











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
INTRODUCCIÓN
La Reserva Nacional de Salinas y Aguada Blanca (RNSAB) hace parte de la Puna, ecorregión altoandina del Perú, ubicada en el centro de la cordillera de los Andes (Brack y Mendiola, 2000; Cuesta et al., 2012). Esta ecorregión presente en Arequipa está representada por ecosistemas en recuperación como pajonales, tólares, yaretales y relictos de bosques de Polylepis Ruiz & Pav. (queñuales) (Zeballos et al., 2010). Estos últimos son ecosistemas altoandinos con flora y fauna única, de gran interés ecológico, sistemático, biogeográfico, y con altos niveles de endemismo. Lamentablemente, son también ecosistemas vulnerables a causa de la fuerte presión antropogénica, por ser el único recurso de madera en zonas de altura (Servat et al., 2002).
Las abejas constituyen el grupo más grande de la superfamilia Apoidea. En la actualidad, se estima que existen más de 20 000 especies de abejas descritas en el mundo (Michener, 2007; Ascher y Pickering, 2020) y por lo menos 5000 especies para la Región Neotropical (Fernández y Sharkey, 2006). Las abejas son reconocidas por el importante papel que cumplen en la polinización de vegetación natural. Este servicio ecosistémico brinda diferentes beneficios que contribuyen en la agricultura, la economía y los medios de vida de la población (Michener, 2007).
Sumado a la importancia de las abejas en estos ecosistemas, en el Perú, los estudios de inventarios, diversidad y distribución de abejas nativas son escasos. Además, las zonas altoandinas entre los 3900-4230 m. s. n. m. donde se realizó el estudio, son las menos estudiadas y no hay información sobre las especies de abejas. Esto en parte se debe a la dificultad de acceso a estos lugares y a las condiciones climáticas adversas, como bajas temperaturas, alta precipitación, viento y nubosidad una gran parte del tiempo, lo cual dificulta los muestreos y observaciones de abejas. A pesar de esto, se hace necesario realizar inventarios y estudios ecológicos para conocer las comunidades biológicas de abejas que habitan en estas regiones, las cuales se encuentran amenazadas por actividades antrópicas como son la extracción de leña, conversión en carbón, quema de pastos, sobrepastoreo y ampliación de la frontera agrícola (Zeballos et al., 2010).
Por lo anterior, el propósito de este estudio fue conocer la diversidad y abundancia de especies de abejas en tres zonas representativas de ecosistemas altoandinos de la Puna en Arequipa, Perú.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en la zona de amortiguamiento de la RNSAB, en la localidad de El Simbral, distrito de Chiguata, aproximadamente a 25 km de la ciudad de Arequipa, Perú (Fig. 1). De acuerdo con la clasificación de Morrone (2001), se encuentra ubicada en la Región andina, subregión Paramo Puneña, Provincia de la Puna y se encuentra localizada a 16° 23' S y 71° 19' W. Presenta una altitud de 3900 a 4230 m. s. n. m.; la temperatura promedio anual es de 13,1 °C, mientras que la precipitación total anual es de 181,8 mm (Montenegro et al., 2010).
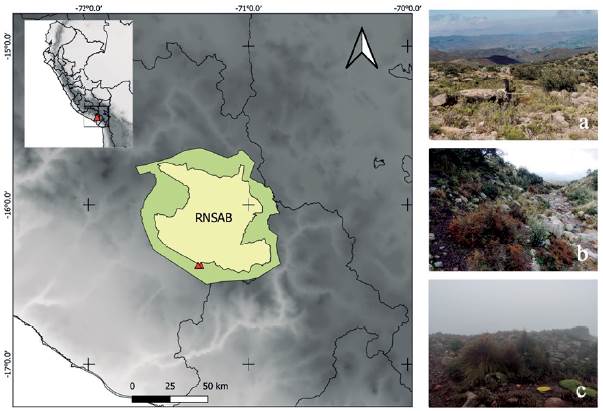
Figura 1: Zonas de estudio: a. Zona 1, b. Zona 2 y c. Zona 3; Zoom de las zonas en el mapa político del Perú-Arequipa. Estas zonas están ubicadas en la zona de transición de la Reserva Nacional de Salinas y Aguada Blanca (RNSAB), en la Provincia de Arequipa, departamento de Arequipa en el Perú.
La vegetación predominante es el resultado de asociaciones de tólares y gramíneas con queñuales (Polylepis), y de plantas y cactáceas almohadilladas en relieves accidentados a colinosos (Zeballos et al., 2010). El Perú es el país con mayor riqueza y endemismo para el género Polylepis con 19 especies (Mendoza y Cano, 2011) de un total de aproximadamente 27-28 especies en ecosistemas altoandinos (Simpson, 1979; Mendoza y Cano, 2010; Segovia-Salcedo et al., 2018). Polylepis pertenece a la familia Rosaceae; es el único género arbóreo presente entre los 3000-5000 m. s. n. m., se caracteriza por tener una altura de 1 a 27 m de alto, por tener fruto seco, con tallos y ramas retorcidas; y ritidoma marrón rojizo que se desprende en pedazos grandes, actualmente se encuentra bien distribuido en el departamento de Arequipa protegido en la Reserva Nacional de Salinas y Aguada Blanca (RNSAB), y en la Reserva Paisajística de la subcuenca del Cotahuasi (Simpson, 1979; Mendoza et al., 2010). Los yaretales por su parte pertenecen a la familia Apiaceae (subfamilia Azorelloideae) y son una asociación arbustiva almohadillada de plantas resinosas como: yaretas (Azorella compacta Phil.) y las acompañantes tales como Pycnophyllum J.Rémy, Nototriche Turcz., Mniodes (A.Gray) Benth. y Werneria Kunth; se caracterizan por alcanzar hasta un metro de altura y tener crecimiento muy lento (aproximadamente 1 mm/ año) y se encuentran formando parches de vegetación con cúmulos de plantas arrosetadas y subarbustos postrados, en las partes más altas de los andes, entre los 3800-4800 m. s. n. m. (Quipuscoa y Huamantupa, 2010; Zeballos et al., 2010; Talavera et al., 2011; Calviño et al., 2016).
En la región andina, a medida que la altitud aumenta va disminuyendo la presión atmosférica y la temperatura; y a su vez aumenta la humedad y la evapotranspiración. Estas condiciones son muy variables presentándose años secos o lluviosos, con sequias, heladas, granizadas o hasta excesos de lluvia. Por otro lado, con la altitud aumenta la insolación y la influencia enfriadora de vientos, lo que genera condiciones adversas que dificultan la vida en general (Montenegro et al., 2010).
Para los sitios de muestreo se ubicaron tres zonas representativas de los ecosistemas Polylepis-Yaretal presentes en el lugar de estudio (Fig. 1). En cada zona se delimitaron transectos de 2400 m2 a 2500 m2 (50 m x 50 m), con un área total de 7400 m2 aproximadamente y con las siguientes características:
Zona 1: 16° 23' S y 71° 19' W a 3900-4090 m. s. n. m. Delimitada por la carretera de Arequipa-Ubinas. Se caracteriza por ser una ladera montañosa que presenta fragmentos boscosos de un relicto de bosque de Polylepis.
Este relicto está representado por una gran densidad de árboles de queñua (Polylepis rugulosa Bitter) y flora herbácea como Senecio collinus DC, Parastrephia, Chuquiraga rotundifolia Weddell, Lupinus, Adesmia spinosissima Meyen ex J. Vogel, Asteráceas y Poáceas (Fig. 1a).
Zona 2: 16° 23' S y 71° 19' W a 3850-4060 m. s. n. m. Esta zona se caracteriza por presentar una quebrada seca y estrecha dentro de un relicto de bosque de Polylepis (Fig. 1b). En época húmeda es atravesada por pequeños riachuelos y presenta vegetación más herbácea con pocos arbustos, de las familias Asteráceas, Poáceas y Fabácea.
Zona 3. 16° 23' S y 71° 18' W a 4100-4230 m. s. n. m. Esta zona se caracteriza por presentar una transición entre un bosque de Polylepis y el ecosistema Yaretal. En esta zona encontramos plantas representativas como Pycnophyllum molle Remy, Werneria paposa Philippi, Nototriche obcuneata (Baker f.) AW Hill, Calamagrostis curvula (Wedd) Pilg., Belloa longifolia, Baccharis buxifolia (Lam.) Pers. y el género Azorella (INRENA, 2007). Al ser la zona más alta, se encuentra con neblina durante gran parte del día (Fig. 1c), con presencia de lluvia y granizos en época húmeda; mientras que, en época seca, presenta gran radiación y viento intenso a lo largo del día.
MATERIALES Y MÉTODOS
Colecta y muestreos
Los muestreos se realizaron mensualmente entre agosto de 2018 y febrero de 2019. La colecta de las abejas se realizó entre las 9:00 y las 13:00 horas con dos métodos de muestreo:
Red entomológica: Los transectos se recorrieron durante cuatro horas de muestreo sin pausas (9:00-13:00) por un colector por un total de 24 días de campo; las abejas se colectaron en las zonas de estudio mientras forrajeaban, descansaban en flores o en vuelo y se sacrificaron en acetato de etilo, siguiendo la metodología descrita por Sakagami et al. (1967).
Platos amarillos: Se colocaron recipientes de polietileno amarillo de alrededor de 40 x 25 x 3 cm, con solución de agua y jabón a mitad de su capacidad con un periodo de actividad de cuatro horas (9:00-13:00), durante 24 días de campo, según la metodología de González et al. (2016). Se eligió únicamente este color porque en el área de estudio, la mayoría de plantas presenta flores amarillas (Asteráceas) y en observaciones preliminares el color fue efectivo para la captura de abejas. Para cada transecto se instalaron nueve platos y los especímenes se conservaron en alcohol al 70 %.
Todos los especímenes colectados se montaron en seco y se identificaron con el uso de claves taxonómicas (Michener, 2000; Silveira et al., 2002; Fernández y Sharkey, 2006; Michener, 2007) y con la ayuda de expertos. El material entomológico (especímenes) se trasladó y depositó en la colección del Museo de Historia Natural de la Universidad Nacional de San Agustín (MUSA), código MUSA-ENT-HYM 00001-00545.
ANÁLISIS DE DATOS
Para analizar la riqueza estimada de las especies colectadas con respecto a la existente en la zona, se emplearon dos estimadores no paramétricos, Chao 1 y Bootstrap, y una curva de acumulación de especies usando el programa EstimateS versión 9.1.0 (Colwell, 2013). Se usó el programa PAST versión 3.23 (Hammer et al., 2001) para calcular los índices de diversidad, riqueza y similitud en las tres zonas estudiadas; se utilizaron los siguientes índices: Shannon-Wiener (H'), Simpson (D). Pielou (J') y Jaccard. También se analizó y se comparó la diversidad y las diferencias significativas entre las áreas estudiadas mediante la prueba T-student, la cual evaluó las varianzas de los valores de diversidad de Shannon-Wiener en dos muestras, con un nivel de significancia de 5 % (p < 0,05) (Hutcheson, 1970; Magurran, 1988).
También se analizaron agrupamientos a partir de la matriz de similitud de los coeficientes de Jaccard, construyendo un dendograma con el método de UPGMA (Unweighted Pair Group Method using Arithmetic Averages) (Romesburg, 1984) y realizado por el Software PAST versión 3.23 (Hammer et al., 2001).
RESULTADOS
En total se colectaron 545 especímenes, diez géneros y 26 morfoespecies; agrupados en cuatro familias de abejas: Apidae, Megachilidae, Halictidae y Colletidae (Tabla 1). La familia Halictidae presentó la mayor abundancia de individuos 475 (87,1 %), seguida de Megachilidae con 43 individuos (7,7 %), Colletidae con 21 individuos (3,8 %) y Apidae con seis individuos (1 %). El género más abundante y diverso fue Caenohalictus con ocho morfoespecies y con un total de 466 individuos (85,5 % del total de individuos colectados) (Fig. 2). Solamente cuatro morfoespecies se representaron por un solo individuo: Bombus funebris (Smith, 1854), (Zona1), Caenohalictus sp3, Caenohalictus sp6 y Acanthopus sp. (Zona 2).
Tabla 1 Número de individuos colectados por género de abejas con el uso dos métodos de colecta. En paréntesis número de morfoespecies colectadas por cada género.
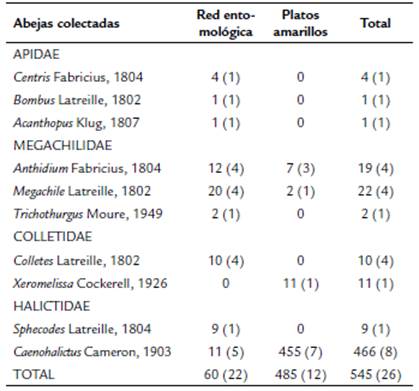
En cuanto a la eficiencia de los métodos de colecta, el método de platos amarillos fue más efectivo para la captura de individuos (485 individuos - 89 %) que el de red entomológica (60 individuos - 11 %) (Tabla 1). Para riqueza de especies, el método más efectivo fue el de red entomológica con 21 morfoespecies (81 %) y platos amarillos con 13 morfoespecies (50 %). Las especies exclusivas para el método de red entomológica fueron: Centris sp., Anthidium sp1, B. funebris, Acanthopus sp., Trichothurgus sp., Megachile sp1, Megachile sp2, Megachile sp3, Colletes sp1, Colletes sp2, Colletes sp3, Colletes sp4 y Caenohalictus sp3; y para platos amarillos fue: Xeromelissa sp., Caenohalictus sp6, Caenohalictus sp7 y Caenohalictus sp8.
En relación a la comparación entre zonas, las que presentaron mayor riqueza de especies (S) fueron las zonas 1 y 2 con 18 morfoespecies (Tabla 2). Los resultados obtenidos para la diversidad en las zonas de estudio muestran que el índice de Shannon-Wiener fue mayor en la zona 2 (H' = 2,048) (Tabla 2). Los valores obtenidos a partir del índice de Shannon para cada zona fueron concordantes con los valores dados por el test de Hutchenson, el cual mostró diferencias significativas entre las dos zonas 1 y 2, con un valor de 2,0102e-05 < 0.05, que indica que la diversidad de la comunidad de abejas en estas dos zonas es diferente. La comparación entre los grupos de las zonas 1/ zona 2 (0,084003), y zona 2/ zona 3 (0,16393) no presentó diferencias significativas.
Tabla 2 Número de individuos (N), riqueza de especies (S) y el índice de diversidad de Shannon (H'), índice de dominancia de Simpson (D) e índice equidad de Pielou (J') entre las zonas de estudio:
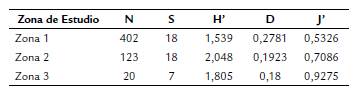
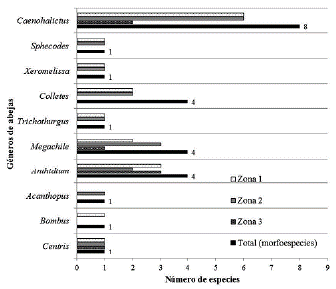
Figura 2 Géneros de abejas presentes en cada zona de estudio y número total de morfoespecies de abejas encontradas por cada género. Familia Apidae: Centris Fabricius, Bombus Latreille y Acanthopus Klug; Familia Megachilidae: Anthidium Fabricius, Megachile Latreille y Trichothurgus Moure; Familia Colletidae: Colletes Latreille y Xeromelissa Cockerell; y Familia Halictidae: Sphecodes Latreille y Caenohalictus Cameron.
Los valores generados por el índice de dominancia D de Simpson fueron bajos para las tres zonas de estudio, oscilando entre 0,18 (zona 3) y 0,27 (zona 1). Estos valores de dominancia se ven reflejados en la equidad según el índice de Pielou, cuyos valores en la zona 3 (J' = 0,9275) fueron más elevados en comparación con las otras dos zonas. Estos resultados indican que la zona 1 presentó mayor dominancia de especies, mientras que la zona 3 mayor equidad (Tabla 2).
La similitud en la composición de las especies obtenidas a partir del índice de Jaccard y del dendograma generado, mostraron que las zonas 1 y 2 tuvieron una similitud de 0,44, zona 1 y 3 (0,32), y zona 2 y 3 (0,25) (Fig. 3); estos valores indican que las zonas 1 y 2 tuvieron más especies compartidas en comparación con la zona 3.
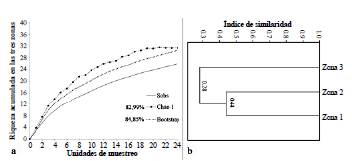
Figura 3 a. Curva de acumulación de especies y eficiencia para cada estimador. b. Dendograma de similitud entre las zonas de muestreo a partir de la presencia y ausencia de morfoespecies de abejas, con datos del índice de Jaccard. El coeficiente de correlación para este dendograma fue de 0,9401.
Se analizó la curva de acumulación de especies de abejas colectadas en las tres zonas estudiadas durante siete meses de muestreo, y se observó que su eficiencia, para cada sitio, fue sobre el 80 %, usando como estimadores Chao1 = 31,33 especies y Bootstrap = 30,64 especies (Fig. 3). Lo que indica que en esas zonas faltaron entre cuatro y seis especies por capturar, en relación al número de especies observadas = 26 (Sobs). Por lo que se sugiere que el esfuerzo de muestreo se acercó a la riqueza presente en esta provincia y que es necesario continuar haciendo estudios en la zona.
DISCUSIÓN
Los estudios de abejas en las regiones altoandinas son escasos, la mayoría de inventarios y análisis de diversidad de abejas en esta región han sido reportados para Colombia (González y Engel, 2004; González et al., 2005), Chile (Montalva et al., 2012; Henríquez-Piskulich et al., 2020) y Argentina (Medan et al., 2002). En el Perú los estudios de abejas, en general, son aún más escasos. Gran parte de estos trabajos han sido enfocados en actividades pecuarias como apicultura y meliponicultura (Chayña y Wily, 2019; Delgado et al., 2019; Delgado y Rasmussen, 2019). Los otros grupos de abejas son poco conocidas, y los escasos reportes que existen se refieren a descripciones de nuevas especies a partir de la revisión de material depositado en colecciones científicas (Vieira y Vivallo, 2009; Engel, 2013; Engel y Rasmussen, 2013; González, et al., 2013; González et al., 2019). En la actualidad no existen estudios de inventarios y diversidad de abejas en ecosistemas altoandinos en Perú; siendo únicamente mencionadas en levantamientos de artropofauna como es el caso de Oroz-Ramos et al. (2017) y Oroz-Ramos et al. (2018) en bosques de Polylepis, donde se reportan tres individuos de la Superfamilia Apoidea, sin llegar a identificarlas. Por lo anterior, el presente trabajo corresponde al primer estudio sobre la diversidad de abejas para ecosistemas altoandinos en Perú, incluyendo un inventario preliminar que permite obtener información sobre las especies de abejas con distribución sobre los 4000 m. s. n. m. Dichos inventarios son una herramienta para los planes de conservación y uso de recursos naturales.
Las familias mejor representadas (número de especies y abundancia relativa) fueron Colletidae, Megachilidae y Halictidae. La familia Andrenidae no se reporta en este estudio, por ser una familia muy escasa y rara para Sudamérica y la región Neotropical. Según Moure et al. (2007), solamente cinco especies de esta familia han sido reportadas para el Perú.
Para este estudio se identificaron un total de 26 morfoespecies (diez géneros). Caenohalictus fue el género que presentó mayor abundancia (85 % del total de individuos colectados) y riqueza (ocho morfoespecies de un total de 26) (Fig. 2). Este género presenta amplia distribución en el Neotrópico y principalmente en los Andes de Sudamérica con siete especies presentes para Perú y especies nuevas por describir (González et al., 2005; Moure et al., 2007). Asimismo, Michener (2007), menciona que la tribu Halictini es más diversa y abundante en los Andes y que su época de reproducción es en verano, lo cual coincide con las fechas en que se realizaron los muestreos en este estudio y donde se colectaron en su mayoría halíctidos.
Para el género Anthidium se colectaron cuatro morfoespecies asociadas a plantas de las familias Asteraceae y Fabaceae (S. collinus, Parastrephia sp, C. rotundifolia y A. spinosissima).González y Griswold (2013) reportan para la región de Puna en Perú, especies de este género: Anthidium adelphumGonzález y Griswold (2013), A. alsinai Urban 2001, A. deceptum Smith 1879, A. funereum Schletterer 1890, A. masunariae Urban 2001, A. nigerrimum Schrottky 1910, A. peruvianum Schrottky 1910, A. toro Urban 2001; indicando que también visitan flores de Asteraceae y Fabaceae. Adicionalmente, se colectaron cuatro morfoespecies del género Megachile. De acuerdo con Durante et al. (2006), en Perú se ha reportado a Megachile garleppi Friese 1994, M. piurensis Cockerell 1911 y M. saulcyi Guérin 1844; está última descrita para la región Andina en países como Bolivia, Argentina, Chile y Perú.
La mayoría de los especímenes no se lograron identificar a nivel de especie debido a la escasa información y pocos reportes que se tienen sobre las especies de abejas para el Perú y zonas altoandinas sobre los 3900 m. s. n. m. Tampoco existen colecciones de referencia y claves taxonómicas de abejas en el país. Únicamente se identificó a nivel de especie, un macho del género Bombus: B. funebris, encontrado en la zona 1, entre los 3900-4090 m.s.n.m. Según Rasmussen (2003), esta especie se encuentra distribuida en el Neotrópico desde Colombia hasta Chile y es reportada para la región de Arequipa. Liévano y Ospina (1984), indican que esta especie alcanza los 4750 m.s.n.m. para Colombia.
Respecto a las especies parásitas, se colectó una morfoespecie de Sphecodes. Según Michener (2007), este género parasita en su mayoría a especies de la familia Halictidae, y también pueden atacar especies de la familia Colletidae y Andrenidae. Asimismo, se encontró una morfoespecie cleptoparásita del género Acanthopus.Gaglianone (2001) señala que este género ataca individuos del género Centris, y probablemente está relacionado con especies de abejas encontradas en el lugar.
El horario escogido para la colecta (9:00 a 13:00) fue seleccionado ya que se observó mayor actividad de abejas mejores condiciones climáticas (radiación solar, fuerza de viento, humedad, etc.) en este horario; en horarios anteriores y posteriores a estos, no se observó actividad de abejas por las condiciones climáticas adversas, sobretodo la temperatura y humedad. El método de platos amarillos fue más fácil de emplear en las zonas 1 y 3 debido que estas presentaban un terreno más abierto en comparación con la zona 2 que presentaba un terreno con laderas abruptas y áreas rocosas y estrechas. Mientras que el método de red entomológica fue difícil de emplear en las tres zonas de estudio debido a la presencia de A. spinosissima de la familia Fabácea, la cual es una especie de arbusto con espinas muy ramificado (Quipuscoa y Huamantupa, 2010), que dificultó el desplazamiento y el esfuerzo al colectar.
El método de platos amarillos proporcionó abundancias relativas más altas que casi en su totalidad eran abejas de menor tamaño (Sphecodes, Caenohalictus, Xeromelissa y Colletes) (Tabla 1). González et al. (2016) indica que las abejas más grandes se capturan con mayor frecuencia con trampas elevadas que con trampas a nivel del suelo; asímismo, los platos amarillos son un método de muestreo que requiere menos horas de trabajo y no depende de colectores capacitados, lo cual elimina el sesgo y es más completa para la evaluación de riqueza mas no tanto para abundancia relativa (Wilson et al., 2008). El método de red entomológica proporcionó menor abundancia relativa en comparación con otros estudios de levantamiento de abejas por el mismo hecho de ser un ecosistema altoandino con menor disponibilidad de recursos, por el terreno accidentado (terreno rocoso y depresiones) que dificultaba el desplazamiento normal, y por el mayor esfuerzo al colectar en bajas temperaturas y en condiciones climáticas adversas como se explicó anteriormente, típicas de los Andes, a grandes altitudes (4000 m. s. n. m.).
CONCLUSIONES
Los ecosistemas altoandinos de Yaretal y de relicto de bosque de Polylepis, presentes en la Puna, a pesar de las condiciones climáticas y geográficas adversas pueden llegar a presentar una considerable riqueza de especies de abejas como se puede observar con los resultados obtenidos en este estudio. Estas especies de abejas están relacionadas con la gran riqueza de flora endémica presente en estos ecosistemas altoandinos. Las abejas nativas a su vez requieren ser más estudiadas para poder conocer mejor la composición de la comunidad e incrementar los esfuerzos de muestreo con el fin de tener una idea real de las especies presentes en esta zona y en otras regiones altoandinas. De esta manera es posible diseñar planes de manejo en su conservación y ampliar el conocimiento sobre la fauna de abejas de Perú.

