











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
The objective of this review article is to describe the current relevance, the main mechanisms presented, and the uses of antimicrobial peptides (AMPs), as new therapies in the clinical area are being evaluated. The information used was mainly compiled from scientific articles with data on human AMPs and their different applications, based on a systematic review of scientific papers and searching without date limits and only papers in English and Spanish. Gray literature was accessed through manual search; no restrictions were made involving study design for a retrospective study. AMPs are an essential part of innate immunity that evolved, in most living organisms, over 2.6 billion years to combat microbial challenge against various bacteria, viruses, fungi, and parasites. The treatment of bacterial infections is increasingly complicated by the ability of bacteria to develop resistance to antimicrobial agents, making it essential to find new therapeutic alternatives (WHO, 2012), and a new therapeutic alternative is antimicrobial peptides.
Antimicrobial peptides
With the increase in antibiotic resistance, the search for alternative antibiotics has become a priority for the treatment of impending antibiotic-resistant strains; therefore, the study of peptides is relevant because they comprise diverse amino acid sequences and structures and perform functions that are essential actors in biological systems, facilitating the effective management of the necessary information that provides the basis for revealing the unanswered questions on the natural behaviors of organisms. It is becoming more urgent to find alternative options for the treatment of antibiotic-resistant strains; as a result, AMPs have been proposed as possible candidates for their use as antimicrobial agents since their mode of action is assumed to be substantially different from existing antibiotics, mainly due to their extensive spectral activity and their low resistance potential (Maroti et al., 2011).
AMPs are a family of approximately 900 molecules that are part of the most primitive innate immune systems in vertebrates, insects, and plants (Tomasinsig et al., 2010). Therefore, an AMP database exists that allows efficient search, prediction, and design of peptides with antibacterial, antiviral, antifungal, antiparasitic, insecticidal, spermicidal, anticancer, chemotactic, immunological modulation or antioxidant activities. Genes encode most AMPs and synthesized by ribosomes, although others are products of secondary metabolites such as lactoferricin, a globular protein isolated mainly from bovine milk, which is synthesized by neutrophils in the blood. Lactoferricin deprives free iron from microorganisms, such as E. coli, that need it to grow and invade the host.
Among the range of peptide, classifications are cationic antimicrobial peptides (AMPs), which are relatively small peptides that have a net positive charge and exhibit some antimicrobial activity, often of broad-spectrum, allowing them to be a source of innate defense against microbial infections (Wang, 2015). Based on Wang et al. (2016), the APD3 database lists more than 2600 examples of AMPs, of which 112 are human peptides, and 100 of those have antimicrobial activities. These examples, in addition to their families and most essential characteristics, are shown in Table 1.
It is important to emphasize that AMPs differ fundamentally from antibiotics in their pharmacodynamic characteristics; the pharmacodynamic differences combine producing a much lower probability of resistance that evolves against antimicrobial peptides (Rodriguez and McDaniel, 2001). Many of the antibiotics produced by microorganisms are peptide molecules. These are produced by the synthesis of nonribosomal peptides (NRPS), a process that involves the expression of large sets of genes that encode multiple enzymes that work sequentially to catalyze the sequence of chemical reactions necessary to synthesize the antibiotic (Yu et al., 2017), and the manipulation ofmodular polyketide synthases (PKSs), which has led to the production of, for example, various erythromycin analogs (Marahiel, 2016).
Antibiotics based on peptides include β-lactamases, such as penicillin cyclic peptide antibiotics, such as polymyxins and bacitracin, glycopeptides, such as vancomycin (Yim et al., 2014), and the lipopeptide daptomycin, one of the most recent antibiotic classes introduced (Robbel and Marahiel, 2010).
AMPs, including those made in human cells, contrast with NRPS antibiotics because AMPs are produced by the normal process of ribosomal translation on an mRNA template. The first product is usually a preprotein that is then processed to the final length of the active AMP. The difference in the genetic origins of AMP and NRPS-antibiotics has consequences for the compositions of the final products since the ribosomally-produced AMPs contain only the usual complement of amino acids found in proteins, and NRPS, on the other hand, are not limited by ribosomal translation and usually contain a mixture of typical amino acids along with noncanonical amino acids that are not found in proteins (Walsch et al., 2013).
There are two common characteristics of most antimicrobial peptides regardless of their structure or size. First, AMPs have a positive charge due to the presence of a large number of basic amino acids (mostly lysine and arginine), and second, approximately 50 % of the amino acids that constitute them are hydrophobic, which enables them to interact with the membranes (Andersson et al., 2016;Waghu et al., 2016). Also, defensins attack the surface of the membrane that surrounds a pathogen, generating holes in the membrane, and beta-defensins such as hBD-1 and hBD-2 become active against gram-negative bacteria. With an abundant expression of hBD-2 in gram-positive infections, the inflammatory response and vasodilation increase and stimulation of cytosines or bacteria are required. hBD-3 is active against gram-positive and gram-negative bacteria and induced by the pathogenicity of microorganisms (PAMP and by TNF-alpha, IL-1beta and IFN-gamma) (Wang, 2015).
Interaction of AMPs with the bacterial cell membrane
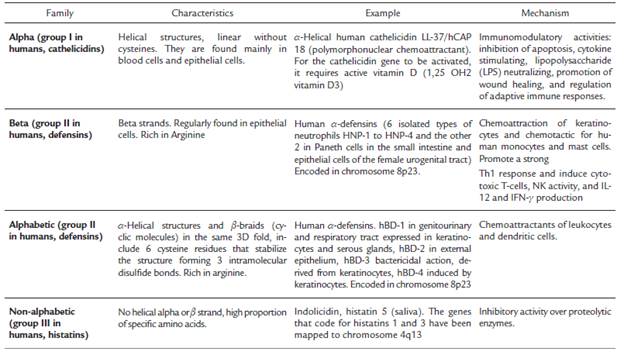
The barrel model suggests that peptides accumulate on membrane surfaces and insert into the membrane when a threshold amount is reached. Interactions between the peptide and lipid hydrophobic side chains are then formed, creating a hydrophilic pore, which produces an osmotic imbalance and disrupts the membrane potential. Conversely, the toroidal pore model suggests that the peptide causes the formation of pores stochastically, requiring fewer peptides for inferred pores (Rivera et al., 2007); however, these models are based on studies performed on lipid vesicles and cannot fully explain the interaction of AMPs with the complex bacterial cytoplasmic membrane.
On the other hand, the aggregate mechanism is like how detergents act, in which the peptide binds to the membrane, forming structures similar to the mycelia, and a channel is formed, through which ions are released, causing cell death due to loss of cytoplasmic content. Because AMPs that act on vegetative cells cause damage to the membrane, there is a loss of transmembrane potential and essential molecules (Zhang and Gallo, 2016), and it has been demonstrated that damage to the membrane disturbs cellular homeostasis, leading to an increase in cell volume and a decrease in total cell numbers (Nguyen et al., 2011). The formation of pores or channels in cells has been studied using fluorescent dyes such as propidium iodide (Nguyen et al., 2011) and Sytox® Green (Lee et al., 2015).
Certain studies have only examined the damage on the membrane, which does not indicate whether the membrane is the initial and only objective of the AMP or if the peptide moves through the membrane into the cytoplasm to target another essential cellular function, such as DNA and RNA synthesis. However, a study by Barns and Weisshaar (2013) suggests that at a low peptide concentration, the peptide interacted with the membrane, but the damage to the membrane was repairable; on the contrary, at a high peptide concentration, a threshold was reached that caused irreversible damage to the membrane. Therefore, it is necessary to give intervals of lethal and sublethal concentrations to obtain more information on how the peptide is directed to the cell. It is also known that rhodamine labeling (Rh-LL-37) has different effects on antimicrobial activity against E. coli, suggesting that this compound binds to peptidoglycan. In other studies, cecropin A (an antimicrobial peptide with leishmania activity) containing CL (anionic phospholipids) causes dissociation in cellular proteins, leading to cellular homeostasis (Wang et al., 2013).
It has also been shown that human a-defensin 5 translocates to the cytoplasm of E. coli and accumulates at the site of cell division, and buforin antimicrobial peptide analogs (Fig. 1) exert their bactericidal activity on E. coli by binding to DNA and RNA after penetrating the cell membrane (Barns and Weisshaar, 2013).
AMP resistance mechanisms
The mechanisms ofresistance to AMPs include proteolytic degradation or sequestration by secreted proteins, impedance by exopolymers and biofilm matrix molecules, elusion of cell surface attraction/membrane alteration and export by efflux pumps. Segregated bacterial proteins, such as proteases, are the first bacterial defense mechanisms that AMPs encounter when interacting with bacteria.
The proteolytic degradation of AMPs by extracellular enzymes represents a simple but effective way to provide resistance of AMPs to microorganisms. Commensal bacteria that live on the epithelial surfaces of mammals, such as staphylococci, secrete various proteases, including metalloproteases such as aureolysin and SepA, and serine endopeptidases such as V8 protease, which are known to degrade linear AMPs, such as human cathelicidin LL-37 (Omardien et al., 2016).
Group A Streptococcus produces a protease called SpeB, and this cysteine protease has been shown to fragment many host AMPs, including LL-37 and 3-defensins (Sieprawska-Lupa, 2004; Hao et al., 2013). Interestingly, the exploitation of host proteins can enhance the proteolytic activity of SpeB; interactions between SpeB and cell wall anchored G-related linkage of alpha2M (GRAB) proteins brings SpeB complexed to the host inhibitory proteinase alpha2-macroglobulin, which shows increased activity towards LL-37, to the membrane surface (Nguyen et al., 2011;Lee et al., 2015).
In addition, as a secondary effect of SpeB proteolytic activity, proteoglycans degrade host-released dermatan sulfate, which completely neutralizes human alpha-defensin, HNP-1. Finally, the proteases of another gram-positive pathogen, Enterococcus faecalis, and the gram-negative Pseudomonas aeruginosa and Proteus mirabilis have also been reported to degrade LL-37 (Nelson et al., 2011).
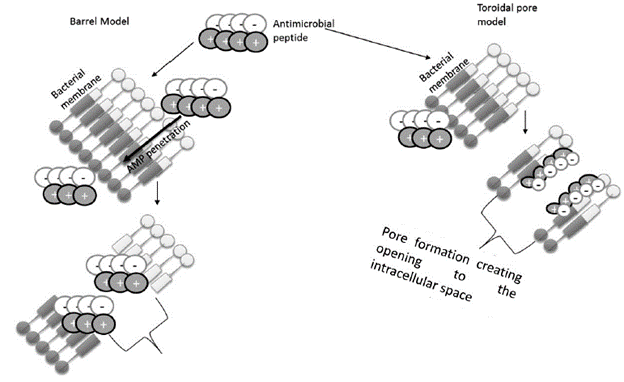
For what has been considered the continued high interest in AMPs as potential therapies for bacterial infections, surprisingly few studies have tried to assess the risk of developing resistance and explore the mechanisms of how it is acquired. The methods to determine this resistance are the broth microdilution method, to obtain the minimum inhibitory concentration (MIC), and the other is the time of death test. The molecular basis of acquired resistance has been characterized in bacterial pathogens through the isolation of resistant strains of natural origin. The serial passages in the presence of AMPs and the direct placement of AMPs are shown in Table 2. (Schmidtchen et al., 2001; modified from Frick et al., 2011;Dobias et al., 2017;Hashemi et al., 2017).
Defensins are essential components of the defense mechanisms in epithelial cells and constitute a family of small cationic peptides (3 kDa-6 kDa). They are classified as defensins α, β and θ. An antimicrobial agent present in bovines is the calcium-binding protein S100A7, also known as psoriasin (Chapman et al., 2016). To date, little is known about its physiological function in bovines, particularly in the mammary gland. In humans, it has been observed to protect the skin from infections against E. coli, although it is also expressed in healthy skin. The importance of β-defensins and psoriasin in the local defense against mastitis lies in the constitutive and inducible expression of the antimicrobial peptides in bovine mammary glands and its antibacterial activity against pathogens.
AMPs for therapeutic use
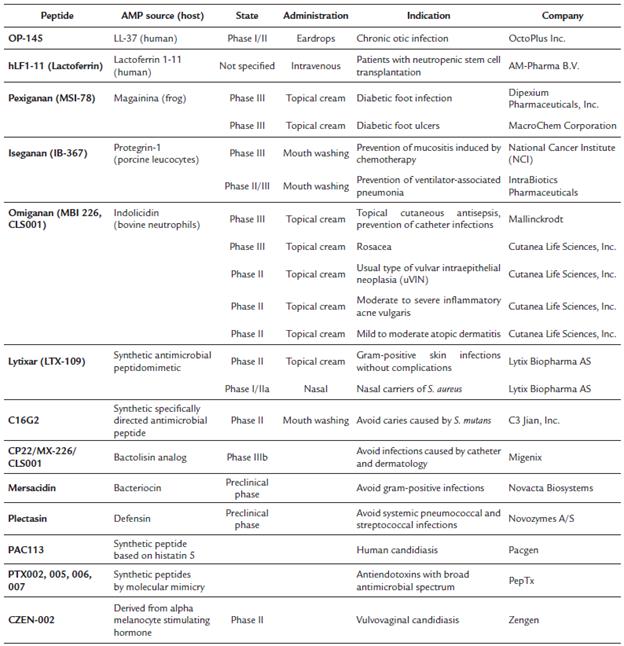
Currently, there are many AMPs in clinical development for the treatment of various bacterial pathogens, but most of them are intended for topical use only (Table 3). This is probably a direct result of the toxicity observed after the systemic administration of polymyxin B and colistin. The only AMP in clinical trials for intravenous administration is lactoferrin 1-11 (hLF1-11) of human origin to treat life-threatening infections that occur in patients with stem cell transplantation. A study showed that hLF1-11 is well tolerated in single and multiple doses as high as 5 mg after intravenous administration (Tetens et al., 2010).
In contrast to standard antibiotics, AMPs are effective against quiescent and actively growing bacteria; they do not require metabolic processes for antimicrobial activity; they show rapid effectiveness kinetics (seconds or minutes) and demonstrate a low propensity for bacterial resistance in vitro. However, AMPs have several limitations that have delayed their successful development for clinical use, including the inhibition of activity in the presence of acidic pH and in a pulmonary environment.
Over the last two decades, there has been a focus on the rationality of engineering AMP amphipathic structures to overcome these intrinsic limitations. Recently, cationic AMP engineering activity (eCAP) has been mentioned. Recent advances in the design of eCAPs and the technology used to produce them have prompted a renaissance of interest in their therapeutic potential. Numerous groups have achieved greater efficacy and reduced toxicity with eCAPs and have demonstrated their ability to kill multiresistant and selected pathogens. Perhaps the most exciting is the recent reports of eCAPs that prevent and disrupt biofilms formed by important human pathogens (Van der Velden et al., 2009; Tellez and Castaño, 2010; Lashua et al., 2016).
MATERIALS AND METHODS
This systematic review was carried out in July of 2018. To conduct the most complete possible compilation, literature was collected using the databases ScienceDirect, Redalyc, Web of Science, Scopus, SciELO, and Google Scholar in English and Spanish without restricting the publication year. We searched for references that contained in the title, abstract, and/or keywords the following search terms with all possible combinations: antimicrobial peptides, antimicrobial peptide products, antimicrobial peptide effects. After eliminating the duplicated documents, we selected original articles, book chapters and theoretical references (i.e., reviews, viewpoints, comments). Gray literature, such as theses, congress memories, and technical reports, were excluded from the analysis, and no restrictions were made involving study design for a retrospective study.
DISCUSSION AND CONCLUSIONS
Since the isolation of the first peptide from frog skin in 1983, several breakthroughs have been made in the isolation, synthesis, and application of AMPs. Nevertheless, many challenges have yet to be overcome in the field of AMP peptidomics, reflecting mainly their behavior in vivo and their structure-function relationships. In terms ofcosts, producing AMPs can be several hundred times more expensive than the production of conventional antibiotics; in terms of the increase in bacterial multi resistance, many resistances are based on changes in the physico-chemical properties of surface molecules and the cytoplasmic membrane. These changes usually confer moderate levels of resistance and are relatively nonspecific. It is possible that these proteins are not exclusively involved in AMP resistance; thus, they can be an alternative for clinical use.
For drug development efforts in the field of AMPs, it is essential that resistance to AMPs, which can develop due to selective pressure, does not rely on dedicated resistance genes that are conferred by horizontal gene transfer, as in the case of many mechanisms of resistance to antibiotics (Juhas, 2015; Melvin et al., 2016).
Even if many details regarding bacterial resistance to AMPs are uncovered, the therapeutic potential of AMPs that lack direct antimicrobial activity but have immunostimulatory properties that enhance natural innate immunity requires further assessment. Although some early preclinical studies have been encouraging, much more data need to be obtained before the value of this intriguing proposition can be adequately evaluated. In summary, although antimicrobial peptides are generally recognized as essential components of natural host innate immunity against the microbial challenge, their promise as a new class of drugs and their value as external therapeutic agents remains to be revealed.
Another major hurdle facing commercialization of peptides is their relatively short shelf life and potential for degradation during storage (Eckert, 2011). Several solutions for this have been proposed, including chemical modification of functional groups within the peptide and various formulation strategies, including loading peptides into various nanoparticles or encapsulating peptides in lipid vessels (da Costa, 2015). Additionally, the use of recombinant fusion peptides allows higher yields of soluble proteins and is potentially a more cost-efficient alternative to solid-phase synthesis chemistry (Haney, 2013).
Several issues require further investigation, such as membrane repair mechanisms, the role of fluidity/rigidity of the membrane, the participation of host components, the mechanisms of resistance of gram-negative membrane receptors (Juhas, 2015; Lohner et al., 2017), as well as continuing to innovate and be able to mass-produce and market new molecules based on the findings. Several studies using AMPs in Drosophila in vitro have demonstrated the potential for synergistic interactions of AMPs in the microbial killing. Hanson et al. (2019) found that certain combinations of AMPs have synergistic contributions to defense against P. burhodogranariea; synergistic loss of resistance may arise in a general fashion: cooperation of AMPs using similar mechanisms of action may breach a threshold microbicidal activity that pathogens are no longer able to resist. For instance, the action of the bumblebee AMP Abaecin, which binds to the molecular chaperone DnaK to inhibit bacterial DNA replication, is potentiated by the presence of the pore-forming peptide Hymenoptaecin (Rahnamaeian et al., 2016).
The approach of using multiple compound mutants, now possible with the development of new genome editing approaches, was especially useful in deciphering the logic of immune effectors. Understanding the role of AMPs in innate immunity holds great promise for the development of novel antibiotics (Chung et al., 2017) and will be predicting key parameters that predispose individuals or populations to certain kinds of infections (Chapman et al., 2018). It will be necessary to study the role of AMPs in not only systemic immunity but also local immune responses, and the various roles that AMPs may play in aging, neurodegeneration, anti-tumor activity, regulation of the microbiota, etc. An approximation at this are toxins found in scorpion venom might be useful to design drugs, tumoral markers, and adjuvants to cancer treatment taking advantage of their high affinity and specificity for certain ionic channels over expressed in many malignant cells. Should be taken into account that these same substances can have other applications, e.g., treatment of infectious diseases taking advantage of their wide spectrum and low resistance that they could generate (Rave et al., 2019).

