aW=0,56522) y la menor, entre esta última y los matorrales espinosos (M3) (aW=0,31034).
Tabla 1 Abundancia relativa de la fauna de escarabajos coprófagos colectada en los cuatro puntos de muestreo de la Serranía de La Lindosa-Guaviare.
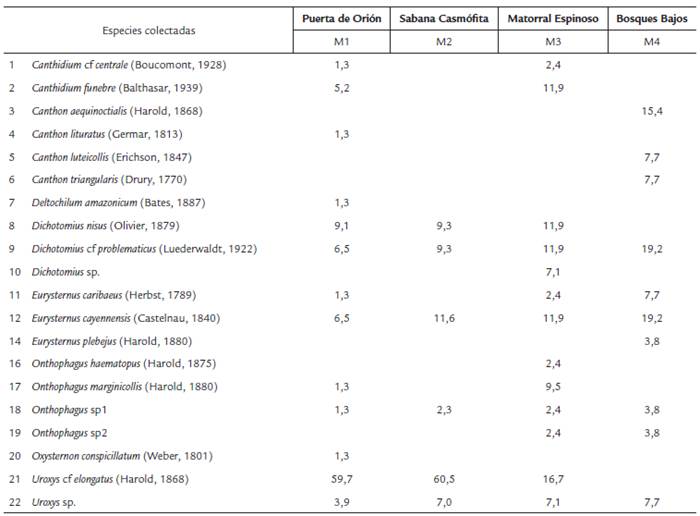
Tabla 2 Estimadores de Diversidad Alfa y Beta para los puntos de muestreo de escarabajos coprófagos en la Serranía de La Lindosa-Guaviare.
Los ensamblajes de escarabajos estercoleros están compuestos por gremios de especies que se diferencian en tres grupos según la forma como realizan su nidificación sobre las fuentes de alimento: paracópridos o cavadores, telecópridos o rodadores y endocópridos o residentes, que a la vez son estrategias importantes en la competencia por recursos dentro de estos gremios (Halffter, 1991; Hanski y Cambefort, 1991). Las especies más abundantes en la mayoría de las estaciones de muestreo fueron Uroxys cf elongatus, Dichotomius nisus y Dichotomius cf problematicus que corresponden a especies de hábito cavador. Otras especies de menor abundancia fueron encontradas en todas las estaciones como las especies cavadoras Onthophagus sp1 y Uroxys sp y las especies endocópridas Eurysternus cayennensis y Eurysternus caribaeus. En general, se colectó un mayor número de especies cavadoras que endocópridas o rodadoras en todas las estaciones de muestreo lo que podría considerarse como un rasgo característico de los ensamblajes de escarabajos coprófagos a nivel de paisaje.
La eficiencia de los muestreos en las distintas localidades medida por los estimadores ACE, CHAO 1 y COLE (Fig. 2) mostró grados muy similares, siendo más eficiente en la estación ubicada en los matorrales espinosos (M3) en la cual la completitud de especies fue cercana al 95 %. Para las demás localidades, se encontraron diferencias cercanas al 90 % entre la fauna colectada y la esperada y tendencia a estabilización de las curvas a partir de las diez unidades muestrales. La tendencia a valores cercanos a cero de los estimadores que evalúan las especies representadas por pocos individuos en los muestreos (Singletons y Doubletons), indica que el número de unidades muestrales utilizadas en los diferentes puntos fue suficiente para colectar la mayoría de la fauna presente en cada estación.
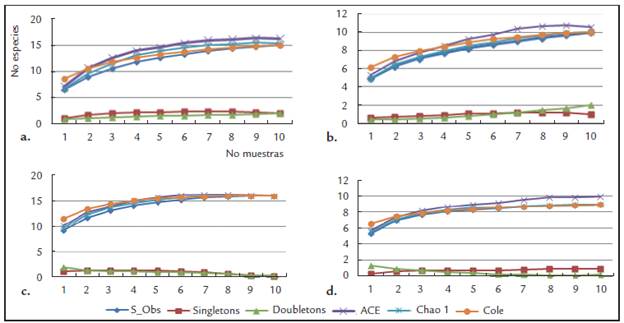
Figura 2 Estimadores de abundancia para las curvas de acumulación de especies en los cuatro puntos de muestreo en la Serranía de La Lindosa-Guaviare. a: afloramientos rocosos de Puerta de Orión, b: sabana casmófita, c: matorral espinoso, d: bosques bajos.
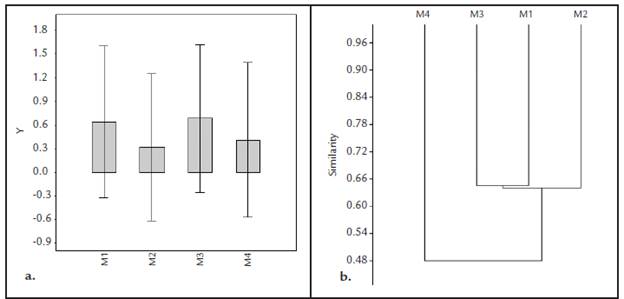
No obstante, no se encontraron diferencias significativas en la composición faunística de los cuatro puntos de muestreo según el análisis Box plot (Fig. 3a), la similaridad entre puntos de muestreo según el diagrama de Bray-Curtis (Fig. 3b), mostró mayor relación entre aquellas ubicadas en los afloramientos de Puerta de Orión (M1) y los matorrales espinosos (M3) (p>0,66) y menor relación entre estas y sabana casmófita (M2) y los bosques bajos (M4). El mayor número de las especies se comparte entre los puntos de muestreo M1, M2 y M3 que corresponden a los afloramientos rocosos de la Puerta de Orión, la sabana casmófita y el matorral espinoso, lo cual explica la marcada diferencia en el dendrograma de similaridad entre estos tres puntos de muestreo y la estación ubicada en los bosques bajos que cuenta con una composición faunística particular (Tabla 1).
DISCUSIÓN
COMPOSICIÓN FAUNÍSTICA Y DIVERSIDAD
Los resultados muestran la baja diversidad propia de este tipo de zonas derivadas de formaciones geológicas guyanesas y de gran exigencia climática (Gentry, 1982), cuyas principales características ecofisiológicas la conforman los bosques bajos, las sabanas y los afloramientos rocosos donde la tipología mayormente arenosa de los suelos y la poca humedad retenida explican las bajas densidades poblacionales y la baja diversidad de escarabajos estercoleros (Lumaret y Kirk, 1991). Valores similares en la diversidad de especies de este grupo ya han sido registrados para algunos sectores de la Serranía de Chiribiquete (Martín-Piera y Fernández-Torres, 1996; Pulido et al., 2003) y para afloramientos rocosos de la Guyana Francesa (Feer, 2012).
A pesar del bajo número de especies registradas, se encontraron diferencias en la composición faunística de escarabajos coprófagos en los puntos de muestreo (Tabla 1), en la mayor riqueza de especies encontrada en los afloramientos rocosos (M1) y el matorral espinoso (M3) en comparación a las demás estaciones de muestreo y en la mayor cantidad de especies no compartidas de los afloramientos rocosos de Puerta de Orión (M1). Más del 54 % de la fauna colectada es compartida; las especies más comunes en los cuatro sitios de muestreo incluyen a las tres especies más abundantes: Uroxys cf elongatus, Dichotomius nisus y Dichotomius cf problematicus, que junto con Eurysternus cayennensis, Eurysternus caribaeus, Onthophagus sp1 y las dos especies del género Uroxys pueden considerarse como aquellas que son características de los ensamblajes de escarabajos coprófagos de la zona norte de la Serranía de La Lindosa.
La unidad biogeográfica, la particularidad de la biota y el alto grado de endemismo de la Guyana colombiana ha sido considerada en varias ocasiones tanto desde la historia de su geología, como por los nexos de su diversidad biológica con los escudos guyanés y brasileño (Lescure y Boulet, 1985; Kubitzki, 1989; Hernández et al., 1992; Cortés y Franco, 1997; Giraldo-Cañas, 2001). Varias de las especies de escarabajos coprófagos encontradas en la Serranía de La Lindosa, han sido registrados anteriormente para la provincia biogeográfica Occidental del escudo Guyanés: C. centrale, C. funebre, C. aequinoctialis, C. luteicollis, D. amazonicum, D. problematicus, E. caribeaus, E. cayennensis, O. haematopus y O. conspiscillatum, fueron colectadas en algunas zonas de la Serranía de Chiribiquete (Pulido et al., 2003), mientras que C. centrale, C. triangularis, E. caribeaus, E. cayennensis y O. haematopus en bosques de la Guyana Francesa (Feer, 2012). C. aequinoctialis, C. luteicollis, D. problematicus, E. caribaeus y O. haematopus son especies que han sido registradas en otras zonas de estudio del Departamento del Guaviare (Escobar, 2000), lo que representa un gran número de especies de amplia distribución y comunes a toda la provincia biogeográfica; muchas de las especies no determinadas en estos estudios especialmente de los géneros Canthidium, Canthon, Dichotomius, Onthophagus y Uroxys, podrían corresponder a especies aún no descritas que ampliarían el número de especies endémicas de los ecosistemas regionales.
Para el caso de los ensamblajes de especies colectados en los puntos de muestreo de la Serranía de La Lindosa, la mayoría de las especies son cavadoras y son muy pocas las especies endocópridas y rodadoras. Las especies cavadoras pertenecen a los géneros Canthidium, Dichotomius, Onthophagus, Oxysternon y Uroxys, las endocópridas al género Eurysternus y las rodadoras a los géneros Canthon y Deltochilum. La dominancia de especies cavadoras es propia de ambientes naturales donde los recursos son escasos y en sitios donde favorece la tipología edáfica (Halffter y Edmonds, 1982; Howden et al., 1991; Fuentes y Camero, 2006). Probablemente, la baja intervención antrópica de los cuatro ambientes en donde se establecieron los puntos de muestreo sea una de las causas de la escases de recursos para este tipo de fauna y que la tipología del suelo favorezca además, el predominio de especies con hábitos de nidificación cavadora. La influencia antrópica como factor importante en el establecimiento de los gremios de los escarabajos estercoleros, ya ha sido reportada en estudios ecológicos en otras zonas del país (Fuentes y Camero, 2006; Pardo-Locarno y Camero, 2014).
La figura 4 muestra las afinidades de la fauna colectada en los sitios de muestreo debidas a la distribución o a la densidad poblacional (Fig. 4a) y las áreas en Colombia cuya similaridad en las condiciones climáticas podrían potencialmente albergar una fauna similar como serían algunas zonas específicas de la Serranía de La Macarena, la Serranía de Chiribiquete y Araracuara o zonas cercanas a los afloramientos rocosos de El Tuparro (Fig. 4b). No obstante, la amplia distribución de varias de las especies colectadas en todos los sitios de muestreo como D. nisus o U. elongatus, sobresale la afinidad de C. fúnebre, O. marginicollis y C. lituratus por los afloramientos rocosos de Puerta de Orión (M1), D. problematicus por la sabana casmófita (M2), Onthophagus sp1 y O. haematopus por los matorrales espinosos (M3) y C. aequinoctialis, C. luteicollis y C. triangularis por los bosques bajos (M4). Las afinidades de la fauna por los sitios de muestreo podrían deberse, más que a la oferta de recursos, a las condiciones microclimáticas y edáficas favorables para la nidificación o reproducción (Halffter y Edmonds, 1982; Hanski y Cambefort, 1991) que se reflejan en el recambio de especies de los bosques bajos con todos los demás ecosistemas (Tabla 2).
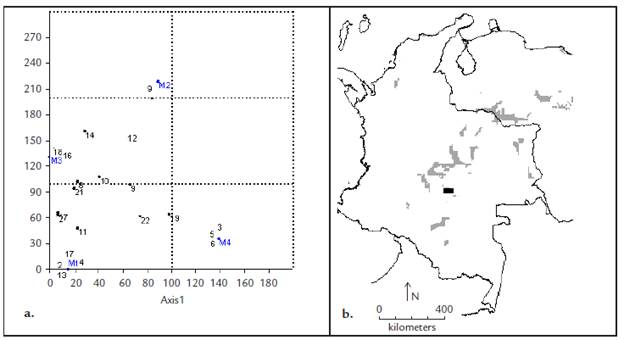
Figura 4 Análisis de correspondencia (DCA) entre la fauna colectada y los puntos de muestreo (a) y resultado del modelamiento de las zonas climáticas similares de Colombia (b) a partir de las condiciones climáticas promedio de la zona de estudio (recuadro). El número asignado a cada especie corresponde a la tabla 1 (Eigenvalues: axis 1=0,51, axis 2=0,08).
SINOPSIS DE LA FAUNA COLECTADA
La sinopsis de la fauna colectada se realizó teniendo en cuenta la presencia en los sitios de muestreo y la distribución geográfica actual en Colombia de acuerdo con los compendios más recientes y fuentes bibliográficas de trabajos realizados en otras zonas de la Guyana colombiana: El género Canthidium (Erichson, 1847) es uno de los géneros más diversos en Colombia para el cual se estiman actualmente 27 especies, muchas de ellas de amplia distribución geográfica (Noriega et al., 2015). Las dos especies colectadas en la zona de estudio: Canthidium cf centrale y Canthidium funebre, fueron encontradas en los afloramientos rocosos de Puerta de Orión (M1) y en los matorrales espinosos (M3). Canthidium centrale se ha colectado en Colombia en la zona montañosa de los Departamento de Antioquia, Chocó, Nariño hasta alturas de 1800 m.s.n.m, en sabanas y selvas de los Departamentos del Vaupés y Vichada (Medina et al., 2001; Quintero et al., 2007; Pardo-Locarno y Camero, 2014) y en inmediaciones de las Serranías de Chiribiquete y de las Guyanas (Pulido et al., 2003; Feer, 2012; Larsen, 2013).
El género Canthon (Hoffmannsegg, 1817), es otro de los géneros con mayor diversidad en Colombia. Se han registrado 34 especies en el país (Noriega et al., 2015), pero este número podría ser mucho mayor debido a la gran cantidad de registros depositados en diferentes colecciones que no han sido determinados a nivel de especie. Canthon lituratus fue colectada solo en los afloramientos rocosos de Puerta de Orión, pero su distribución geográfica hasta el momento en Colombia abarca los Departamentos de Bolívar, Tolima y Vaupés en alturas inferiores a los 500 m.s.n.m. (Medina et al., 2001). Las otras tres especies del género: C. aequinoctialis, C. luteicollis y C. triangularis, fueron encontradas únicamente en la estación de muestreo de los bosques bajos (M3); aunque C. aequinoctialis ha sido registrada en bosques bajos de los Andes (Fuentes y Camero, 2006; Pardo-Locarno y Camero, 2014), las dos especies restantes solo se han registrado en selvas bajas de la Orinoquía y la Amazonía colombiana y en zonas de la Guyana biogeográfica (Escobar, 2000; Medina et al., 2001; Pulido et al., 2003; Quintero et al., 2007; Medina y Pulido, 2009; Feer, 2012; Larsen, 2013).
Las dos especies reconocidas del género Dichotomius se colectaron en la mayoría de las estaciones de muestreo: Dichotomius nisus en las estaciones M1, M2 y M3 y Dichotomius cfproblematicus en todas las cuatro; otra especie no identificada del género se colectó solo en la estación ubicada en los matorrales espinosos (M3). El género Dichotomius (Hope, 1838) es muy diverso en Colombia; a la fecha se han registrado 32 especies (Sarmiento y Amat, 2014), aunque un gran número de ejemplares sin identificación reposan en muchas colecciones entomológicas. D. nisus se ha registrado en los departamentos de Casanare, Cundinamarca, Guainía, Meta y Vichada hasta los 1200 m.s.n.m., mientras que D. problematicus tan solo en los Departamentos de Guaviare y Meta en alturas menores a los 350 m.s.n.m. (Escobar, 2000; Medina et al., 2001; Medina y Pulido, 2009) y en la Serranía de Chiribiquete (Pulido et al., 2003).
Para el género Eurysternus (Dalman, 1824) se han registrado 17 especies en Colombia (Camero y Lobo, 2010). Se colectaron pocos ejemplares de las tres especies registradas en las estaciones de muestreo de la Serranía de La Lindosa: E. cayennensis se colectó en todas las estaciones de muestreo, E. caribaeus en las estaciones M1, M3 y M4 y E. plebejus solo en la estación de bosques bajos (M4). No se registraron otras especies reportadas para el Departamento del Guaviare, especialmente aquellas de mayor tamaño y amplia distribución como E. foedus o E. hamaticollis. Tanto E. caribaeus como E. cayennensis han sido reportadas en otras zonas del Departamento del Guaviare y en los trabajos de Pulido et al., (2003); Feer, (2012); Medina y Pulido, (2009) y Larsen, (2013) para la Guyana Occidental.
El género Onthophagus (Latreille, 1802), es otro de los géneros más diversos en Colombia y cuenta con 31 especies (Noriega et al., 2015), aunque existen muchos ejemplares no identificados en las colecciones de referencia que podrían aumentar en gran número la diversidad de especies de este género para Colombia. En las estaciones de muestreo se colectaron cuatro especies, dos de ellas plenamente identificadas: O. haematopus en la estación de matorrales espinosos (M3) y O. marginicollis en los afloramientos de Puerta de Orión (M1) y en los matorrales espinosos (M3). Ambas especies ya han sido registradas en trabajos anteriores para la Guyana Occidental y el Departamento del Guaviare (Escobar, 2000; Pulido et al., 2003; Feer, 2012; Larsen, 2013) como para bosques andinos en alturas menores a los 300 m.s.n.m. (Medina et al., 2001; Fuentes y Camero, 2006). Una de las dos especies no identificadas se encontró en todas las estaciones de muestreo, mientras que la otra en las estaciones de los matorrales espinosos (M3) y en los bosques bajos (M4).
Finalmente, el género Uroxys (Westwood, 1842) está representado en Colombia por 15 especies reconocidas (Noriega et al., 2015); sin embargo, también existe gran cantidad de especímenes no identificados en las colecciones de referencia que podrían elevar el número de especies en Colombia. En la zona de muestreo se colectaron dos especies: Una no identificada con distribución en todas las estaciones de muestreo y Uroxys cf elongatus que se encontró en las estaciones de muestreo localizadas en los afloramientos de Puerta de Orión (M1), en las sabanas casmófitas (M2) y en los matorrales espinosos (M3). U. elongatus ya había sido registrada con distribución en la Orinoquia colombiana (Medina y Pulido, 2009)
CONCLUSIONES
La baja riqueza de especies encontrada para la zona norte de la Serranía de La Lindosa corresponde con la diversidad esperada para zonas naturales de baja intervención que presentan escasos recursos y grandes exigencias climáticas. Dichotomius cf. problematicus, Eurysternus cayennensis, Dichotomius nisus, Onthophagus marginicollis y Uroxys elongatus hacen parte de un pequeño número de especies encontradas bajo diferentes coberturas vegetales que podrían considerarse como las especies típicas de los ensamblajes de especies de la zona.
Dichotomius nisus, Dichotomius cf problematicus y Uroxys elongatus son las especies que presentaron las mayores densidades poblacionales bajo todos los tipos de cobertura vegetal, y junto con las especies de los géneros Onthophagus, Oxysternon y Uroxys encontradas en las distintas zonas, conforman el gremio dominante de nidificación cavadora, más favorecido por las condiciones climáticas y edáficas.
La mayoría de las especies colectadas han sido reportadas anteriormente en estudios realizados en la región, lo cual podría apoyar la idea de la biota de la Guyana Occidental como unidad biogeográfica, con mayores nexos con los escudos guyanés y brasileño que con la biota andina.


