











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los arrecifes de coral suministran una gran diversidad de hábitats para muchas especies, siendo los peces uno de los grupos más variados. El ciclo de vida completo de estas especies tiene lugar en dichos arrecifes, donde encuentran lugares de reproducción, alimentación, crianza y refugio (Mumby et al., 2004). Sin embargo, los peces arrecifales se han visto amenazados por factores naturales y antropogénicos como el cambio climático, la acidificación, los cambios del nivel del mar, enfermedades, tormentas, la sobreexplotación de recursos y las prácticas de pesca inapropiadas (Pet-Soede et al., 1999; Veron et al., 2009). Es importante conocer la dinámica reproductiva de las comunidades ícticas asociadas a los sistemas arrecifales, ya que ayuda a comprender su funcionamiento ecológico y biológico. Además, la dinámica reproductiva se ha utilizado para describir y determinar cambios evolutivos como crecimiento, tamaño, comportamiento, identificación entre especies y diferenciación entre machos y hembras (Francis, 2006).
La familia Sciaenidae es de importancia comercial y deportiva, habita en los arrecifes coralinos y tiene una amplia distribución (Bannikov et al., 2009). La familia está compuesta por diez subfamilias, 67 géneros y 283 especies aproximadamente (Nelson et al., 2016). Los esciénidos o corvinas se caracterizan por emitir diferentes sonidos gracias a músculos que accionan la vejiga gaseosa para que funcione como cámara de resonancia, por lo que también son llamados tambores (Bannikov et al., 2009; Santos et al., 2013). Son habitantes frecuentes de los arrecifes, donde reposan en el día y migran a zonas adyacentes para alimentarse en la noche (Krumme, 2009).
El género Pareques Gill 1876 está ampliamente distribuido en América tropical con siete especies (Castro et al., 1999; Robertson et al., 2015; Nelson et al., 2016), de las cuales P. acuminatus (Bloch y Schneider 1801) es un representante en los arrecifes del Caribe colombiano (Polanco et al., 2011). Pareques acuminatus se encuentra en la lista roja de especies amenazadas en categoría LC (baja preocupación) (Chao et al., 2015).
Pareques acuminatus es una especie carnívora de tamaño pequeño (23 cm), que se alimenta de invertebrados móviles como braquiuros, generalmente xánthidos y fauna constituyente de pastos marinos; habitualmente se encuentra en aguas claras y someras con un ámbito de profundidad hasta 70 m (Hernández et al., 2007; Dubiaski-Silva y Masunari, 2008; Simón et al., 2016). Se puede observar en formaciones coralinas y rocosas asociadas con Sargassum cymosum, formando grupos pequeños en la base de tales complejos estructurales (Accioly y Molina, 2008; Dubiaski-Silva y Masunari, 2008; Honorio et al., 2010).
Los estudios sobre la biología y la ecología de P. acuminatus son escasos, así como sobre su importancia en el ecosistema arrecifal. En Colombia no se han realizado trabajos concernientes a la especie, por lo cual se pretende ampliar el conocimiento biológico y conocer algunos aspectos ecológicos de los hábitats arrecifales de la región de Santa Marta, así como aportar al acervo informativo sobre los peces arrecifales del Caribe colombiano.
MATERIALES Y MÉTODOS
Área de Estudio
Entre marzo de 2017 y abril de 2018 se realizaron 15 visitas a los arrecifes de cinco localidades. El Banco de las Ánimas se localiza en la plataforma continental del Golfo de Salamanca, al norte de la Ciénaga Grande de Santa Marta (11°07'N, 74°31'W), con profundidades entre 15 y 50 m (Bula-Meyer y Díaz-Pulido, 1995); Playa Blanca está a 4,5 km de El Rodadero (11°21'N, 74°24'W) en la bahía de Gaira y se caracteriza por presentar un oleaje leve o moderado (Herrera, 2010); la bahía de Taganga (11°26'N, 74°20'W) presenta una gran diversidad de formaciones coralinas de fondos arenosos (Agenda Caribe, 2006); la ensenada de Granate, ubicada en el Parque Natural Nacional Tayrona (PNNT) (11°29'N, 74°19'W), tiene un desarrollo coralino con una profundidad entre 5 y 24 m (Garzón y Cano, 1991); isla Aguja, también en el PNNT (11°31' N, 74°20'w), posee un sustrato coralino rocoso con profundidades hasta 40-45 m (Botero, 1987).
Censo de especies en cada localidad
Se empleó el método de evaluación visual cronometrada según Caldwell et al. (2016). Inicialmente, en cada localidad un observador realizó un censo visual por 30 min alrededor de cada formación arrecifal. Adicionalmente, se registraron por fotografías y videos las especies de peces dentro del arrecife donde P. acuminatus estuvo presente. La captura de los especímenes se realizó por pescadores artesanales mediante redes de mano y luego se depositaron en bolsas plásticas y se refrigeraron para su conservación y transporte al laboratorio. Los registros de video y fotografías se analizaron para determinar las preferencias de hábitat de P. acuminatus, para lo cual se describieron las características de los lugares donde se observó la especie.
Morfometría y Merística
Se realizó la caracterización de cada individuo con la metodología recomendada por González et al. (2011). Se tomaron 28 medidas de cada ejemplar, pero para el análisis comparativo se consideraron solo diez, de acuerdo con lo registrado por Cervigón (1991): Longitud de las aletas pélvicas (L.A.Pe), Longitud de la cabeza (L.C), Diámetro del ojo (D.O), Longitud interorbital (L.I.O), Altura del cuerpo (A.C), Altura del pedúnculo caudal (A.P.C), Longitud del pedúnculo caudal (L.P.C), Longitud de la aleta pectoral (L.A.P), Longitud de la mandíbula (L.M) y Longitud del rostro (L.R). Los datos se presentan en porcentajes de la longitud estándar (LS). Además, se tomó el peso total (PT) de cada individuo en una balanza analítica (Ohaus Pioneer, precisión de 0,001 gr). En la merística se tomaron ocho conteos, cinco de los cuales se compararon igualmente con Cervigón (1991). Se incluyó el conteo de las branquiespinas, para lo cual se extrajeron los arcos branquiales de cada pez y se colocaron en una caja de Petri para separar y enumerar tales elementos.
Con los datos se obtuvieron valores de frecuencia y se realizaron los estadísticos descriptivos (promedio, desviación estándar, máximo y mínimo). Se realizaron histogramas de frecuencia de tallas de los machos, hembras e indeterminados y se establecieron siete intervalos de clases.
Sexo y estado de maduración gonadal
Para el análisis de las muestras se siguió la metodología recomendada por Camacho (2011) y Lucano-Ramirez et al. (2011), que consiste en hacer la disección de los ejemplares y retirar las gónadas. Se identificó el sexo y el estado de madurez gonadal a nivel macroscópico, de acuerdo con el método propuesto por Vegas (1987), en el que se distinguen los machos y las hembras en cuatro estadios de desarrollo gonadal. Luego se tomó el peso de las gónadas y se preservaron en formol neutralizado al 10 % para su posterior procesamiento histológico.
Para la identificación de las fases reproductivas se extrajo la gónada derecha del pez, se utilizó una porción de la parte anterior de la muestra y se le realizó un corte transversal. Posteriormente estos cortes se introdujeron en rejillas histológicas para su procesamiento. Para la fase histológica se identificaron las fases reproductivas de las gónadas a partir de fotografías y para la descripción se siguieron los criterios recomendados por Brown-Peterson et al. (2011).
Para la estimación de la fecundidad se extrajo una submuestra de la gónada izquierda y se realizó un corte transversal, el cual se estandarizó para todos los ejemplares a una porción de 0,12 g. Se llevó a cabo siguiendo los métodos gravimétricos (Holden y Raitt, 1974) y de análisis de imágenes descritos por Kilbansky y Juanes (2008). Para la separación de los oocitos se utilizó NaCl al 1 % (Vieira et al., 1999) y se dejaron 30 min para su separación. Luego se lavaron con agua destilada y se utilizó un tamiz de 75 μιη donde estos se retuvieron y se colocaron sobre papel filtro. Posteriormente, se vaciaron sobre una placa Bogorov en la que se limpiaron y se separaron bajo un estereoscopio. Finalmente, se fotografiaron bajo microscopio para contarse y medirse con el programa Image J.
Se tomó la longitud total, el peso de los ejemplares, el peso de la gónada, el peso de la submuestra y el número y diámetro de los ovocitos. Se estimó la fecundidad parcial y la fecundidad relativa de cada hembra madura y se calculó el índice gonadosomático, relacionado con la madurez gonadal. Para la estimación del diámetro de los oocitos, se midió la longitud del diámetro mayor y menor de cada uno por medio del programa Image J y se empleó la ecuación propuesta por Camacho (2011).
La fecundidad parcial se estimó con la ecuación propuesta por Lucano-Ramirez et al. (2005):
No es el número de ovocitos en una muestra de la gónada, PG es el peso de la gónada y PM el peso de la muestra gonadal (0,12 g). La fecundidad relativa con respecto a la talla se obtuvo dividiendo la fecundidad parcial ya calculada por la LT del pez. Se realizaron análisis de correlación entre la fecundidad parcial con las variables LT, peso total y peso gonadal.
El índice gonadosomático (IGS) se estimó según lo propuesto por Rosenblum et al. (1987) y Rodríguez (1992):
Wg es el peso de la gónada y Wt es el peso total del ejemplar.
RESULTADOS
Especies acompañantes
Para las localidades estudiadas donde P. acuminatus estuvo presente, se observaron en total 53 especies de peces (Tabla 1). La riqueza de especies acompañantes fue baja en Taganga (cinco especies) y Granate (18 especies), seguida por Playa Blanca (21 especies) e Isla Aguja (22 especies), mientras que en el banco de las Ánimas se observaron 40 especies acompañantes. Las familias con mayor presencia fueron: Haemulidae (diez especies), Pomacentridae (ocho), Holocentridae (cuatro) y Chaetodontidae (cuatro). Las especies Myripristis jacobus Cuvier, 1829, Canthigaster rostrata (Bloch, 1786), Haemulon flavolineatum (Desmarest, 1823), Haemulon steindachneri (Jordan y Gilbert, 1882), Stegastes partitus (Poey, 1868) y Chromis multilineata (Guichenot, 1853) fueron comunes en todas las localidades.
Tabla 1 Especies acompañantes asociadas a Pareques acuminatus en los diferentes sitios de muestreo
| Familia | Especie | B. de las Ánimas | Playa Blanca | Isla Aguja | Granate | Taganga |
|---|---|---|---|---|---|---|
| Sciaenidae | Equetus punctatus (Bloch & Schneider, 1801) | x | x | |||
| Pareques acuminatus (Bloch & Schneider, 1801) | x | x | x | x | x | |
| Odontoscion dentex (Cuvier, 1830) | x | |||||
| Labridae | Bodianus rufus (Linnaeus, 1758) | x | x | x | x | |
| Halichoeres garnoti (Valenciennes, 1839) | x | x | ||||
| Thalassoma bifasciatum (Bloch, 1791) | x | x | x | |||
| Holocentridae | Myripristis jacobus Cuvier, 1829 | x | x | x | x | x |
| Holocentrus rufus (Walbaum, 1792) | x | |||||
| Holocentrus adscensionis (Osbeck, 1765) | x | x | x | |||
| Canthigaster rostrata (Bloch, 1786) | x | x | x | x | x | |
| Haemulidae | Anisotremus surinamensis (Bloch, 1791) | x | ||||
| A. virginicus (Linnaeus, 1758) | x | |||||
| Haemulon macrostomum Günther, 1859 | x | |||||
| H. aurolineatum Cuvier, 1830 | x | |||||
| H. flavolineatum (Gilbert, 1882) | x | x | x | x | x | |
| x | x | x | x | x | ||
| H. plumierii (Lacepède, 1801) | x | x | ||||
| H. melanurum (Linnaeus, 1758) | x | |||||
| H. sciurus (Shaw, 1803) | x | |||||
| Brachygenys chrysargyreum Günther, 1859 | x | x | x | |||
| Pomacentridae | Stegastes partitus (Poey, 1868) | x | x | x | ||
| S. adustus (Troschel, 1865) | x | |||||
| S. planifrons (Cuvier, 1830) | x | |||||
| Abudefduf saxatilis (Linnaeus, 1758) | x | |||||
| A. taurus (Müller & Troschel, 1848) | x | |||||
| Chromis multilineata (Guichenot, 1853) | x | x | x | |||
| C. cyanea (Poey, 1860) | x | x | ||||
| Microspathodon chrysurus (Cuvier, 1830) | x | |||||
| Priacanthidae | Heteropriacanthus cruentatus (Lacepède, 1801) | x | ||||
| Priacanthus arenatus Cuvier, 1829 | x | |||||
| Scaridae | Sparisoma aurofrenatum (Valenciennes, 1840) | x | x | |||
| Scarus vetula Bloch & Schneider, 1801 | x | x | x | |||
| Monacanthidae | Cantherhines macrocerus (Hollard, 1853) | x | x | |||
| C. pullus (Ranzani, 1842) | x | |||||
| Scorpaenidae | Pterois volitans (Linnaeus, 1758) | x | x | x | ||
| Pomacanthidae | Pomacanthus paru (Bloch, 1787) | x | x | x | x | |
| Holacanthus tricolor (Bloch, 1795) | x | |||||
| H. ciliaris (Linnaeus, 1758) | x | x | ||||
| Acanthuridae | Acanthurus tractus Poey, 1860 | x | x | x | ||
| A. coeruleus Bloch & Schneider, 1801 | x | x | ||||
| A. chirurgus (Bloch, 1787) | x | |||||
| Ostraciidae | Lactophrys triqueter (Linnaeus, 1758) | x | x | |||
| Serranidae | Cephalopholis cruentata (Lacepède, 1802) | x | x | x | ||
| Diplectrum sp | x | |||||
| Chaetodontidae | Chaetodon sedentarius Poey, 1860 | x | x | |||
| C. ocellatus Bloch, 1787 | x | |||||
| C. striatus Linnaeus, 1758 | x | |||||
| C. capistratus Linnaeus, 1758 | x | x | ||||
| Mullidae | Mulloidichthys martinicus (Cuvier, 1829) | x | x | |||
| Pseudupeneus maculatus (Bloch, 1793) | x | x | ||||
| Lutjanidae | Lutjanus apodus (Walbaum, 1792) | x | ||||
| L. synagris (Linnaeus, 1758) | x | |||||
| L. jocu (Bloch & Schneider, 1801) | x | |||||
| Muraenidae | Gymnothorax funebris Ranzani, 1839 | x | ||||
| Total Especies | 41 | 22 | 23 | 19 | 6 | |
Morfometría y Merística
De los 85 especímenes de P. acuminatus recolectados de acuerdo con la longitud estándar (LS), 73 fueron adultos y 12 juveniles. De los adultos, 59 ejemplares presentaron longitud estándar (LS) entre 102 y 154 mm, longitud total (LT) entre 116 y 176 mm y longitud horquilla (LH) entre 114 y 173 mm (Tabla 2). Los juveniles presentaron amplitudes de LS de 28 a 55 mm, de LT de 33 a 66 mm y de LH de 31 a 61 mm (Tabla 2).
Tabla 2 Estadística descriptiva y datos expresados en porcentajes de LS en adultos y juveniles de P. acuminatus (mm). Longitud de las aletas pélvicas (L.A.Pe), Longitud de la cabeza (L.C), Diámetro del ojo (D.O), Longitud interorbital (L.I.O), Altura del cuerpo (A.C), Altura del pedúnculo caudal (A.P.C), Longitud del pedúnculo caudal (L.P.C), Longitud de la aleta pectoral (L.A.P), Longitud de la mandíbula (L.M) y Longitud del rostro (L.R).
| Morfometría | Este estudio (Adultos) | Cervigón (1991) | Juveniles | ||||
|---|---|---|---|---|---|---|---|
| Mínimo | Máximo | Promedio (LS) | Ámbitos (LS) | Mínimo | Máximo | Promedio (LS) | |
| L.A.Pe | 19,4 | 24,1 | 22,0 | 21,8 - 25,0 | 18,2 | 29,0 | 23,6 |
| L.C | 29,0 | 35,0 | 31,0 | 33,0 - 36,1 | 25,0 | 35,6 | 32,3 |
| D.O | 7,4 | 9,8 | 8,7 | 7,9 - 9,5 | 8,3 | 19,4 | 11,0 |
| L.I.O | 6,6 | 9,1 | 7,7 | 7,7 - 8,6 | 6,3 | 16,1 | 9,4 |
| A.C | 31,6 | 38,3 | 34,2 | 35,7 - 39,0 | 17,9 | 27,3 | 23,3 |
| A.P.C | 8,2 | 10,4 | 9,0 | 9,0 - 10,5 | 6,5 | 10,0 | 8,2 |
| L.P.C | 21,0 | 28,0 | 23,0 | 23,8 - 25,6 | 23,8 | 29,0 | 26,9 |
| L.A.P | 19,0 | 24,0 | 22,0 | 21,8 - 22,7 | 16,7 | 26,1 | 21,8 |
| L.M | 8,1 | 13,6 | 9,8 | 11,6 - 12,6 | 7,1 | 11,1 | 9,0 |
| L.R | 7,1 | 10,0 | 8,4 | 8,7 - 10,6 | 4,8 | 10,0 | 7,3 |
La merística se evaluó en los individuos adultos (n=73) y se registra en la Tabla 3. Sin embargo, con propósitos comparativos solo se consideron los cinco caracteres definidos por Cervigón (1991); se describe la frecuencia de los radios de la segunda aleta dorsal, la aleta anal, de la aleta pectoral, el número de branquiespinas de la rama superior y el de la rama inferior (Tabla 3). Para los juveniles se analizaron aquellos entre 28 y 55 mm de LS con los mismos caracteres medidos en los adultos (Tabla 3). Se observan ciertas diferencias menores en los valores modales de los contajes de la aleta dorsal y de branquiespinas, las cuales podrían indicar diferencias poblacionales que deberán ser exploradas en el futuro.
Tabla 3 Distribuciones de frecuencia de los caracteres merísticos en adultos y juveniles de la especie.
| Adultos | Juveniles | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carácter/Frecuencia | Especie | Cervigón (1991) | Frecuencia | Amplitud (mm) | Moda (mm) | |||||||||
| Amplitud (LS mm) | Moda (mm) | |||||||||||||
| Esp y Rad.A.Dor | 37 | 38 | 39 | 40 | 41 | X, 37-41 | 37 | IX-I, 37-41 | 37 | 38 | 39 | 40 | X, 37-40 | 38 |
| 32 | 23 | 8 | 7 | 3 | 1 | 7 | 3 | 1 | ||||||
| Esp y Rad.A.Anal | 7 | 8 | II, 7-8 | 7 | II, 7-8 | 7 | 8 | II, 7-8 | 7 | |||||
| Frecuencia | 56 | 17 | 11 | 1 | ||||||||||
| Rad.A. Pect. | 16 | 17 | 18 | 16-18 | 16 | 16-18 | 13 | 17 | 16-17 | 16 | ||||
| Frecuencia | 49 | 22 | 2 | 10 | 2 | |||||||||
| Bran. Sup. | 5 | 6 | 7 | 5-7 | 7 | 5-7 | 6 | 7 | 6-7 | 7 | ||||
| Frecuencia | 4 | 6 | 63 | 3 | 9 | |||||||||
| Bran. Inf. | 10 | 11 | 12 | 13 | 14 | 10-14 | 12 | 10-14 | 12 | 13 | 14 | 12-14 | 14 | |
| Frecuencia | 13 | 9 | 22 | 16 | 13 | 1 | 3 | 8 | ||||||
| n | 73 | 24 | ||||||||||||
Aspectos reproductivos
Para la determinación del sexo y del estado gonadal se observaron las gónadas de 73 individuos. Se encontraron 39 hembras en los distintos estados de desarrollo, con una mayor abundancia del estadio III de maduración (n = 17) y con variaciones en la talla entre 113 y 176 mm, con un promedio de 138,1 ± 15,8 mm. Para los machos se encontraron 34 individuos; el estadio II fue el más abundante (n = 17) y la talla varió entre 78 y 183 mm, con un promedio de 139,8 ± 26,1 mm. No fue posible identificar el sexo de 12 especímenes. Con respecto a la talla, se encontró un mayor número de hembras en tallas intermedias. Entre los seis indidividuos de mayor talla (165-186 mm) cuatro fueron machos y dos hembras y entre los individuos pequeños (77 98 mm) solo se encontraron machos.
Los ovarios de P. acuminatus se identificaron como dos lóbulos alargados, generalmente de igual tamaño, localizados a cada lado del cuerpo en la cavidad abdominal y recubiertos por una membrana que los separa de los demás órganos. El tamaño y la coloración cambiaron según los diferentes estados de madurez. El análisis histológico indicó que presentan desarrollo ovocitario asincrónico, dado que contienen ovocitos en distintas etapas de desarrollo. Se identificaron dos fases reproductivas en las hembras analizadas: fase en desarrollo y fase capaz de desove.
La fase en desarrollo correspondió a una hembra con la gónada en estado III de madurez. En este estado se observan ovarios lobulados, turgentes y opacos de coloración naranja claro, las gónadas son voluminosas y se presentan algunos vasos sanguíneos (Fig. 1a). Esta fase se caracteriza por la presencia de ovocitos con alveolos corticales (CA), ovocito vitelogénico primario (Vtg1), ovocito vitelogénico secundario (Vtg2) y ovocito de crecimiento primario (PG) (Fig. 1b).
En la fase capaz de desove se encontraron seis hembras con tallas entre 118 y 142 mm de LT. Estas correspondieron a hembras en estado IV de madurez, que se caracteriza por presentar ovarios lobulados, de coloración amarillenta a rojo, paredes rugosas y con vasos sanguíneos más prominentes; se observaron ovocitos en reabsorción (Figs. 1c y 1e). Los ovocitos se advierten a manera de grumos amarillos claros en la superficie ovárica (Fig. 1c).
El estadío IV se caracterizó por la presencia de ovocitos vitelogenados terciarios (Vtg3), alveolos corticales (CA) y ocurrencia de ovocitos en diferentes fases de crecimiento primario (PG) y secundario (nucléolo cromatina y con alveolos corticales) Se observan gránulos de vitelo (Gv) ubicados por todo el citoplasma entre los alvéolos corticales, los cuales aumentan progresivamente (Fig. 1d). También pueden verse ovocitos hidratados de forma irregular y de gran tamaño y con presencia de vesículas (Fig. 1f).
En las gónadas de las hembras maduras de P. acuminatus se percibieron oocitos en diferentes tamaños y estados de maduración, lo cual es característico de especies con desove parcial. Los ovocitos se observaron en hembras con estadío III y IV de madurez sexual, con LT entre 118 y 152 mm y un peso entre 26,06 y 58,89 g. El peso de la gónada osciló entre 1,10 y 2,64 g. El diámetro promedio de los ovocitos maduros fue de 0,53 ± 0,04 mm, siendo el más pequeño de 0,48 mm y el más grande de 0,59 mm (Tabla 4).
Tabla 4 Estimación de la fecundidad y del índice gonadosomático en P. acuminatus.
| Individuo | Peso Total (g) | LT (mm) | Peso de la gónada (g) | Peso de la muestra (g) | Número de oocitos | Diámetro de oocitos (mm) | Fecundidad parcial | Fecundidad relativa | Índice gonadoso-mático |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 42,87 | 142 | 2,08 | 0,12 | 698 | 0,58 | 12099 | 85,2 | 4,85 |
| 2 | 26,06 | 118 | 1,11 | 0,12 | 525 | 0,52 | 4856 | 41,15 | 4,26 |
| 3 | 32,71 | 128 | 1,1 | 0,12 | 488 | 0,48 | 4473 | 34,95 | 3,36 |
| 4 | 29 | 127 | 1,12 | 0,12 | 605 | 0,55 | 5647 | 44,46 | 3,86 |
| 5 | 58,89 | 152 | 1,78 | 0,12 | 838 | 0,5 | 12430 | 81,78 | 3,02 |
| 6 | 39,5 | 135 | 1,2 | 0,12 | 815 | 0,5 | 8150 | 60,37 | 3,04 |
| 7 | 46,38 | 134 | 2,64 | 0,12 | 616 | 0,59 | 13552 | 101,13 | 5,69 |
| Promedio | 39,34 | 133,71 | 1,58 | 0,12 | 655 | 0,53 | 8744 | 64,15 | 4,01 |
| Desv. Estándar | 11,34 | 11,03 | 0,61 | 0,00 | 135,25 | 0,04 | 3900,04 | 25,51 | 1,00 |
La fecundidad se determinó en siete hembras maduras, de las cuales cinco se colectaron en Taganga y dos en Playa Blanca. Estas se encontraron en estadios III y IV de madurez gonadal y su LT varió entre 118 y 152 mm. El peso de la gónada fluctuó entre 1,10 y 2,64 g. La fecundidad parcial promedio fue de 8744 ± 3900 ovocitos (Tabla 4). El mayor número de oocitos (13 552 ovocitos) se obtuvo en una hembra de 134 mm LT y 46,38 g, mientras que la menor fecundidad (4473) se registró en una hembra de 128 mm LT y 32,71 g. Individuos con diferentes tallas presentaron una fecundidad parcial diferente, y a su vez hembras con peso gonadal similar mostraron números de ovocitos distintos. La fecundidad relativa a la talla fue de 64,15 ± 25,51 oocitos/mm. El valor más alto fue de 101,13 oocitos/mm en una hembra de 134 mm y el más bajo de 34,95 oocitos/ mm en una hembra de 128 mm (Tabla 4).
El índice gonadosomático promedio fue de 4,01 ± 1,00. El valor más alto fue de 5,69 en una hembra de 134 mm LT con un peso de 46,38 g y el más bajo fue 3,02 en una hembra de 152 mm con un peso de 58,89 g (Tabla 4).
Las regresiones entre la fecundidad parcial y las variables LT, peso total y peso gonadal mostraron modelos lineales positivos. Entre la fecundidad parcial y la talla de las hembras maduras el coeficiente fue R2 = 0,61. Con respecto al peso total, el ajuste fue de R2 = 0,74. Con respecto al peso gonadal y a la fecundidad parcial, la correlación fue de R2 = 0,83 y entre el peso total y LT se encontró el mejor ajuste (R2 = 0,88).
DISCUSIÓN
Pareques acuminatus es un pez que habita en ambientes muy diversos, relacionados con fondos coralinos y sustratos rocosos y fangosos, en hábitats someros. Normalmente vive en grupos pequeños. Los juveniles se pueden observar sobre sustrato arenoso cerca de piedras y formaciones coralinas (Cervigón, 2012; Robertson et al., 2015). En los sitios estudiados, P. acuminatus se observó en fondos coralinos, debajo de las rocas, y en pequeños grupos acompañado de una variedad de especies de distintas familias. Por lo tanto, se confirma que esta especie puede vivir en diferentes ambientes, como sustratos rocosos y fondos duros coralinos, y es representativa de la ictiofauna de fondos arrecifales someros en la región de Santa Marta.
Con respecto a las especies con las que comparte el hábitat, para el Caribe se destacan las familias Haemulidae, Pomacentridae, Serranidae, Scaridae, Labridae, Lutjanidae y Chaetodontidae (Grijalba et al., 2004). En las localidades estudiadas las familias mejor representadas fueron Haemulidae, Pomacentridae, Holocentridae y Chaetodontidae; las especies acompañantes más comunes fueron M. jacobus, C. rostrata, H. flavolineatum y H. steindachneri; en segunda instancia están Bodianus rufus (Linnaeus, 1758) y Pomacanthus paru (Bloch, 1787) y en tercer lugar S. partitus y C. multilineata. Con excepción de B. rufus y P. paru, las especies acompañantes coinciden básicamente con lo registrado por Grijalba et al. (2004).
Myripristis jacobus fue la especie acompañante más representativa en todos los sitios de muestreo, mientras que S. partitus y C. multilineata estuvieron presentes en tres de cinco. La dominancia de este conjunto de taxones puede deberse a condiciones ecológicas de alimentación y refugio que parecen favorecer su permanencia (Grijalba et al., 2004). La estrecha relación de P. acuminatus con un elevado número de especies indica que no es una especie solitaria, sino que está asociada con una gran diversidad de peces arrecifales.
Teniendo en cuenta los datos registrados por Cervigón (1991) y los ejemplares examinados, se encontró que las mediciones morfométricas con menor amplitud fueron longitud de la cabeza, altura del cuerpo, longitud del pedúnculo caudal, longitud de la mandíbula y longitud del rostro. La longitud de la aleta pélvica, el diámetro del ojo, la longitud interorbital, la altura del pedúnculo caudal y la longitud de la aleta pectoral oscilaron en espectros más amplios de los registrados por Cervigón (1991). Aunque no se encontró una tendencia en los caracteres analizados en comparación de los datos registrados, se podría considerar que las diferencias podrían depender del observador y de pequeñas divergencias metodológicas.
Los caracteres merísticos cuantificados para los adultos coinciden en gran parte con los registrados por Cervigón (1991) y Smith (1997). La única diferencia con Smith (1997) fue para los radios en la aleta dorsal (37-40), mientras en este estudio se encontró entre 37 y 41. Los datos con Cervigón (1991) fueron todos coincidentes. Para los juveniles, la merística mostró mayor número de branquiespinas en comparación con los adultos. Esto indica que a medida que va creciendo el individuo éstas se van reduciendo en número, lo cual puede deberse a variables relacionadas con la alimentación y con el aparato branquial, como el tamaño de la cabeza y el de la boca. En los juveniles la longitud de la cabeza tiende a ser más grande que en los individuos adultos (Barón, 2006; González et al., 2011). De esta manera, los caracteres merísticos y morfométricos permitieron verificar que los individuos de P. acuminatus recolectados en Santa Marta no difieren de los de otras localidades del Caribe, y probablemente se trata de una sola población, al menos en el sur del Caribe.
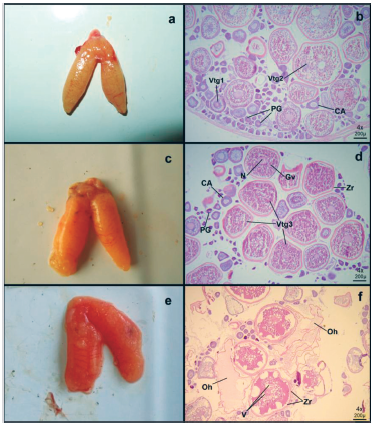
Figura 1 a) Fotografía de ovarios de ejemplar de 152 mm LT. b) Microfotografìa (4x) de ovario. CA: ovocitos con alveolos corticales. Vtgl: ovocito vitelogénico primario. Vtg2: ovocito vitelogénico secundario. PG: ovocito de crecimiento primario. c) Fotografía de ovarios de ejemplar de 135 mm LT y d) Microfotografìa (4x) de ovario. Vtg3: ovocito vitelogénico terciario. PG: ovocitos de crecimiento primario. CA: alveolos corticales. Gv: gránulos de vitelo, N: núcleo, Zr: zona radiada. e) Fotografía de ovarios de ejemplar de 134 mm LT. f) Microfotografía de ovocitos en fase capaz de desove en vista (4x). Zr: zona radiada, Oh: ovocitos hidratados, V: vesículas.
En nuestro estudio se halló una mayor proporción de hembras de P. acuminatus que de machos. Al respecto y según Saiz (1975) y Escobar (1994), es frecuente en peces de la familia Sciaenidae encontrar una mayor abundancia de hembras, lo que reflejaría factores genéticos hereditarios, variaciones en la disponibilidad de alimento y agrupamientos para conservar la especie y asegurar la fecundidad solamente con algunos machos. Igualmente, las hembras parecen tener una alta tasa de crecimiento y una mayor esperanza de vida que los machos (Cardoso et al., 2018). Escobar (1994), Torres et al. (1999), Cancio et al. (2006) y Cardoso et al. (2018) encontraron igualmente en Micropogonias furnieri (Desmarest, 1823), Stellifer venezuelae (Schultz, 1945), Bairdiella ronchus (Cuvier, 1830) y Macrodon ancylodon (Bloch & Schneider, 1801) una proporción de hembras mayor que la de machos.
El desarrollo gonadal mostró cambios morfológicos relacionados con la ovogénesis, con un desarrollo ovárico en diferentes fases y etapas de desarrollo. La mayoría de las muestras correspondieron a hembras en estadío IV, las cuales presentaron una capacidad total para el desove. Según Eraso et al. (2017), en este estadío las gónadas ya han alcanzado su tamaño máximo, con los vasos sanguíneos mucho más visibles con respecto al estado anterior. Lowe (1987), Castillo (1998) y Lucano-Ramírez et al. (2011) describen en peces de aguas tropicales que el desove está principalmente influenciado por condiciones como las temperaturas apropiadas y la disponibilidad de alimento. Sin embargo, para la región de Santa Marta se encontraron hembras capaces de desovar todo el año.
La descripción histológica de las fases (en desarrollo y capaz de desove) y las etapas de desarrollo (oocitos vitelogénicos, alvéolos corticales y crecimiento primario) coinciden con las características mencionadas por Silva et al. (2016) para la especie Cynoscion leiarchus (Cuvier, 1830). Estos autores especifican cinco etapas de desarrollo de ovocitos y un desarrollo ovocitario de tipo asincrónico con fecundidad parcial. Se confirma este tipo de reproducción para P. acuminatus, en la que se pueden encontrar en los ovarios diferentes etapas de desarrollo antes y después del desove (Holden y Raitt, 1974). Estas características se han observado en las especies de la familia Sciaenidae mencionadas anteriormente; para M. furnieri, S. venezuelae, C. leiarchus y M. ancylodon, Escobar (1994), Silva et al. (2016) y Cardoso et al. (2018) describen una reproducción parcial y un desarrollo completamente asincrónico. Winemiller y Layman (2005) indican que este patrón de comportamiento puede considerarse como una respuesta adaptativa a las condiciones ambientales. Por lo tanto, estos rasgos reproductivos confirman que aquellas especies que están estrechamente relacionadas desarrollan estrategias reproductivas similares, con lo cual evitan la competencia intraespecífica por recursos limitados y superan las variaciones ambientales (Wootton, 1992; Amarasekare, 2003).
La estimación del diámetro promedio de los ovocitos maduros en P. acuminatus (0,53 ± 0,04 mm) fue mayor a los 0,45 mm registrados por Lasso et al. (1998) en Plagioscion squamosissimus (Heckel, 1840), a pesar de que esta especie de esciénido es de talla mayor. La fecundidad parcial fue de 8744 ± 3900 ovocitos, lo cual se aproxima a lo estimado por Flores y Hirt (2002) para Pachyurus bonariensis (Steindachner 1879), una especie de esciénido de talla pequeña; dichos autores encontraron 11 000 ovocitos en esta especie. Con respecto a este parámetro, la fecundidad parcial no es solamente el número de huevos o de ovocitos que una hembra podría producir después del desove, sino también es una medida de la capacidad reproductiva de los individuos (Vazzoler, 1996).
Muchos factores pueden afectar la fecundidad de los peces, tales como las condiciones ambientales y la falta de alimento y refugio (Csirke, 1980; Murua et al., 2003). Los cambios que se presentan en la fecundidad están estrechamente relacionados con el tamaño de los individuos o con la edad (Murua et al., 2006; Cooper et al., 2013). Esta tendencia no se evidenció para las hembras analizadas, ya que no todas aquellas de mayor peso y talla produjeron una mayor cantidad de ovocitos. Kjesbu et al. (1991) mencionan que las hembras que se encuentran en mejores condiciones son las que pueden presentar una mayor fecundidad. Finalmente, el peso gonadal indicó una correlación importante con respecto a la fecundidad parcial (R2 = 0,83). Silva et al. (2016) encontraron en el esciénido Cynoscion leiarchus un R2 de 0,67 para esta variable y mencionan que el peso de la gónada es el mejor predictor de la fecundidad, comparado con otras medidas como la LT, el peso del individuo o la edad. La relación entre el peso total y la LT mostró un mejor ajuste positivo para las hembras maduras. Según Ricker (1975), las relaciones alométricamente positivas pueden deberse a las condiciones nutricionales y a algunas diferencias entre poblaciones de la misma especie o dentro de la misma población.

