











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El chile o ají (Capsicum spp.) incluye tanto plantas herbáceas anuales o bianuales como plantas perennes y arbustivas, pertenece a la familia Solanaceae y tiene sus orígenes en la región tropical de América del Sur, en lo que hoy se conoce como República de Bolivia (Olmstead et al., 2008). El género Capsicum está conformado por 25 especies (Baral y Bosland, 2002), de las cuales cinco son las más cultivadas, siendo Capsicum annuum la primera a nivel mundial (Yoon et al., 2004). El cultivo del ají es uno de los más promisorios como producto de exportación en Colombia y ha venido ganando participación en los mercados gracias a sus propiedades condimentarias, medicinales y fácil trasformación agroindustrial (Corporación Colombia Internacional, 2006). De acuerdo con datos de la FAO, (2013) la producción mundial de ají y pimiento fresco va en aumento en los últimos años, pasando de 26 700 847 toneladas en el 2006 a producir 36 092 631 toneladas en el 2017. El continente con mayor producción de ají es Asia, en donde destacan países como China e Indonesia mientras que en el continente americano el mayor productor es México seguido por Estados Unidos. En Colombia, en el año 2017 se produjeron 22 030 toneladas de ají, siendo el departamento de Bolívar el mayor productor aportando el 21 % de la producción nacional seguido por el Valle del Cauca con un 19 % (MADR - Agronet, 2019). El ají posee un gran potencial económico, siendo actualmente uno de los cultivos más promisorios como producto de exportación principalmente a países como Estados Unidos, el cual es el mayor importador de ají, tanto seco como verde. Aunque el ají ha venido ganando fuerza en la apuesta exportadora agropecuaria, por su tendencia histórica de exportación en el país y por la creciente demanda internacional del mismo, las estadísticas de la FAO muestran que la producción de ají en Colombia disminuyó drásticamente a partir del año 2006, pasando de una producción de 49 500 toneladas a 17 853 toneladas en 2017 (FAO, 2013).
Una de las principales limitantes en la producción de ají son las enfermedades de tipo viral, causadas comúnmente por virus pertenecientes a los géneros: Potyvirus, Cucumovirus, Tospovirus, Crinivirus y Begomovirus (Kenyon et al., 2014). Los Begomovirus (Familia Geminiviridae) presentan un genoma de ADN cadena sencilla circular, son transmitidos por la mosca blanca (Bemisia tabaci) e infectan cultivos de interés económico exclusivamente pertenecientes a las dicotiledóneas (Brown et al., 2015). Los begomovirus pueden causar hasta el 20 % de pérdidas de la producción del cultivo de ají (Hernández-Espinal et al., 2018) y su alta incidencia en los últimos 30 años en las zonas tropicales y subtropicales del planeta los ha catapultado como uno de los principales grupos de virus emergentes, que limitan la producción de hortalizas. Su aparición está directamente correlacionada con el incremento en las poblaciones de su insecto vector, la mosca blanca (Roberts et al., 2004). En Latinoamérica, los begomovirus han sido encontrados limitando la producción de ají en diferentes países: en México, se reportó al virus huasteco de la vena amarilla del chile (Pepper Huasteco yellow vein virus, PHYVV), antes conocido como el virus huasteco del chile (Pepper Huasteco virus, PHV) (Torres et al., 1993), así como al virus tigre leve del chile (Pepper mild tigre virus, PepMTV) (Brown et al., 1989); por su parte en América central, el virus del mosaico dorado del chile (PepGMV) (Lotrakul et al., 2000) antes conocido como virus del chile de Texas (Texas pepper virus, TPV); mientras que en Jamaica, se encontró el virus de la hoja rizada enana del tomate (Tomato dwarf leaf curl virus, ToDfLV) (Roye et al., 1999) en Trinidad y Tobago, el virus del mosaico amarillo del tomate (Tomato yellow mosaic virus, ToYMV), ahora conocido como virus del mosaico amarillo de la papa (Potato yellow mosaic virus, PYMV) (Umaharan et al., 1998); en Cuba, el virus de la hoja rizada amarilla del tomate (Tomato yellow leaf curl virus, TYLCV) (Quiñones et al., 2001); en Perú se reportó el virus de la hoja enrollada del chile (PepLRV) (Martínez et al., 2013); y en isla Guadalupe se reportó que Capsicum annuum también puede ser hospedero de PYMV (Urbino et al., 2004).
Para el 2017 el Valle del Cauca fue el segundo departamento productor de ají en Colombia, con una producción de 4 221 toneladas distribuidas en 193 hectáreas cosechadas, aportando un 19 % a la producción total del país (MADR - Agronet, 2019). La producción de ají en este departamento se ha visto afectado por enfermedades de tipo viral, de las cuales hasta el momento se desconoce la identidad molecular de muchas de ellas. En particular en Colombia a la fecha no se cuentan con estudios detallados sobre las enfermedades causadas por begomovirus en plantas pertenecientes al género Capsicum. Con base en estos hechos, el objetivo de esta investigación fue identificar la presencia de begomovirus en cultivos de Capsicum spp. en el Valle del Cauca empleando estrategias moleculares.
MATERIALES Y MÉTODOS
Colecta del material vegetal
Se colectaron muestras de plantas de ají tabasco (Capsicum frutescens), ají habanero (Capsicum chinense), ají cayena (Capsicum annuum var. acuminatum) y ají jalapeño (Capsicum annuum) provenientes de cultivos comerciales a cielo abierto ubicados en los municipios de Toro, Vijes, La Unión, Zarzal, Guacari, Roldanillo, Bolívar, Candelaria, Tuluá y Palmira, en el Valle del Cauca, en los años 2016 y 2018 (Tabla 1). Se tomaron hojas jóvenes con síntomas de enfermedad viral: mosaicos, corrugamiento o rugosidad de la hoja, clorosis generalizada, acortamiento de los entrenudos, deformación y reducción del limbo foliar, y deformación de frutos, así como también se colectaron muestras de hojas en plantas asintomáticas. Las hojas fueron recolectadas al azar alrededor de todo el cultivo. El material se almacenó en bolsas de papel debidamente etiquetadas y colocadas dentro de neveras de icopor con hielo seco donde se mantuvieron hasta ser llevadas al Laboratorio de Sanidad y Microbiología agrícola de la Universidad Nacional de Colombia sede Palmira para su posterior análisis. Las plantas fueron fotografiadas y los sitios de colecta fueron georreferenciados con el equipo GPS-CSX-60 (Garmin®).
Extracción de DNA genómico
Para llevar a cabo los análisis moleculares, se realizó una extracción de ADN genómico total de cada muestra vegetal. El material vegetal se molió con nitrógeno líquido usando un molino eléctrico de acero inoxidable (B. E. Classics®). Se realizó la extracción de ADN genómico total utilizando la metodología reportada por Doyle y Doyle, (1987) con la siguiente modificación: se adicionó 0,1 volumen de SDS 20 % al buffer de lisis. Posteriormente para verificar la calidad y cantidad del ADN genómico total extraído, se realizó una electroforesis en gel de agarosa al 0,8 % (p/v), teñido con bromuro de etidio (10 ng/μl). Finalmente, el gel se visualizó y fotografió en un transiluminador (Molecular Imager Gel DocXR+ Systems BIORAD®) utilizando el software Quantity-One® 4.6.5 provisto por el fabricante del equipo.
Detección de begomovirus mediante reacción en cadena de la polimerasa (PCR)
La detección molecular de begomovirus se llevó a cabo mediante la técnica de PCR empleando un juego de iniciadores universales MP16 y MP82 (Umaharan et al., 1998) que amplifican un fragmento de 0,4 - 0,5 kb del componente genómico A geminiviral. Este fragmento amplificado abarca la región intergénica así como parte del marco de lectura abierto de la proteína de la cápside (CP). La reacción de PCR se llevó a cabo en un volumen total de 25 ul con los siguientes componentes: 1,0 μM de cada iniciador, 0,2 mM de una mezcla de dNTPs, buffer 1X, 1,25 U Taq polimerasa (Fermentas), 500 ng DNA genómico total y agua Milli-Q hasta completar el volumen final deseado. Para la amplificación se usó un termociclador BioRad® (Modelo C1000) siguiendo las condiciones de amplificación reportadas por Umaharan et al., (1998). Como control positivo se utilizó ADN plasmídico que porta el componente genómico A de PYMV, un begomovirus bipartita endémico que afecta tomate en Colombia (Vaca-Vaca et al., 2012); mientras que como control negativo se usó agua.
Para la caracterización molecular del begomovirus se utilizó un segundo juego de iniciadores universales de begomovirus, RepDGRN y YMAC-N ( Mauricio-Castillo et. al., 2007) que amplifican un fragmento de 1,4 kb del componente genómico ADN-A y que abarca la región que porta los genes AL1 (Rep), región intergénica y AR1 (CP). La reacción de PCR se llevó a cabo en un volumen total de 25 ul siguiendo las condiciones de amplificación descritas por el autor.
Clonación y secuenciación
Los fragmentos 1,4 kb amplificados por PCR fueron ligados en el vector pGem T-Easy (Promega®) y transformados en células competentes de E. coli dh5a Rec(-) por choque térmico (Sambrook y Russell, 2001). Se realizó extracción de ADN plasmídico de las células transformantes empleando el kit QIAprep Spin Miniprep Kit (Qiagen®) y se verificó la presencia del inserto mediante análisis de restricción con la enzima EcoRI. El ADN plasmídico de las clonas que portaban inserto fue enviado a secuenciar en Macrogen (Seúl, Corea del Sur).
Análisis bioinformático
Para cada fragmento viral se realizó el ensamble de las lecturas de secuenciación empleando el software CLC Main workbench v7.0 (Qiagen®). Con el fin de conocer la identidad de las secuencias de nucleótidos, se realizó una comparación usando el algoritmo Blastn contra la base de datos "refseq_genomic" de GenBank (Altschul et al., 1997). De acuerdo con este análisis, se seleccionaron ocho secuencias de begomovirus que presentaban mayor identidad con las secuencias del virus aislado en cultivos de ají de Palmira y Vijes (Tabla 2). Y se adicionaron al análisis, cuatro secuencias de begomovirus aislados de ají en México y Brasil; y un begomovirus aislado de yuca (Manihot esculenta L.) en África, para incluirlo como secuencia externa del análisis. Todas las secuencias de nucleótidos de los begomovirus arriba mencionadas se obtuvieron del GenBank y se editaron en el programa Bioedit para realizar un alineamiento múltiple en el programa Clustal W, junto con la secuencia de nucleótidos de los begomovirus aislados de ají de Palmira y Vijes (Larkin et al., 2007). A partir del alineamiento múltiple se calcularon los valores de identidad entre estas secuencias a nivel de nucleótidos empleando el software Sequence Demarcation Tool SDT 1.2® (Muhire et al., 2014).
Tabla 2 Porcentaje de identidad de secuencia de nucleótidos del fragmento de 1,4 kb del componente genómico de DNA-A de begomovirus aislados de Capsicum spp. de Palmira y Vijes con otros begomovirus, utilizando el software SDT v1.2.
| Virus /Número de accesión en GenBank | Acrónimo | Hospedero | País | % identidad nt) | ||
|---|---|---|---|---|---|---|
| Familia | Especie | Vijes | Palmira | |||
| Passionfruit leaf distortion virus / NC_031764 | PLDV | Pasifloraceae | Passiflora edulis | Colombia | 87,2 | 86,6 |
| Bean dwarf mosaic virus / NC_001931 | BDMV | Fabaceae | Phaseolus vulgaris | Colombia | 86,9 | 84,9 |
| Corchorus yellow spot virus / NC_008492 | CoYVYuV | Malvaceae | Corchorus siliquosus | México | 85,3 | 84,6 |
| Bean white chlorosis mosaic virus / NC_022005 | BChMV | Fabaceae | Phaseolus vulgaris | Venezuela | 84,5 | 84,9 |
| Sida yellow vein virus / NC_004661 | SiYVV | Malvaceae | Sida rhombifolia | Honduras | 83,8 | 83,4 |
| Sida yellow mottle virus strain Cuba / NC_016082 | SiYMoV | Malvaceae | Sida rhombifolia | Cuba | 83,3 | 84,7 |
| Sida golden mosaic Honduras virus / NC_004659 | SiGMHV | Malvaceae | Sida rhombifolia | Honduras | 83,3 | 83,8 |
| Tomato severe rugose virus / NC_009607 | ToSRV | Solanaceae | Capsicum sp | Brasil | 78,3 | 78,5 |
| Tomato golden vein virus / NC_038807 | TGVV | Solanaceae | Solanum lycopersicum | Brasil | 77,5 | 76,9 |
| Pepper leafroll virus isolate / KC769819 | PepLRV | Solanaceae | Capsicum baccatum var. pendulum | Perú | 76,9 | 76,7 |
| Pepper huasteco yellow vein virus / NC_001359 | PHYVV | Solanaceae | Capsicum sp | México | 76,4 | 76,5 |
| Pepper golden mosaic virus / NC_004101 | PepGMV | Solanaceae | Capsicum sp | México | 72,1 | 71,2 |
| African cassava mosaic virus / NC_001467 | ACMV | Euphorbiaceae | Manihot esculenta | Kenya | 68,3 | 66,7 |
Los análisis filogenéticos para los virus identificados se realizaron con el alineamiento múltiple de las secuencias seleccionadas en el programa Mega 7 (Kumar et al., 2016). La historia evolucionaria se infirió usando el método de Neighbor-joining (Saitou y Nei, 1987) y las distancias evolutivas se obtuvieron usando el método de máxima probabilidad (Tamura et al., 2004) con 1 000 bootstrap.
RESULTADOS Y DISCUSIÓN
Detección por primera vez de begomovirus en muestras de ají colectadas en el Valle del Cauca
Un total de 197 muestras de cuatro variedades de ají fueron colectadas en los municipios de Toro, Vijes, La Unión, Zarzal, Guacari, Roldanillo, Bolívar, Candelaria, Tuluá y Palmira, localizados en el Valle del Cauca: 34 de ají cayena (Capsicum annuum var. acuminatum), 63 de ají habanero (Capsicum chinense), 94 de ají tabasco (Capsicum frutescens) y seis de ají jalapeño (Capsicum anuum). Las plantas de la variedad cayena y jalapeño en su mayoría se presentaron asintomáticas o con síntomas muy leves, como se observa en la Fig. 1c y d; es decir, no se apreciaron manifestaciones visibles de la interacción entre estas y el virus. Mientras que las variedades tabasco y habanero mostraron sintomatología característica de enfermedad viral, en donde predominaba la presencia de mosaicos y deformación de la lámina foliar (rugosidad) (Fig. 1a y b). También, algunas plantas presentaban clorosis intervenal y generalizada, acortamiento de los entrenudos, deformación de frutos y enanismo.

Figura 1 Variedades de ají (Capsicum spp.) colectadas en campo y síntomas observados en la mayoría de las muestras positivas a begomovirus. a., ají habanero (Capsicum chínense) con síntomas como deformación rugosa de la lámina foliar y mosaicos; b., ají tabasco (Capsicum frutescens) con deformación rugosa de la lámina foliar e islas verdes; c., ají cayena (Capsicum annuum var. acucminatum) asintomática; d., ají jalapeño (Capsicum annuum) asintomática.
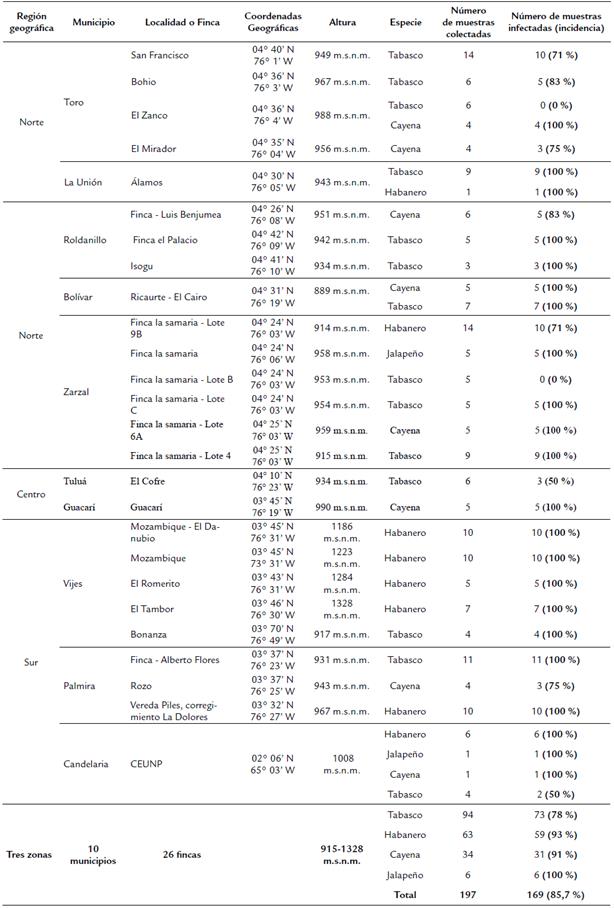
Mediante la técnica de PCR se detectaron como positivas para infección por begomovirus el 85,7 % (169/197) de las muestras colectadas en campo (Tabla 1). Estas muestras amplificaron un fragmento de aproximadamente 0,4 kb del componente genómico A geminiviral (Fig. 2), el cual presenta un tamaño diferente con respecto al control positivo PYMV (ver asterisco). Este resultado indica que el virus detectado en ají es diferente a PYMV, un begomovirus que afecta tomate en Colombia. Esto es interesante, ya que muchos de los begomovirus que infectan tomate se han reportado que afectan ají; por ejemplo, en Trinidad y Tobago, y en la Isla Guadalupe se ha reportado que PYMV afecta Capsicum (Umaharan et al., 1998; Urbino et al., 2004); en Cuba, TYLCV se ha detectado en cultivos de ají y frijol (Quiñones et al., 2001; Martínez et al., 2002); en Perú se reportó que PepLRV infecta cultivos de ají, frijol y tomate (Martínez et al., 2013); y en México, se ha reportado que PHYVV y PepGMV afectan tomate y ají (Torres et al., 1993; Lotrakul et al., 2000).
Figura 2 Detección molecular de begomovirus en plantas de ají, con el empleo de PCR según la metodología propuesta por Umaharan et al., (1998). M, Marcador de peso molecular GeneRuler® 1kb DNA Ladder (Thermo Scientific®); 1-11, muestras colectadas en el corregimiento de Rozo; 12-21, muestras colectadas en el corregimiento de Mozambique-Vijes; 22-28, Muestras colectadas en la vereda el Tambor-Vijes; 29-38, muestras colectadas en el corregimiento de Mozambique-Vijes; C+: Control positivo (begomovirus PYMV); C-: Control negativo (agua). El asterisco indica el tamaño del fragmento amplificado de aproximadamente 0,4 kb.
En la Tabla 1, se observa que la mayor cantidad de muestras positivas para begomovirus se presentó en la zona Centro-Sur del Valle del Cauca, representada por los municipios de Tuluá, Guacarí, Vijes, Palmira y Candelaria; en donde se colectaron 84 muestras de ají (habanero, 48; tabasco, 25; cayena, diez; jalapeño, uno), y de las cuales 78 fueron positivas con un porcentaje de incidencia de 92,8 %. La zona norte, representada por los municipios de Toro, La Unión, Zarzal, Roldanilllo y Bolívar presentó un porcentaje de incidencia menor, 80,5 % (91/113). La variedad con mayor porcentaje de incidencia fue jalapeño, donde el 100 % de las muestras resultaron positivas para este virus. La variedad habanera presentó un 93,6 % de incidencia, cayena un 91,1 % y por último, tabasco con un 77,6 %. Aunque la mayoría de las muestras de la variedad cayena y jalapeño se mostraron asintomáticas, tras los análisis moleculares, un alto porcentaje resultaron positivas para begomovirus (Tabla 1). Esto podría indicar que estas variedades estarían funcionando como hospedero asintomático de este virus y fuente de inoculo para otras variedades de ají.
Con base en la literatura reportada a la fecha y en los resultados de esta investigación, este sería el primer reporte en Colombia de un begomovirus infectando cultivos comerciales de Capsicum spp. a lo largo y ancho del Valle del Cauca. Basados en el gran porcentaje de muestras de ají que resultaron positivas para begomovirus (85,7 %), se puede deducir que estos virus se encuentran distribuidos biogeográficamente afectando esta solanácea y muy probablemente hacen presencia en otras regiones del país debido al gran movimiento e intercambio de material vegetal entre productores a nivel país.
En el Valle del Cauca, los reportes de begomovirus afectando cultivos de interés agronómico, no son nuevos. Desde 1975, se han reportado infecciones begomovirales en cultivos de fríjol, donde fueron detectados los virus del mosaico dorado y del moteado clorótico del frijol (Gálvez et al., 1975). En 1990, Morales et al., (1990) caracterizaron parcialmente el virus del mosaico enano del fríjol (BDMV) en muestras de fríjol. Posteriormente, Vaca-Vaca et al., (2012) identificaron un begomovirus, PYMV, como el virus que predomina y afecta los cultivos de tomate en las principales zonas productoras de esta solanácea en Colombia (Valle, Región cafetera, Cundinamarca, Santander y Antioquia). Y recientemente, Vaca-Vaca et al., (2017) reportaron el virus de la distorsión de la hoja de maracuyá (Passionfruit leaf distortion virus, PLDV), un nuevo begomovirus infectando maracuyá en el Valle del Cauca.
Los resultados de la presente investigación sugieren la presencia de un geminivirus no descrito en la literatura, que se adicionaría a los previamente descritos en América infectando al cultivo del ají. Por ejemplo, Torres et al., (1993) reportaron el virus PHYVV en la región Huasteca de México infectando C. annuum, variedad Serrano. Umaharan et al., (1998) en Trinidad y Tobago identificó los virus PHYVV y PYMV en C. annuum y C. frutescens.Lotrakul et al., (2000) aisló PepGMV de ají tabasco (C. frutescens) y de Capsicum chinense variedad habanero en Costa Rica. También se reportó que el virus de la distorsión de la hoja clorótica del tomate (Tomato chlorotic leaf distortion virus, TCLDV) aislado de tomate en Venezuela puede infectar plantas de C. chinense (Zambrano et al., 2011). En ensayos por biobalistica en Venezuela se inoculo el virus del mosaico de Euphorbia de Venezuela (Euphorbia mosaic Venezuela virus, EuMVV) en C. annuum y se observó amarillamiento y distorsión de la hoja (Zambrano et al., 2012). Además, en Perú en otras especies, como C. bacatum en Cv. Pendulum, se detectó y aisló PepLRV de chile amarillo peruano (Martínez et al., 2013). Con base en lo expuesto anteriormente se establece que los begomovirus figuran entre las principales amenazas fitopatológicas para el cultivo de ají en este hemisferio.
En el caso concreto del Valle del Cauca los resultados de esta investigación demuestran que la mayoría de los materiales de ají sembrados son susceptibles al ataque de los begomovirus. Se podría de alguna manera exceptuar las variedades cayena y jalapeño, las cuales, aunque fueron fenotípicamente asintomáticas, resultaron positivas a begomovirus en los análisis moleculares. Este resultado podría explicarse si tenemos en cuenta que, posiblemente los begomovirus presentes en cayena y jalapeño, hasta el momento se están adaptando a estos hospederos, siendo probablemente el factor movimiento y dispersión vía floema, el principal cuello de botella que les ha impedido alcanzar otros tejidos vegetales y generar allí la sintomatología geminiviral típica, tal y como lo demuestran los estudios de Hipper et al., (2013). Por lo cual estas variedades podrían ser una opción temporal para los productores de la zona, mientras se desarrollan materiales tolerantes o resistentes a este nuevo begomovirus en las variedades habanero y tabasco.
Caracterización molecular de begomovirus detectados en ají en Palmira y Víjes
Para conocer la identidad de los begomovirus detectados en muestras de ají habanero (C. chinense) colectadas en Palmira y Vijes, municipios donde la presencia de begomovirus fue alta (100 % de infección por begomovirus), fragmentos virales de 1,4 kb fueron clonados. Estos fragmentos virales portan una parte de los genes que codifican para las proteínas Rep y CP, e incluye toda la región intergénica. Se obtuvieron cinco clonas independientes a partir de muestras de ají colectadas en Palmira y el análisis de la secuencia de nucleótidos de estas mostró que todas eran el mismo virus (99-100 % identidad). De manera similar, la secuencia de ADN de seis clonas independientes obtenidas de muestras de ají colectadas en Vijes, fueron idénticas entre nucleótidos (99-100 % identidad). Por lo tanto, se obtuvieron dos secuencias de nucleótidos de 1429 y 1425 pb cada uno, correspondientes a un begomovirus identificados en ají habanero colectado en Vijes y Palmira, respetivamente. Los fragmentos virales mostraron un 91,6 % de identidad entre ellos, por lo que se pueden considerar aislados de una misma especie, ya que el porcentaje de identidad está entre el límite entre especies (< 91 %) y aislados (< 94 %) (Brown et al., 2015). Estas secuencias fueron depositadas en la base de datos de GenBank con los siguientes números de accesión: MG888725, para el begomovirus de Piles, Palmira y MG888726, para el begomovirus de Vijes. El análisis de secuencia de nucleótidos en el software SDT (Tabla 2), mostró que los begomovirus aislados en Vijes y Palmira comparten la identidad más alta (87 % y 86 %) con el virus de la distorsión de la hoja de maracuyá (PLDV, número de accesión NC_031764), un begomovirus aislado de maracuyá (Passiflora edulis) en Valle del Cauca (Vaca-Vaca et al., 2017); así como un bajo porcentaje de identidad (76,9 % y 76,7 %) con el virus de la hoja enrollada del chile (PepLRV, KC769819), un begomovirus detectado afectando el cultivo de ají (Capsicum baccatum var. Pendulumen) en Perú (Martínez et al., 2013). Con base al criterio de demarcación actual para las especies de Begomovirus establecidas por el Comité Internacional de Taxonomía de Virus (ICTV) en 2017 (< 91 % a nivel de nucleótidos del genoma del virus) (Brown et al., 2015), se plantea la hipótesis de que los begomovirus aislados de ají en este trabajo de investigación podrían ser una nueva especie geminiviral que está infectando este cultivo en Colombia. Para confirmar esta hipótesis, se está trabajando en la obtención del genoma completo ADN-A y ADN-B de estos begomovirus.
Los análisis filogenéticos (Fig. 3) muestran que los begomovirus aislados de Vijes y Palmira se agrupan en un mismo clado, pero separados de PLDV y BDMV, dos begomovirus que afectan maracuyá y frijol en Colombia, con los cuales presentó un mayor acercamiento, pero con una identidad no muy cercana, pues es menor al 91 %. Asimismo, los begomovirus aislados de ají se agrupan también con virus aislados de arvenses como Sida rhombifolia y Corchorus siliquosus, pero no con otros virus aislados de ají en el continente americano. Este resultado coincide con los obtenidos a partir de la comparación de pares de secuencias de nucleótidos (Tabla 2), en donde begomovirus aislados de ají en Perú y México mostraron baja identidad y se localizan en grupos separados del grupo de begomovirus de Colombia. Una posible explicación para esto es la existencia de dos grandes barreras ecológicas, una hacia el norte en lo que se conoce como el tapón del Darien, es decir, la selva tropical entre Panamá y Colombia; y hacia el sur, la selva húmeda tropical del Amazonas, barrera que separa al país del Perú, Ecuador y Brasil. Estas dos barreras ecológicas generaron para los begomovirus de Colombia un escenario único para evolucionar y especiarse bajo condiciones de selección diferentes a las que pudieron estar sometidos los geminivirus que afectan ají en Perú y México. Sustentan esta afirmación los resultados previamente reportados por Vaca-Vaca et al., (2017), quienes caracterizaron un begomovirus nuevo aislado de maracuyá (PLDV) en Valle del Cauca, el cual se agrupó en un clado independiente del begomovirus que infecta este cultivo en Brasil.
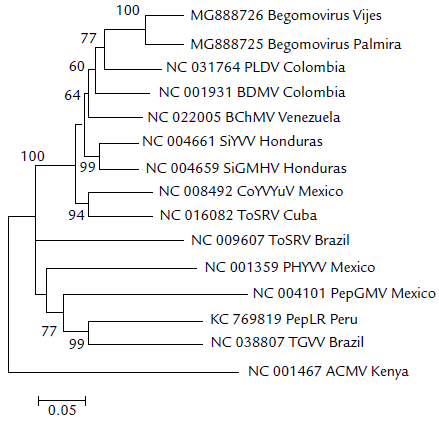
Figura 3 Análisis filogenético de los begomovirus aislados de ají en Palmira y Vijes. Árbol filogenético construido con el método Neighbor-joining usando un fragmento de 1,4 kb del genoma A que porta la región 5'de los genes AC1 (Rep) y proteína de la cápside (CP), y la región intergénica empleando 1 000 bootstrap. Para los nombres de las secuencias de begomovirus usados en el análisis ver Tabla 2. El virus del mosaico de yuca de África (Africa cassava mosaic virus -ACMV) se utilizó como grupo externo.
La alta distribución y diseminación de este begomovirus en cultivos de ají en el Valle del Cauca, podría ser explicada por fenómenos como el aumento de poblaciones de B. tabaci biotipo B (Roberts et al., 2004), el cual tiende a tener un mayor potencial biótico y desplazar al biotipo A. Esto fue demostrado en estudios en la costa norte del país (Quintero et al., 2001) y en el Valle del Cauca donde el biotipo B desplazo no solamente al biotipo A, sino también a Trialeurodes vaporariorum, especie que por muchos años fue la más predominante en estos departamentos (Rodríguez et al., 2005; Rodríguez et al., 2012). Otras posibles explicaciones son también la expansión e intensificación de los sistemas de producción, la reducción de la variación genética en las especies cultivadas y el uso generalizado e indiscriminado de insecticidas en la agricultura. En consecuencia, B. tabaci y los virus que transmite ya no están restringidos a hábitats nativos ni a límites geográficos naturales (Brown, 2010).
Estos factores podrían favorecer una mayor presión de selección en el vector B. tabaci, donde se pueden generar especies cripticas adaptadas a múltiples hospederos como se ha encontrado en Cuba (Martínez et al., 2016). Basados en los hechos hasta aquí expuestos, bien se podría pensar que los begomovirus podrían colonizar nuevos cultivos de interés agronómico en Colombia y llegar a nuevas regiones geográficas donde previamente no se habían reportado.
Adicionalmente, las arvenses o plantas acompañantes, podrían tener un papel muy importante en la aparición y distribución de los begomovirus aislados de los cultivos de ají en el Valle del Cauca. Esto debido a que las arvenses pueden actuar como reservorios temporales de begomovirus así como de ser fuente de inoculo primario para la diseminación de begomovirus a través de la mosca blanca, desde las propias arvenses hacia las plantas cultivadas de interés económico (Ambrozevicius et al., 2002; López-López et al., 2012). En diversos estudios se ha identificado que arvenses tales como Desmodium uncinatum, Macroptilium lathyroides, Rhynchosia minima, Malva sp., Malvastrum sp., Sida rhombifolia, Eragrostis tenella, Ehretia tpunifolia, Megathopa villosa, y Pavonia sp. actúan como fuente de begomovirus que infectan frijol común (Morales et al., 2000). López-López et al., (2012; 2014) y Vaca-Vaca et al., (2018) identificaron arvenses asociados al cultivo de tomate en Valle del Cauca que eran hospederos de begomovirus: Desmodium sp., Amaranthus dibius, Laportea aestuans, Rivina humilis, Lantana camara, hybanthus atenuattus, Vervena sp., Crotón hirtus, Petiveria alliace, Amaranthus dubius, Rhynchosia minima, Rivina humilis, Desmodium sp., Caesalpinia sp. y Plumbago sp. Razón por la cual, podríamos decir que cabe la posibilidad de que estos virus aislados de cultivos de ají, pudieron haber estado latentes en las arvenses; y con la llegada y adaptación de B. tabaci biotipo B a nuevos agro ecosistemas, se diseminaron a cultivos de interés agronómico como tomate, frijol, maracuyá y ahora a cultivos de ají en el Valle del Cauca. Todos los resultados hasta aquí expuestos apoyan la hipótesis de que el virus aislado de ají en Valle del Cauca podría ser una nueva especie begomoviral. Actualmente, se está trabajando en la obtención del genoma completo de este virus para utilizarlo como fuente de inoculo para evaluar colecciones de ají en la búsqueda de un cultivar de ají resistente a este nuevo begomovirus.
CONCLUSIONES
Se detectó la presencia de begomovirus en cultivos de ají de las variedades cayena, habanero, tabasco y jalapeño, ubicados en diez municipios del Valle del Cauca (Colombia) productores de esta solanácea, con un porcentaje de infección por begomovirus del 85,7 %. Esto se constituye en el primer reporte de un begomovirus infectando cultivos de ají en Valle del Cauca y Colombia.
Se caracterizaron a nivel molecular dos fragmentos begomovirales de 1,4 kb aislados de ají habanero colectado en Palmira y Vijes (Valle del Cauca), respectivamente. Los análisis bioinformáticos indicaron que estos begomovirus eran diferentes a otros previamente reportados que limitan la producción de ají en el mundo; y que están más relacionados con virus endémicos del Valle del Cauca, PLDV y BDMV, virus que infectan maracuyá y frijol, respectivamente. Estos análisis estarían indicando que este begomovirus que infecta cultivos de ají podría ser una nueva especie que no ha sido reportada en el mundo.
Los resultados de este trabajo de investigación sirven de materia prima esencial para implementar estrategias de manejo integrado que permitan reducir la infección y diseminación de este begomovirus en cultivos de ají en el departamento, y el país.


