











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Se ha demostrado que hay relación entre la gemación de algunos virus y la vía de clasificación endosomal. La primera evidencia vino de la interacción descrita entre la proteína Gag del virus de la inmunodeficiencia humana (VIH) y la proteína celular Tsg101 (Garrus et al., 2001; Martin-Serrano et al., 2001), la cual, entre otras funciones, hace parte del complejo de clasificación endosomal conocido como ESCRT (por sus siglas en inglés: Endosomal Sorting Complexes Required for Transport). Después se demostró que la sobreexpresión de algunos mutantes de Tsg101 inhibía la propagación del virus sin causar efectos adversos aparentes sobre ESCRT y ese resultado planteó la posibilidad de desarrollar agentes antivirales dirigidos a interferir la interacción Tsg101/Gag (Goila-Gaur et al., 2003). Aunque el VIH fue el primer virus para el que se describió la participación de proteínas de la vía endosomal en los procesos de ensamblaje y gemación viral (Garrus et al., 2001; Martin-Serrano et al., 2001; Bache et al., 2003; Katzmann et al., 2003; Martin-Serrano et al., 2003; Janvier et al., 2011), para el año 2013 ya se había descrito interacciones entre virus y proteínas ESCRT para retrovirus, arenavirus, paramixovirus, reovirus, picornavirus, filovirus, rabdovirus, herpesvirus y hepadnavirus (Votteler y Sundquist, 2013).
Específicamente para flavivirus, hay reportes que relacionan procesos de ensamblaje o liberación con proteínas de la maquinaria ESCRT; es el caso del virus de la encefalitis japonesa (JEV) y Tsg101 (Chiou et al., 2003; Tabata et al., 2016) y el virus de la fiebre amarilla (YFV) y Alix (Carpp et al., 2011). Aunque también se demostró que dominantes negativos de Alix afectaban levemente la producción de partículas virales del West Nile Virus (WNV), el "knock-down" de Alix con ARN de interferencia (iRNA) no produjo efecto alguno sobre la liberación del virus y eso sugirió que WNV no se propaga usando la maquinaria ESCRT (Garg et al., 2013).
Para el virus del dengue (DENV), otro flavivirus, el análisis del proteoma de células endoteliales humanas infectadas con DENV reveló alteración en los niveles de traducción de 35 proteínas, entre las que Alix se encontró sobre-expresada. Este resultado llevó a proponer un papel fundamental de Alix durante la replicación de DENV y la posibilidad de convertir a esta proteína en un blanco terapéutico para reducir el nivel de replicación de DENV "in vivo" (Pattanakitsakul et al., 2010). Hoy es claro que además de participar en la formación de las vesículas intraluminales, la citocinesis, el tráfico de receptores de señalización celular y la apoptosis, Alix tiene un papel importante en el ensamblaje y gemación de varios virus (Bissig et al., 2014).
Luego, en un trabajo dirigido a identificar interacciones entre proteína humanas y proteínas de DENV, se encontraron 139 interacciones que involucraban 105 proteínas humanas, entre las cuales estaba Alix (Khadka et al., 2011). Finalmente, se reportó que el "knock-down" para Alix usando siRNAs o el sistema CRISPR-Cas9 tenía gran efecto en la producción de DENV en células HEK 293T/17 (Thepparit et al., 2019). Sin embargo, este último resultado entró en contradicción con un trabajo previo que demostró que disminuir la expresión de Alix usando siRNAs en células 293A, no tenía efecto alguno sobre la liberación de DENV (Tabata et al., 2016). Ligado a esta controversia sobre la participación de Alix en la propagación de DENV, surgió también el interrogante sobre cual o cuales proteínas virales podrían interactuar con ella.
El genoma del DENV codifica una poliproteína precursora de tres proteínas estructurales (Cápside, Pre-proteína M y Envoltura) y siete no estructurales (NS1, NS2A, NS2B, NS3, NS4A, NS4B y NS5) (Deubel et al., 1986; Mackow et al., 1987; Deubel et al., 1988), que en todos los flavivirus se ubican de forma similar a lo largo del genoma. En el caso de NS3, se demostró que su interacción con Alix contribuye a la liberación de las partículas infecciosas de YFV (Carpp et al., 2011), mientras que su asociación con Tsg101 parece ser determinante para el ensamblaje del virus JEV (Chiou et al., 2003). Específicamente para DENV, se encontró interacción entre NS3 y Alix (Khadka et al., 2011), y más tarde entre NS3 y Tsg101 (Tabata et al., 2016). NS3 es una proteína multifuncional con actividades proteasa (Falgout et al., 1991), ARN helicasa, NTPasa (nucleósido 5'-trifosfatasa) (Li et al., 1999) y RTPasa (ARN 5'-trifosfatasa) (Bartelma et al., 2002). Adicionalmente, NS3 es una proteína inmuno-dominante y por tanto uno de los blancos más significativos de la respuesta inmune durante la infección con DENV (Duangchinda et al., 2010).
El objetivo de este trabajo fue explorar la participación de la proteína Alix en los procesos de transcripción, traducción, ensamblaje y liberación de DENV, así como su interacción con la proteína NS3.
MATERIALES Y MÉTODOS
Ensayos de interferencia.
Cultivo Celular.
Células A549 (ATCC® CCL-185™) fueron cultivadas en medio RPMI (Sigma Aldrich) suplementado con suero fetal bovino (SFB, Hyclone) al 10 % y mantenidas a 37 °C, en una atmosfera de 5 % de CO2. Al alcanzar una confluencia del 90 %, las células fueron disociadas enzimáticamente con una solución de tripsina 0,3 % (Sigma Aldrich) y EDTA 0,5 mM (Sigma Aldrich) por 5 minutos a 37 °C. Luego, dependiendo del ensayo a realizar, las células se sembraron nuevamente en laminillas pre-tratadas con Poli-L-lisina 10 Mg/ml (Sigma Aldrich) o en placas de 24 o 96 pozos.
Infección con DENV2.
El virus dengue serotipo 2 (DENV2) usado fue el pasaje 6 de un aislamiento colombiano adaptado a células C6/36 (INS-2009) (Morales etal., 2017). Células A549 fueron incubadas con una suspensión de DENV2 (en medio RPMI suplementado con FBS al2%)a una multiplicidad de infección de 1 (MOI:1) por 1 hora. Luego, el inóculo viral fue retirado, las células se mantuvieron en cultivo por 48 horas y la infección fue confirmada por inmunofluorescencia indirecta (IF). Las células fueron fijadas sobre laminillas con paraformaldehído (PFA, Carlo Erba) al 4 % en buffer fosfato salino (PBS), tratadas con una solución 50 mM de cloruro de amonio (Sigma Aldrich), permeabilizadas con 0,3 % Tritón X-100 (Sigma Aldrich) y bloqueadas con suero de cabra (Gibco) al 10 % en PBS. Luego, las células fueron incubadas por 1 hora a 37 °C con un anticuerpo monoclonal dirigido contra la proteína de envoltura (E) del virus de la encefalitis de San Luis (1:500) (Millipore. MAB8744), el cual tiene reacción cruzada con la proteína E del DENV2. A continuación, las células fueron lavadas e incubadas con un anticuerpo anti-ratón conjugado con biotina (1:200) (Vector. BA9200) y posteriormente con una solución de estreptavidina acoplada a Alexa 594® (Molecular Probes). Finalmente, se contrastaron los núcleos con Hoechst 33342 (Sigma Aldrich), las laminillas fueron montadas con vectashield (Vector) y se observaron bajo microscopio (Zeiss Axioimager A2) utilizando un sistema de fluorescencia X-Cite serie 120Q. Con la herramienta cell counter (Image J NIH) se contaron células totales y células infectadas sobre las imágenes para determinar los porcentajes de infección. Como control se evaluaron células tratadas con el mock (sobrenadante de células C6/36 no infectadas).
Tratamiento con siRNA.
En un trabajo previo en el que se evaluó la participación de proteínas ESCRT en la producción de DENV2, encontramos que la proteína Vps4B no participaba en los procesos de ensamblaje y liberación viral (Bueno, 2015). Por eso, en este estudio se usó a Vps4B como control. Así, se utilizó un "pool" de cuatro interferentes de Alix (M-004233-02-0010) y un "pool" de cuatro interferentes de Vps4B (M-01319-02-0010) (siRNA siGENOME SMARTpools) (GE Healthcare Dharmacon). Según el fabricante, estas mezclas de interferentes garantizan entre 75 y 80 % de disminución en la expresión.
Para el experimento se preparó una solución de OligofectaminaTM Reagent (Invitrogen) en medio RPMI y se incubó por 15 minutos a temperatura ambiente (T° amb) en placas de 24 pozos. A continuación, los siRNA fueron adicionados a una concentración final de 100 nM y se incubaron por 15 minutos a T° amb. En paralelo, células A549 infectadas con DENV2 se disociaron enzimáticamente, se adicionaron sobre la mezcla anterior (2 x 105 células) y se incubaron por 4 horas a 37 °C y 5 % de CO2. Al cumplir el tiempo, se adicionó medio RPMI con SFB al 2 % hasta completar un volumen final de 500 μ! en cada pozo, y se incubó nuevamente la placa por 48 horas. Como control se utilizaron células A549 no infectadas y no interferidas, células infectadas y no interferidas, y células infectadas e interferidas con un siRNA no relacionado (D-001206-13-20) (GE Healthcare Dharmacon), de aquí en adelante siRNA-NR. Este experimento se hizo por triplicado (tres cultivos independientes) y cada condición por duplicado. A las 48 horas de incubación post interferencia, se evaluó la viabilidad de las células A549 infectadas e interferidas y sus controles, utilizando sales de MTT® (Roche).
Confirmación de la interferencia: PCR cuantitativo.
La extracción de ARN se hizo con Trizol® Reagent (Invitrogen), siguiendo las recomendaciones del fabricante. Para la cuantificación de los transcritos se utilizó el sistema SuperScript III RT/Platinum® Taq Mix One Step 2X SYBR Green Reaction Mix (Invitrogen), 200 ng de ARN y 0,1 µM de oligonucleótidos específicos para Alix (oligo sentido: TAC-CGG-CTC-TCT-GAC-CAG-TT y oligo antisentido: GAT-CCG-TGT-CGT-AAC-GTC-TG) o para Vps4B (oligo sentido: TAA-GAC-GGA-GTT-CCT-AGT-GC y oligo antisentido: CGT-GAG-ACT-GTT-CTG-AGT-GG), diseñados sobre las secuencias NM_013374.6 y NM_004869.4, respectivamente. Como gen de referencia se utilizó el gen 18SARNr. La expresión relativa se obtuvo aplicando el análisis de Schefe (Schefe et al., 2006) al hacer una relación entre la expresión de los transcritos de interés y el gen de referencia, en las células interferidas versus las interferidas con el siRNA-NR. Se usaron dos cultivos independientes y cada condición se evaluó por duplicado.
Confirmación de la interferencia: Ensayos de "Western Blot" (WB).
Células A549 infectadas e interferidas y sus controles fueron tratadas con buffer RIPA (1 % Nonidet P40, 0,5 % deoxicolato de sodio y 1 % SDS en PBS). Se adicionó luego coctel de inhibidores de proteasas (1:100) (Sigma Aldrich. P8340) y el lisado se sometió a sonicación. Los extractos fueron cuantificados por el método del ácido bicinconínico y 30 μg de cada uno fueron sometidos a SDS-PAGE (poliacrilamida 10 %). Las proteínas fueron transferidas a membranas de PVDF (Thermo Fisher Scientific) que fueron luego sumergidas en buffer de bloqueo TBST (10 mM Tris HCl pH 7,5, 150 mM NaCl, 0,1 % Tween-20) más BSA (1 %) y leche descremada (2 %). A continuación, las membranas se incubaron con el anticuerpo monoclonal anti-Alix (1:100) (Abcam. ab117600) o con el anticuerpo policlonal anti-Vps4B (1:5000) (Abcam. ab102687) durante toda la noche a 4 °C. Al día siguiente se adicionó el anticuerpo secundario correspondiente y se incubó durante 1 hora a T° amb. Se usó un anti-ratón acoplado a peroxidasa (1:1000) (Thermo-Scientific) para Alix y un anti-conejo acoplado a peroxidasa (1:2000) (Thermo-Scientific) para Vps4B. El revelado de la señal se hizo con el sistema SuperSignal™ West Pico PLUS (Thermo Scientific. 34080). Como control de carga se hizo la detección de /?-actina con un anticuerpo primario anti-actina (1:5000) (Sigma. A5060) y un anticuerpo secundario anti-conejo acoplado a peroxidasa (1:2000) (Thermo-Scientific).
Efecto de la interferencia de Alix y Vps4B en la transcripción y traducción viral.
Se tomaron células A549 interferidas para Alix o Vps4B y sus controles, y se evaluó por RT-PCR la expresión del transcrito de NS1 y por "Western Blot" (WB) la expresión de la proteína NS3. Para el experimento de RT-PCR, el ARN fue extraído como se describió antes y se hizo la síntesis de ADN complementario (ADNc) usando M-MLV retro-transcriptasa (Promega) y finalmente una PCR para los genes nsl y 18S ARNr. Los oligonucleótidos usados fueron diseñados con el programa Primer-BLAST (www.ncbi.nlm.nih.gov/tools/primer-blast/) con base en las secuencias NP_739583.2 (nsl) (sentido: TGC-AGG-CCG-ATA-GTG-GTT y antisentido: TGC-AGG-CCG-ATA-GTG-GTT) y NR_003286.2 (18S ARNr) (oligo sentido: ATA-CAT-GCC-GAC-GGG-CGC-TG y oligo antisentido: AGG-GGC-TGA-CCG-GGT-TGG-TT). La reacción de amplificación se hizo con 1,5 mM de MgCl2, 200 μM de dNTPs, 1 μM de oligonucleótidos y 0,5 U de ADN polimerasa GoTaq Flexi (Promega). El programa de amplificación comprendió una desnaturalización inicial a 94 °C por 2 minutos, 30 ciclos de 94 °C por 45 segundos, 55 °C por 45 segundos y 72 °C por 60 segundos, seguidos por una extensión final de 72 °C por 10 minutos.
Para los ensayos de WB se usó un anticuerpo anti-NS3403_599 producido previamente por el laboratorio (Morales et al., 2017). Brevemente, se incubó la membrana toda la noche a 4 °C con anti-NS3403_599 (1:1000) y al día siguiente se lavó y se incubó con el anticuerpo anti-rata acoplado a peroxidasa (1:2500) (Thermo-Scientific. 31470) durante una hora a T° amb. El revelado se hizo con peróxido de hidrogeno (0,02 %) y diaminobencidina (DAB) (0,05 %). Como control de carga se hizo la detección de β-actina con un anticuerpo primario anti-actina (1:10000) (Sigma. A5316) y un anticuerpo secundario anti-ratón conjugado con biotina (1:5000) (Vector. BA9200).
Efecto de la interferencia de Alix y Vps4B sobre el ensamblaje y la liberación viral.
Se recolectaron los sobrenadantes de cultivos de células A549 infectadas e interferidas y de los cultivos control. Luego, se adicionó RPMI sobre las monocapas y las células se lisaron por congelación/descongelación sucesiva, haciendo dos ciclos en los que la suspensión celular se congeló primero a -80 °C y se llevó luego a 4 °C hasta descongelación. Simultáneamente, células LLC-MK2 (ATCC® CCL-7™) fueron sembradas en placas de 96 pozos con medio DMEM (Dulbecco's modified Eagle's médium. Sigma Aldrich) suplementado con SFB (10 %) y fueron mantenidas por 24 horas a 37 °C en una atmosfera de 5 % de CO2. A continuación se retiró el medio y se adicionaron 50 μl de los sobrenadantes o 50 μl de los lisados de las monocapas. Las células fueron llevadas nuevamente a incubación y 48 horas después fueron fijadas y permeabilizadas tal como se describió antes en el protocolo de la IF. Luego, las células fueron incubadas con el anticuerpo primario anti-E y luego con el anticuerpo anti-ratón conjugado con biotina, como se describió antes. Finalmente, se adicionó estreptavidina peroxidasa y las células infectadas se observaron usando una solución de DAB al 0,1 % y peróxido de hidrogeno al 0,02 %. Se adquirieron imágenes utilizando un microscopio invertido (Zeiss Axiovert 40) y el software Axio-Vision. Sobre las imágenes se contaron células totales e infectadas, utilizando la herramienta cell counter (Image J). Finalmente, se determinó el porcentaje de infección de tres cultivos independientes y cada condición se evaluó por duplicado. Los datos se analizaron con el software GraphPad Prism empleando la prueba ANOVA y el análisis post Hoc con la prueba de Tukey. Los valores p < 0,01 fueron considerados significativos. Los datos fueron presentados como promedio ± D.E.
Clonación.
Se amplificó por PCR la región codificante de la proteína NS3 de DENV2, se clonó en un vector de expresión y el constructo resultante se usó para transfectar células A549. La expresión heteróloga de la proteína NS3 fue evaluada mediante ensayos de WB e IF. Para amplificar el gen que codifica para la proteína NS3, se diseñó una pareja de oligonucleótidos a partir de la secuencia genómica de DENV2 (NC_001474.2). Oligo sentido: acc atg GCC-GGA-GTA-TTG-TGG (en minúscula la secuencia kozak) y oligo antisentido: CTT-TCT-TCC-GGC-TGC-AAA-TTC-CT. La reacción de amplificación se llevó a cabo con 1 mM MgCl2, 200 μM dNTPs, 0,5 μM de "primers", 0,5 U ADN polimerasa Taq Platinum (Invitrogen) y como plantilla se usó ADNc de DENV2. El programa de amplificación comprendió una desnaturalización inicial a 95 °C por 5 minutos, 35 ciclos de 95 °C por 30 segundos, 55 °C por 45 segundos y 72 °C por 60 segundos, seguidos por una extensión final de 72 °C por 7 minutos. El producto de PCR (1860 pb) fue purificado con el kit GenJet (Thermo-Scientific) y clonado en el vector pcDNA3.1/V5-His-TOPO (Invitrogen) siguiendo las condiciones del fabricante. La expresión en este vector lleva a la producción de proteínas de fusión con un epítope V5 y una etiqueta de histidinas (His-tag). Con el constructo pcDNA3.1/V5-His-TOPO/NS3 se transformaron bacterias TOP10 (Invitrogen) mediante choque térmico y la confirmación de colonias positivas se hizo por PCR, utilizando los oligonucleótidos del vector (T7 y BGH_reverse. Invitrogen). Finalmente se hizo la purificación del ADN plasmídico utilizando el kit GeneJET Plasmid (Thermo-Scientific).
Transfección de células A549.
Los ensayos de transfección de las células A549 con el constructo pcDNA 3.1/V5-His-TOPO/NS3 se realizaron con Lipofectamina LTX (Invitrogen), siguiendo las condiciones recomendadas por el fabricante. Se partió de células A549 que fueron cultivadas en placas de 12 pozos hasta una confluencia del 70 - 80 %, en medio DMEM (Sigma Aldrich) suplementado con SFB (10 %) a 37 °C y una atmosfera de 5 % de CO2. Una vez agregados los lipocomplejos, las células fueron mantenidas en cultivo como se describió antes por 48 horas, momento en el cual se evaluó la expresión de la proteína NS3 por WB. Para esto, extractos de proteínas fueron obtenidos, cuantificados y sometidos a SDS-PAGE y WB con anti-NS3403_599, como se describió antes. El revelado de la señal se hizo con el sistema SuperSignal™ West Pico PLUS y las imágenes fueron adquiridas usando el sistema Image Quant LAS 4000 (GE Healthcare).
Obtención de línea A549 transfectada, estable.
La obtención de una línea celular A549 transfectada estable se hizo por selección con Geneticina (Calbiochem 345812). La concentración de Geneticina a usar se estableció con la premisa que las células no transfectadas debían morir después de siete días de crecimiento bajo presión del medicamento a esa determinada concentración. Las concentraciones usadas para construir la curva de susceptibilidad a Geneticina fueron 0, 250, 500, 750, 1000, 1250 y 1500 μg/ml. Transcurridos siete días, se hizo un ensayo de MTT y los resultados fueron expresados como supervivencia relativa, tomando como 100 % de supervivencia la hallada para el control de células cultivadas sin Geneticina. Sobre la línea estable, de aquí en adelante A549_NS3, se hicieron ensayos de IF para monitorear la proporción de células transfectadas. La proteína NS3 fue detectada usando el anticuerpo anti-NS3403_599 (1:250) y un anticuerpo secundario anti-rata acoplado a biotina (1:200) (Vector. AB9401). Como control de una proteína celular, se detectó Alix con un anticuerpo monoclonal anti-Alix (1:800) (Thermo-Scientific. MA1-83977) y un anticuerpo secundario anti ratón biotinilado (1:400) (Vector. AB9200). Finalmente, las células fueron lavadas y se hizo la incubación con una solución de estreptavidina acoplada a Alexa 594® (Molecular Probes). Los núcleos fueron contrastados con Hoechst 33342 como se describió antes y las células fueron observadas bajo microscopio (Zeiss Axioimager A2) utilizando un sistema de fluorescencia X-Cite serie 120Q y el software ZEN 2012.
Ensayos de interacción proteína: proteína.
Interacción entre ALIX y NS3: ensayos de inmunoprecipitación.
Extractos de proteínas de células A549 y A549_NS3 fueron usados en ensayos de inmunoprecipitación (IP). Brevemente, se tomaron 300 μg de extracto de proteína, se adicionaron 10 μl de anticuerpo anti-NS3403_599 y la mezcla se dejó en agitación a 4 °C durante 6 horas. Luego, se adicionaron perlas de agarosa A/G (Thermo-Scientific) y se incubó a 4 °C durante toda la noche con agitación. Al día siguiente, se recuperaron las perlas por centrifugación, se recuperó el sobrenadante (fracción no unida) y se hicieron lavados con PBS más 1 mM de PMSF. El precipitado final se resuspendió en buffer Laemmli (6X) y se analizó mediante SDS-PAGE y WB, empleando los anticuerpos anti-NS3403_599 y anti-Alix.
Interacción entre ALIX y NS3: ensayos de "pull-down".
Células A549 y A549_NS3 fueron infectadas con DENV2 (como se describió arriba) y a continuación se evaluó la interacción de la recombinante NS3 con Alix. Para esto, se prepararon extractos de proteínas por sonicación de células resuspendidas en buffer de unión (PBS y 10 mM de imidazol). Luego, se cuantificaron los extractos (como se describió antes) y 200 μg de proteína fueron incubados con 100 μl de resina HisPur Ni-NTA (Thermo Scientific) durante toda la noche a 4°C en agitación constante. Al día siguiente se retiró el sobrenadante (fracción no unida) y la resina se lavó con PBS más imidazol (25 mM). Finalmente, el pellet obtenido se resuspendió en buffer Laemmli (6X) y se sometió a SDS-PAGE y WB usando los anticuerpos anti-NS3403_599 y anti-Alix (como se describió antes).
RESULTADOS
Ensayos de interferencia.
Los ensayos de MTT demostraron supervivencia de 95 a 100 % de las células tratadas con los siRNA, comparadas con el control no tratado. Los ensayos de qPCR confirmaron la reducción de la transcripción para Alix y Vps4B en las células tratadas con los siRNA específicos, al comparar con la transcripción en las células tratadas con el siRNA-NR (Fig. 1a). Por su parte, los ensayos de WB confirmaron la reducción de la expresión de las proteínas Alix (Fig. 1b) y Vps4B (Fig. 1c) en las células tratadas con los siRNA específicos. La expresión de la proteína control (actina) fue uniforme en los tres sistemas celulares (Figs. 1b y 1c).
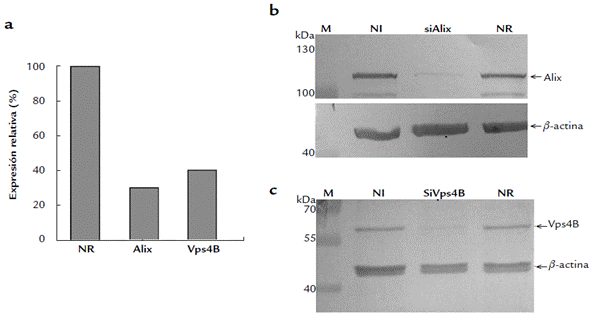
Figura 1 Disminución de la transcripción y traducción de los genes alixy vps4b en células A549: Ensayos con siRNA. (a): Cuantificación por qPCR de los transcritos de células tratadas con siRNA para Alix (Alix), células tratadas con siRNA para Vps4B (Vps4B) y células tratadas con un siRNA no relacionado (NR). Los datos se muestran como porcentajes de expresión relativa al control NR. (b): WB mostrando la expresión de Alix en células no interferidas (NI), células interferidas con siRNA para Alix (siAlix) y en células tratadas con siRNA-NR (NR). (c): WB mostrando la expresión de Vps4B en células no interferidas (NI), células interferidas con siRNA para Vps4B (siVps4B) y en células tratadas con siRNA-NR (NR). Las flechas indican el peso molecular de Alix (≈ 100 kDa), Vps4B (≈ 49 kDa) yβ-actina (≈ 42 kDa), esta última usada como control de carga. M: marcador de peso molecular (PageRuler™ Prestained Protein Ladder 10-250 kDa_Thermo Scientific). En cada carril se cargaron 30 μg de extracto de proteína.
La interferencia de Alix o Vps4B no inhibió la transcripción ni la traducción viral.
El ensayo de RT-PCR no reveló diferencias evidentes entre el transcrito nsl de las células no interferidas, las células tratadas con los siRNA específicos para Alix o Vps4B y las células interferidas con el siRNA NR (Fig. 2a). Por su parte, la traducción viral se evaluó por WB con un anticuerpo anti-NS3 y no hubo diferencia entre los tres sistemas evaluados (Fig. 2b). Estos resultados muestran que la expresión reducida de Alix o Vps4B no causa un efecto notable en la transcripción o la traducción viral.
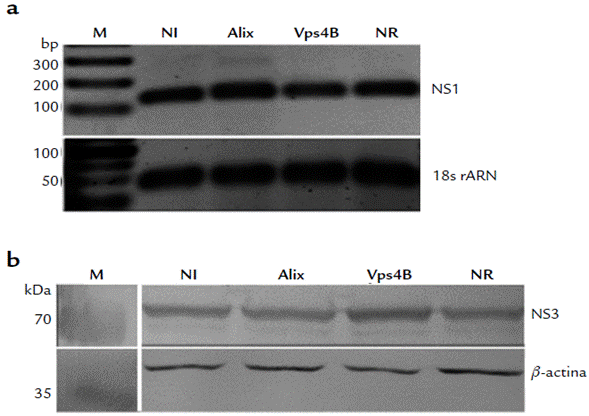
Figura 2 Efecto de la interferencia de Alix y Vps4B sobre la transcripción y traducción de DENV2 en células A549. (a): Gel de agarosa al 2 % teñido con bromuro de etidio. Mediante RT-PCR, se evaluaron los transcritos de los genes 18S ARNr y nsl en células infectadas con DENV2 pero no interferidas (NI), y en células infectadas con DENV2 e interferidas con siRNA para Alix (Alix), con siRNA para Vps4B (Vps4B) o con un siRNA no relacionado (NR). Amplicon ns1: 120 pb. Amplicon 18SARNr: 80 pb. M en el gel de NS1: marcador de peso molecular GeneRuler™ 100 bp DNA Ladder (Thermo Scientific). M en el gel de 18S ARNr: O'RangeRuler 50 bp DNA Ladder (Thermo Scientific). (b): WB mostrando la expresión de las proteínas NS3 (≈ 70 kDa) y β-actina (≈ 42 kDa) en células infectadas no interferidas (NI), células infectadas e interferidas para Alix y Vps4B, y células control (NR). M en el WB para NS3: marcador de peso molecular PageRulerTM Prestained Protein Ladder 10-250 kDa (Thermo Scientific). M en el WB para β_actina: marcador de peso molecular Spectra™ Multicolor Broad Range Protein Ladder (Thermo Scientific).
La interferencia de Alix, pero no de Vps4B, redujo la producción de DENV2 en células A549.
El efecto de la interferencia en la producción viral se evaluó usando cultivos de células A549 infectadas con DENV2 y tratadas con siRNA específico para Alix o para Vps4B. Para cada sistema, se tomó el medio del cultivo (sobrenadante) o el lisado de las células, se agregó uno u otro sobre monocapas de células LLC-MK2, y tras 48 horas de incubación se estableció el porcentaje de células infectadas. Cuando las células LLC-MK2 fueron cultivadas con los sobrenadantes o los lisados de las células A549 interferidas para Alix, se observó una reducción significativa en el número de células infectadas (Fig. 3a). En contraste, no se encontró diferencia en el número de células LLCMK-2 infectadas después de incubarlas con los sobrenadantes o los lisados de las células A549 interferidas para Vps4B (Fig. 3b). Esto sugiere que Alix participa de alguna manera en el ensamblaje o liberación de DENV2, mientras que Vps4B parece no hacerlo. El análisis se hizo comparando el porcentaje de infección entre los sistemas interferidos y los controles; estos son, las células infectadas no interferidas y células infectadas tratadas con el siRNA-NR.
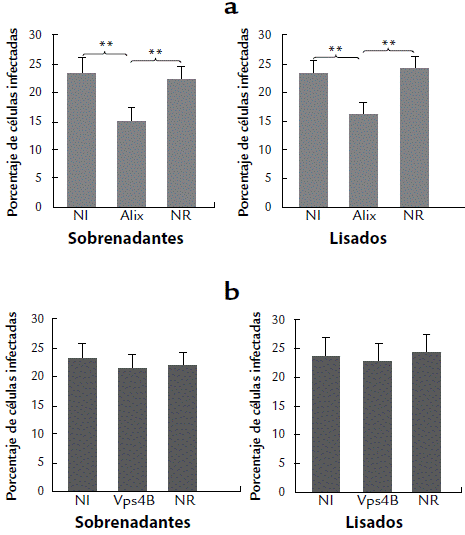
Figura 3 Efecto de la interferencia de las proteínas Alix y Vps4B sobre la producción de DENV2. Se muestra el porcentaje de infección viral en células LLCMK-2 tratadas con los lisados (para evaluar ensamblaje) o los sobrenadantes (para evaluar liberación) de células A549 infectadas con DENV2 e interferidas para las proteínas Alix (a) o Vps4B (b). Como controles se usaron células infectadas y no interferidas (NI), y células infectadas e interferidas con un siRNA no relacionado (NR). Se muestra el promedio de tres cultivos independientes y dos réplicas por condición. El doble asterisco indica las diferencias estadísticamente significativas (p < 0,01) que se obtuvieron al comparar con los controles NI y NR.
Evaluación de la expresión heteróloga de la proteína NS3 en células A549.
Se estableció una línea celular A549_NS3 en la que la proteína NS3 se expresaba de manera estable en el 100 % de las células bajo presión con Geneticina. La eficiencia de la transfección se confirmó mediante ensayos de WB (Fig. 4a) con el anticuerpo anti- NS3403_599. En las células transfectadas se observó una señal a la altura de «74 kDa, la cual correspondió al tamaño esperado para la proteína NS3 recombinante. Por su parte, los ensayos de IF sobre un cultivo A549_NS3 evidenciaron la expresión de NS3 en el 100 % de las células (Fig. 4b) y su localización citoplasmática, con una mayor intensidad hacia la región perinuclear. No hubo marcación en las células A549 no transfectadas. Adicionalmente, se realizó la inmunodetección de Alix como control de una proteína celular (Fig. 4b), y como era de esperarse, Alix se detectó tanto en células transfectadas, como en las células no transfectadas. Aunque el "merge" evidenció que NS3 y Alix colocalizan, ese resultado no permite establecer si hay una interacción directa entre ellas o si simplemente ocupan el mismo compartimento celular.
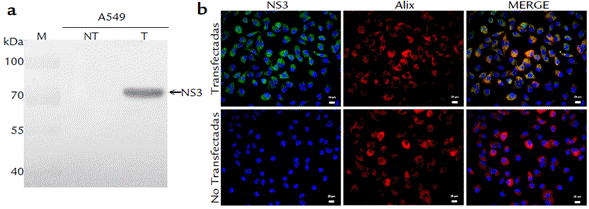
Figura 4 Expresión de la proteína NS3 en células A549 transfectadas. (a): WB con anticuerpo anti-NS3403_599 sobre células transfectadas con un constructo que expresa la proteína NS3_His-tag (T) y sobre células no transfectadas (NT). Se indica la posición de la proteína NS3 recombinante (≈ 74 kDa). Se cargaron 30 μg de extracto de proteína celular en cada carril. M: Marcador de peso molecular Page-RulerPlus (Thermo-Scientific). (b): Inmunofluorescencia (IF) usando anti-NS3403_599 como anticuerpo primario, sobre una línea celular A549 transfectada y expresando establemente la proteína NS3 recombinante. La IF se detectó con estreptavidina acoplada a Alexa 594® (Molecular Probes) y los núcleos fueron contrastados con Hoechst 33342 (Sigma Aldrich). Barra de 20 μm.
Evaluación de la interacción entre la proteína NS3 y Alix: ensayos de inmunoprecipitación.
Para indagar sobre la posible interacción entre NS3 y Alix, se hicieron ensayos de inmunoprecipitación (con el anticuerpo anti-NS3403_599) sobre células de la línea A549_ NS3. Una vez se hizo la IP, se prepararon dos membranas para hacer WB con el anticuerpo anti-NS3403_599 y con el anticuerpo anti-Alix. Como se había demostrado en un trabajo anterior, la proteína NS3 fue atrapada con el anticuerpo anti-NS3403_599 de manera específica (Fig. 5a) (Morales et al., 2017). Sin embargo, al usar el anticuerpo anti-Alix no se detectó señal alguna (Fig. 5a). Estos resultados indican que no existe una interacción directa entre NS3 y Alix o que en este ensayo esa interacción no pudo ser revelada. Esto último podría deberse a que el sitio de interacción entre NS3 y Alix es bloqueado cuando el anticuerpo anti-NS3403_599 se une a la proteína viral.
Evaluación de la interacción entre la proteína NS3 y ALIX: ensayos de "pull-down".
Para retar el planteamiento anterior sobre el bloqueo de la interacción NS3 y Alix (mediada por la unión entre NS3 y el anticuerpo anti-NS3403_599), se hicieron ensayos de "pulldown" con la línea A549_NS3. En este sistema, la proteína NS3 recombinante se puede anclar a través de su His-tag a una resina de níquel, dejando toda la secuencia de NS3 disponible para interactuar. En las células A549_NS3, la proteína NS3 fue detectada en el extracto (Fig. 5b, carril "Ext") y también en el "pull-down" (Fig. 5b, carril "Pull"). Se descartó que la señal observada en el carril "Pull" fuera unión inespecífica, ya que los carriles PL y UL (que corresponden a los sobrenadantes que resultaron del primer y último lavado de la resina) estaban limpios. Adicionalmente, en las células A549 infectadas con DENV2, pero no transfectadas, la proteína NS3 fue detectada en el extracto crudo pero no en el "pull-down". Esto confirma que la resina no unió inespecíficamente a NS3 y que las interacciones que se observaron en el "pull-down" del sistema A549_NS3, se dan sólo a través del His-tag. Cuando se hizo el WB con el anticuerpo anti-Alix, se reveló que la proteína Alix co-precipita con la recombinante NS3 en las células A549_NS3 (Fig. 5b). En las células A549 infectadas con DENV2 (pero no transfectadas), no hubo señal alguna de Alix en el "pulldown" (Fig. 5b).
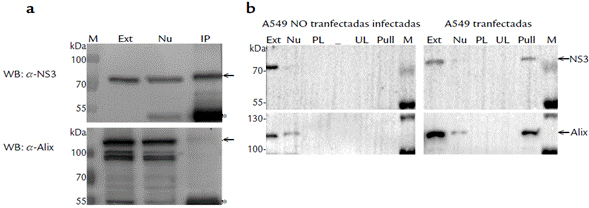
Figura 5 Evaluación de la interacción NS3/Alix: Ensayos de co-inmunoprecipitación y "pull-down". (a): Ensayo de co-inmunoprecipitación. M: Marcador de peso molecular Page-RulerPlus (Thermo-Scientific). Ext: extracto de células A549 transfectadas con un constructo que expresa la proteína NS3_His-tag. Nu: Fracción no unida (sobrenadante pos-inmunoprecipitación. IP: inmunoprecipitación con el anticuerpo anti-NS3403_599. Arriba: WB revelado con el anticuerpo anti-NS3403_599. Abajo: WB revelado con el anticuerpo anti-Alix. Las flechas indican las bandas correspondientes a NS3 (arriba) (≈ 74 kDa) y Alix (abajo) (≈ 100 kDa). Los asteriscos señalan las cadenas pesadas de las inmunoglobulinas. (b): Ensayos de "pull-down" usando células A549 infectadas con DENV2 y células A549 no infectadas, pero transfectadas con un constructo que expresa la proteína NS3_His-tag. El "pull-down" se hizo con una resina de unión a His-tag (Ni-NTA). Al terminar el ensayo, la resina fue cargada en un SDS-PAGE y analizada por WB con los anticuerpos anti-NS3403_599 y anti-Alix. Ext: extracto celular. Nu: Fracción no unida; sobrenadante recuperado después de la incubación con la resina y justo antes de los lavados. PL: Sobrenadante del primer lavado de la resina. UL: Sobrenadante del último lavado de la resina, antes del "pull-down". "Pull": "pull-down". M: Marcador de peso molecular Page-RulerPlus (Thermo-Scientific). Las flechas señalan las bandas correspondientes a la proteína NS3 recombinante (≈ 74 kDa) y Alix (≈ 100 kDa).
DISCUSIÓN
El dengue es una enfermedad de impacto global cuyo tratamiento es sólo de apoyo, debido a que no hay antivirales específicos o una vacuna aprobada. A pesar de esto, es poco lo que se sabe sobre los mecanismos de patogénesis del DENV (Whitehorn y Farrar, 2010) y eso incluye a las estrategias del virus para superar la barrera física que representa la membrana plasmática durante su entrada (infección) y salida (liberación) de la célula hospedera.
En este estudio se estableció que la expresión reducida ("knock-down") de Alix o Vps4B no inhibe la transcripción o la traducción de DENV2 en células A549. También se demostró que el "knock-down" para Vps4B no altera el ensamble o la liberación viral, mientras que el "knock-down" de Alix tiene efecto en ambos procesos.
Los componentes de ESCRT comprenden los complejos ESCRT-0, -I, -II y III, además de la proteína VPS4 y varias proteínas auxiliares, incluyendo a Alix. VPS4 es una proteína esencial para la formación de cuerpos multivesiculados (MVB), ya que disocia los complejos ESCRT-III de los MVB y de ese modo asegura su disponibilidad para nuevas rondas de formación de vesículas (Babst et al., 1998). Algunos estudios han demostrado que VPS4 es necesaria para la gemación de ciertos virus ARN (Demirov y Freed, 2004). Por ejemplo, la liberación de HCV se ve significativamente afectada en sistemas dominante-negativo (Corless et al., 2010; Tamai et al., 2012) o "knock-down" (Ariumi et al., 2011) para Vps4B. En contraste, no hubo efecto significativo en la liberación de WNV al usar un transdominante de Vps4B que impedía la disociación de los componentes de ESCRT-III (Garg et al., 2013). Del mismo modo, Tabata y colaboradores encontraron que VPS4B no era importante para la liberación de DENV y JEV (Tabata et al., 2016). En este trabajo, nosotros encontramos que el "knock-down" de Vps4B no afectó el ensamble o la liberación de DENV2. Esto sugiere que la liberación de este virus podría obedecer a una fisión de membranas, más que a un estrangulamiento mediado por Vps4B.
Respecto a Alix, en este trabajo demostramos que el "knock-down" de esta proteína no inhibe la transcripción ni la traducción DENV2, pero sí disminuye el ensamble y la liberación viral. Nuestros resultados son diferentes a los reportados por Tabata y colaboradores en 2016, pues ellos encontraron que el bloqueo de la expresión de Alix no afectaba la liberación de DENV2. Como nosotros, Tabata y colaboradores lograron el "knock-down" de Alix mediante interferencia, usando un siRNA diseñado por ellos en un trabajo anterior (Morita et al., 2010). En nuestro caso, usamos un "pool" de cuatro siRNAs que demostraron alcanzar una reducción de ~ 70 % de los transcritos de Alix (Fig. 1a) y una disminución notable de la expresión de la proteína (Fig. 1b) (La casa comercial anuncia entre 75 y 80 % de silenciamiento). Sin embargo, a pesar del goteo de expresión, vimos que el "knock-down" de Alix tuvo un efecto negativo y significativo en la producción de DENV2. Algo notable es que aunque Tabata y colaboradores concluyeron que Alix no está relacionada con la liberación de DENV o JEV, ellos encontraron que Alix se acumulaba en fracciones enriquecidas con organelos en los cuales sucede la replicación de JEV (no hay datos para DENV).
Finalmente, aunque esto no dirime la controversia, fue publicado un trabajo que apoya nuestros resultados. En este estudio los autores encontraron una disminución significativa de la producción de DENV2 en sistemas "knockdown" o "knock-out" (silenciamiento total) para Alix, los cuales fueron obtenidos con siRNAs y el sistema CRISPR-Cas9, respectivamente (Thepparit et al., 2019).
Una vez se estableció el efecto que tenía la expresión disminuida de Alix en el ensamblaje y liberación de DENV2, la pregunta que surgió fue si Alix podía interactuar con NS3. Esta proteína fue de particular interés, ya que hasta ese momento, las únicas interacciones reportadas entre proteínas virales y proteínas ESCRT en Flavivirus, eran NS3_Alix para YFV (Carpp et al., 2011) y NS3_Tsg101 para JEV (Chiou et al., 2003; Tabata et al., 2016). Los resultados encontrados en los ensayos de IP con un anticuerpo anti-NS3 no revelaron interacción alguna, mientras que en los ensayos de "pull-down" se detectó simultáneamente a Alix y a NS3. Creemos que los ensayos de IP no revelaron la interacción debido a la naturaleza del anticuerpo policlonal anti-NS3 usado, ya que fue producido contra una proteína recombinante que abarcaba los aminoácidos 403 a 599 de NS3. En ese orden de ideas, suponemos que el anticuerpo unido a esta región sobre NS3, podría ocultar el sitio de unión a Alix. Esta hipótesis puede apoyarse en el trabajo de Khadka y colaboradores, en el cual se construyeron plásmidos con la secuencia completa o parcial de cada proteína de DENV2, para retarlos luego contra una librería de ADNc humano y revelar interacciones entre las proteínas virales y las proteínas celulares. Específicamente para NS3, los nueve constructos resultantes contenían la secuencia codificante para la proteína completa (residuos 1 a 617) y los fragmentos 1 a 180, 181 a 462, 463 a 617, 1 a 343, 344 a 462, 1 a 462, 181 a 617 y 344 a 617. Entre todos los constructos (de las diez proteínas virales) con los que se retó la genoteca, Alix sólo interactuó con tres de ellos y todos correspondían a clones NS3: el clon con la secuencia completa, el clon con los residuos 344 a 617 y el clon con los residuos 463 a 617 (Khadka et al., 2011). Esto sugiere que la secuencia de unión NS3_Alix podría estar alojada en la región 463 a 617 de NS3, la cual pudo estar enmascarada en los ensayos de IP debido a que el anticuerpo anti-NS3 fue producido contra la región 403 a 599.
Por el contrario, en los ensayos de "pull-down" la extensión total de la proteína NS3 se encontraba disponible y fue posible revelar su interacción con Alix. El "pull-down" permitió ver que NS3 y Alix precipitaron juntas, pero no permitió establecer si la interacción es directa o indirecta.
CONCLUSIONES
Se estableció que el "knock-down" de Alix o Vps4B no inhibe la transcripción o la traducción de DENV2. Por el contrario, el "knock-down" de Alix afectó notablemente el ensamble y la liberación viral, y eso convierte a esta proteína en un blanco interesante para el estudio de interacciones virus-hospedero. La exploración de estas potenciales interacciones y el desarrollo de estrategias para interrumpirlas, pueden ser el punto de partida para desarrollar terapias antivirales encaminadas a reducir el ensamble y la liberación de DENV.


