











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El ébano (Caesalpinia ebano H. Karst.) es una especie forestal de la familia Fabaceae, nativa del Bosque seco tropical (bs-T) de Colombia; tiene un alto potencial en la silvicultura y en la ingeniería ecológica, por su buena cobertura arbórea. El bioma bs-T es uno de los ecosistemas más degradados, se encuentra distribuido en fragmentos aislados de diversos tamaños, el cual representa solo un 8 % de su cobertura original en el país (García et al., 2014; Bocanegra-González et al., 2019). Como consecuencia de la disminución del área forestal en el Caribe colombiano, C. ebano se ha reportado como una especie silvestre en peligro (EN A2cd), protegida de la tala con finalidad extractiva de madera. Para la zona, también se han reportado especies forestales EN, como guayacán (Bulseniaarbórea [Jacq.] Engl.) y ceiba tolúa (Bombacopsis quinata [Jacq.] Dugand.) (López y Cárdenas, 2002).
Dadas las pocas poblaciones de ébano en Colombia, el estudio de la población base y su variabilidad genética comprende una de las principales etapas para guiar posteriores estudios. La investigación en torno a esta especie es escasa, sobre todo la relacionada con las actividades del mejoramiento genético forestal. Autores como Bernal et al. (2017), sugieren estudios ecológicos y de poblaciones particularmente para ébano y carreto (Aspidosperma polyneuron Müll. Arg.), así como el emprendimiento de acciones para su conservación, debido a su valor ecológico, bienes y servicios ambientales, y socioeconómicos que representan.
Los parámetros genéticos de una población objeto de mejoramiento, están en función de los efectos ambientales que influyeron sobre dicha población al momento de tomar los valores fenotípicos de las respectivas variables poligénicas (Falconer y Mackay, 1996), y se fundamentan principalmente en la estimación de la variabilidad fenotípica y genética. Estos valores, asociados con los coeficientes de correlación entre caracteres, son importantes en los programas de mejoramiento porque permiten conocer la variabilidad genética, el grado de expresión de una generación a otra, posibilitan el incremento de características cuantitativas mediante la selección directa o indirecta y sirven de guías para seleccionar los rasgos determinantes de los caracteres poligénicos (Hallauer et al., 2010).
La predicción del valor genético depende de la estimación de los componentes de varianza (Abreu et al., 2017). Para tal fin, se considera el coeficiente de variación genético, la heredabilidad en sentido amplio y estricto, el avance y la ganancia genética (Cruz et al., 2012; Jamoza et al., 2014).
La heredabilidad mide la proporción de la variación fenotípica en una población que se debe a factores genéticos. Los avances genéticos derivan tanto del control genético de las características de interés como de la variabilidad genética existente en la población y regulan la mejora de un carácter bajo una presión de selección particular (Soomro et al., 2008; Pérez et al., 2018). La función más importante de este parámetro en los estudios genéticos de caracteres cuantitativos es su papel predictivo para indicar la confiabilidad del valor fenotípico con respecto al valor genotípico, como valor guía en el mejoramiento, además proporciona información sobre el grado en que ciertos rasgos pueden transmitirse a generaciones sucesivas (Mohamad et al., 2017). Caracteres con una alta heredabilidad permiten una mayor posibilidad de éxito en la selección (Ali et al., 2016), debido a la menor influencia de los efectos ambientales.
Los estudios de parámetros genéticos en especies forestales son amplios y trascienden en diversos países, donde la silvicultura tiene importancia económica, ecológica, cultural y/o etnobotánica (Escobar-Sandoval et al., 2018). Lo que ha permitido conocer en gran medida la estructura genética de las poblaciones, identificar y seleccionar árboles con características superiores para generar cambios en las frecuencias de genes y genotipos de una población.
En Colombia, se han estimado parámetros genéticos para caracteres de germinación y de dimensiones de semillas y, además, se han reportado trabajos sobre avances genéticos para caracteres como el DAP, volumen comercial, altura total y forma del fuste, en varias especies nativas (Rodríguez y Nieto, 1999; Espitia et al., 2010; Espitia et al., 2011; Correa et al., 2013; Espitia et al., 2016; Espitia-Camacho et al., 2018a).
El objetivo de esta investigación fue estimar la variabilidad fenotípica, los parámetros genéticos, la heredabilidad y la ganancia genética esperada para nueve características biométricas del fruto y las semillas de C. ebano.
MATERIALES Y MÉTODOS
Localización y periodo de evaluación
El estudio de campo se adelantó durante el año 2018 sobre frutos y semillas de árboles de C. ebano, colectados en el área de influencia de Montería-Colombia. El material vegetal fue estudiado en la Universidad de Córdoba, cuyas coordenadas corresponden a 8°47'16" N, y 75°51'28" W; localidad que se caracteriza por poseer una temperatura de 28 °C, humedad relativa del 84 %, precipitaciones anuales promedias de 1200 mm, altitud de 15 m. s. n. m, un brillo solar de 5,5 horas luz/día (2000 horas luz/año) y pertenece a la zona media del bosque seco tropical del valle del Sinú (Palencia et al., 2006).
Material genético forestal
La muestra estuvo constituida por diez árboles de ébano obtenidos mediante muestreo aleatorio simple. De cada árbol se colectaron al azar diez frutos sanos y maduros, para un total de 100 frutos. Al momento de la cosecha de los frutos, los árboles presentaron altura entre 5 y 10 m, edad de seis a 15 años y diámetro a la altura del pecho de 25 a 40 cm. Por su condición de planta alógama (Diallo et al., 2008), las semillas de cada uno de los frutos de cada árbol objeto de investigación, fueron consideradas como una familia de medios hermanos.
Variables fenotípicas
Un total de 11 variables cuantitativas concernientes a las características del fruto y las semillas de ébano fueron medidas: Peso fresco del fruto (PFR), largo máximo del fruto (LFR), ancho máximo del fruto (AFR), grosor máximo del fruto (GFR), número de semillas/fruto (NSF), peso total de las semillas por fruto (PSF), peso fresco de una semilla (P1S), peso de cien semillas (P100S), volumen de una semilla (V1S), densidad de una semilla (D1S) y el número de semillas por kilogramo (NSK).
En cada uno de los frutos de cada árbol fue medido el PFR (g), LFR (cm), AFR (cm), GFR (cm), NSF (#), PSF (g), en tanto que las variables asociadas con características de la semilla, como el P1S (g), V1S (mL) y D1S (g/mL) se estimaron como el promedio de las mediciones individuales por cada semilla presente dentro de cada fruto por árbol.
La D1S fue estimada a partir de la relación (P1S/V1S). El P100S se estimó a partir de la ecuación: P100S = P1S*100 y el NSK fue calculado como el cociente: (1x105 g / P100S).
Diseño experimental y análisis de datos
Las observaciones en las diez familias de medios hermanos, se evaluaron asumiendo un Diseño Completamente al azar, con diez repeticiones (frutos/árbol). Para cada una de las variables, se realizaron análisis de varianza a una vía, con efectos aleatorios, y se estimaron los coeficientes de variación fenotípicos (CVF), varianza fenotípica media entre familias de medios hermanos (VF), varianza genética media entre familias de medios hermanos (VG), varianza ambiental media entre familias de medios hermanos (VA), varianza genética aditiva media entre familias de medios hermanos (VGA), coeficientes de variación genotípicos (CVG), heredabilidad media familiar (h2Fam), heredabilidad en sentido estricto (h2 E), ganancia genética esperada (GGE) e índice de variabilidad genética b (CVG/CVF).
Para la ejecución del análisis de varianza y estimación de los anteriores parámetros genéticos, se utilizó el software computacional para Windows de libre acceso GENES V.2016.6.0 (Cruz, 2016).
La heredabilidad media familiar (h2 Fam) y en el sentido estricto (h2 E) para cada variable, fue estimada de la forma clásica, como se describe a continuación:
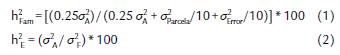
Donde: σ2 Fam = varianza genética media para familias de medios hermanos (HM); σ2 F= varianza fenotípica media para HM y σ2 A = varianza genética aditiva media para HM; estos parámetros fueron calculados a partir de los cuadrados medios esperados (Tabla 1).
Tabla 1 Fuentes de variación (FV), grados de libertad (gl) y cuadrados medios (CM) esperados para el análisis de varianza, bajo el uso del diseño completamente al azar con diez tratamientos y diez repeticiones.
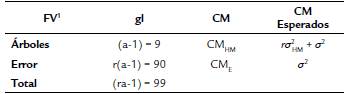
1 FV: fuentes de variación; gl: grados de libertad; a: número de árboles (10); r: repeticiones (10); CM: cuadrados medios; a: número de árboles (10); r: repeticiones (10); CMHM: Cuadrados medios para familias de medios hermanos (HM); CME: Cuadrados medios del error o residual; σ2 HM varianza media entre familias de HM; σ 2 HM = σ 2 G = varianza genética -(CMHM - CME) / r; σ 2: varianza ambiental de observaciones individuales = CME; σ 2 E: varianza ambiental media entre familias de HM, σ 2 E = (σ 2/r); σ 2 A: varianza genética aditiva media, σ 2 A = (4 σ 2 HM/r); σ 2 F: varianza fenotípica media entre familias de HM, σ 2 F = (σ 2 G + σ 2 E), (Venko-vsky y Barriga, 1992).
La ganancia genética esperada (GGE) para cada variable, se estimó a través de la siguiente ecuación (Venkovsky y Barriga, 1992; Espitia-Camacho et al., 2018a):

Dónde: k = diferencial de selección al 5 % de intensidad de selección, equivalente a 2,06; σ = Desviación estándar fenotípica; h2 E = heredabilidad en sentido estricto.
La GGE también se expresó como porcentaje de la media.
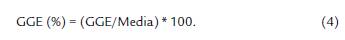
RESULTADOS
Análisis de varianza para las características biométricas
Los cuadrados medios del análisis de varianza consignados en la Tabla 2, acusaron diferencias altamente significativas (p < 0,01) entre árboles para siete de las 11 características: PFR, LFR, GFR, NSF, PSF, V1S y D1S, mas no para AFR, P1S, P100S y NSK.
Tabla 2 Análisis de varianza y coeficientes de variación para 11 características biométricas del fruto y las semillas de Caesalpinia ebano H. Karst. en Montería, Colombia. 2019.
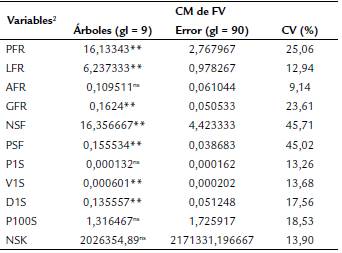
2 FV: fuentes de variación; CV (%): coeficiente de variación; gl: grados de libertad; **: significativos al 1% de probabilidad; ns: no significativo; PFR: peso fresco del fruto (g); LFR: largo del fruto (cm); AFR: ancho del fruto (cm); GFR: grosor del fruto (cm); NSF: número de semillas por fruto; PSF: peso de semillas por fruto (g); P1S: peso de una semilla (g); V1S: volumen de una semilla (mL); D1S: densidad de una semilla (g/mL); P100S: peso de cien semillas (g); NSK: número de semillas por kilogramo.
Las variables AFR, LFR, P1S, V1S, NSK, presentaron valores bajos de dispersión, con coeficientes de variación (CV) entre 9 y 14 %; D1S, P100S, GFR y PFR, se agruparon en valores medios de dispersión, con CV entre 18 y 25 %, mientras que PSF y NSF presentaron alta dispersión con CV de 45,71 % y 45,02 %, respectivamente, los cuales se consideraron muy altos (Tabla 2).
Parámetros genéticos
Los parámetros genéticos derivados de las características biométricas del fruto y las semillas de C. ebano se encuentran registrados en la Tabla 3, excepto para las variables P100S y NSK, que presentaron varianzas de muy baja magnitud de acuerdo con el análisis de varianza.
Los coeficientes de variación fenotípicos (CVF), oscilaron entre 9,14 % para el ancho del fruto (AFR), y 45,42 % en el número de semillas por fruto (NSF). Las estimaciones promedio de las varianzas genéticas (VG), resultaron menores que la varianza ambiental (VA) en dos caracteres, AFR y P1S. Las magnitudes de estos parámetros influyeron directamente en la magnitud de la varianza genética aditiva media (VGA), coeficiente de variación genético (CVG) y los índices de variabilidad genética b (Tabla 3).
Tabla 3 Parámetros genéticos esperados para nueve características biométricas del fruto y las semillas de Caesalpinia ebano H. Karst.
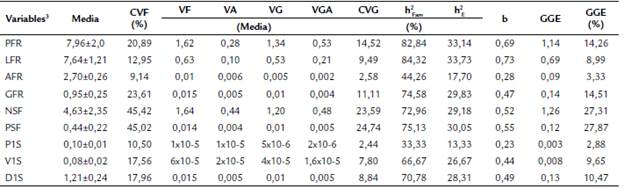
3 CVF: coeficiente de variación fenotípico; VF: varianza fenotípica media entre familias de medios hermanos (HM); VA: varianza ambiental media entre familias de HM; VG: varianza genética media HM; VGA (Media): varianza genética aditiva media entre HM; CVG: coeficiente de variación genotípico; h2 Fm: heredabilidad media familiar; h2 E: heredabilidad en sentido estricto; b: índice de variabilidad genética, relación (CVG/CVF); GGE: ganancia genética esperada; PFR: peso fresco del fruto (g); LFR: largo del fruto (cm); AFR: ancho del fruto (cm); GFR: grosor del fruto (cm); NSF: número de semillas por fruto; PSF: peso de semillas por fruto (g); P1S: peso de una semilla (g); V1S: volumen de una semilla (ml); D1S: densidad de una semilla (g/mL).
Los valores más altos de estimación de heredabilidad familiar (h2 Fam) y en el sentido estricto (h2 E) se encontraron para el largo del fruto (LFR) y peso fresco del fruto (PFR), con valores de 84,32 % y 33,73 %, y, 82,84 % y 33,14 %, respectivamente. Las nueve variables se agruparon acorde con su valor de h2 Fam en alta (> 50 %): LFR, PFR, PSF, GFR, NSF, D1S y V1S; y "media (20 - 50 %): AFR y P1S. Según los valores de h2 E, las variables se clasificaron en heredabilidad media (20-50 %): LFR, PFR, PSF, GFR, NSF, D1S y V1S baja (< 20 %): AFR y P1S (Tabla 3).
La ganancia genética esperada expresada como porcentaje de la media (GGE %) fue mayor al 10 % en cinco de las nueve variables evaluadas, y se agrupó como alta (GGE > 20 %): PSF y NSF; media (10 % > GGE < 20 %): GFR, PFR y D1S; y baja (gGE < 10 %): V1S, LFR, AFR y P1S (Tabla 3).
DISCUSIÓN
Análisis de varianza para las características biométricas
El valor de la significancia detectada en 64 % de las variables estudiadas, indica la existencia de diferencias genéticas entre árboles, la cual permitiría realizar selección y clonación de árboles con características biométricas de frutos y semillas deseables, ya que ello incide en cambio en la estructura genética de la población (Santos et al., 2018); estos hallazgos son coherentes con estudios preliminares en otras especies de la familia Fabaceae, como Poecilanthe parviflora Benth. (Valadares et al., 2009), Dalbergia cearensis Ducke. (Nogueira et al., 2010) y Mimosa scabrella Benth. (Menegatti et al., 2019).
Ausencia de significancia se encontró para las características asociada con la semilla, excepto en los caracteres volumen y densidad de una semilla, lo cual posiblemente sea reflejo de una base genética estrecha, generada por el efecto fundador (Espitia-Camacho et al., 2018a), lo que amerita la introducción de genes. La alta variabilidad para los valores fenotípicos restantes son respuesta a las variaciones de los factores genéticos y ambientales expresados de manera diferencial entre individuos (Mohamad et al., 2017), causado por su recombinación genética (Tabla 2), mediada por la polinización cruzada y los efectos climáticos (Luna-Cedeño et al., 2018), lo cual genera una población heterogénea y heterocigota, debido a la presencia de nuevas combinaciones genéticas, efectos ambientales y la interacción entre ellos como factores generadores de la variabilidad en estos caracteres (Gómez-Jiménez et al., 2010).
La mayorplasticidad fenotípica expresada por las variables de los frutos puede deberse a que su crecimiento está sujeto a la humedad del medio y a su posición en el árbol en el aprovechamiento de los fotoasimilados (Daymond y Hadley, 2008); pues los frutos de mayor tamaño se forman primero y se encuentran en la parte superior de la copa por la mayor disponibilidad de azúcares y proteínas (Houle et al., 2007). Estas variaciones dentro y entre los árboles están sujetas a varios factores, como la constitución genética a nivel de individuos, tipo de población y polinización, insectos plagas, agentes patogénicos, posición geográfica, nutrición y edad de la planta madre, deterioro y nivel de madurez a cosecha, los cuales afectan las características biométricas y fisiológicas del fruto y las semillas (Silva et al., 2015; Luna-Cedeño et al., 2018; Leão et al., 2018).
La existencia de valores fenotípicos no significativos en las variables de respuesta: AFR, P1S, P100S y NKS, sugiere la existencia de una importante semejanza genética entre los 10 árboles estudiados. Hecho que reduce la posibilidad de selección y lograr un avance genético, ya que la selección de árboles actúa sobre la diferencia genética existente entre individuos de una misma población (Machado et al., 2016; Shafique et al., 2016).
Parámetros genéticos
En la descomposición de la varianza fenotípica (VF), se detectó mayor participación de la varianza ambiental (VA) que la varianza genética (VG), en las características ancho de fruto (AFR), grosor del fruto (GFR), peso de una semilla (P1S), volumen de una semilla (V1S) y densidad de una semilla (D1S), más no así para las variables peso de fruto (PFR), longitud del fruto (LFR), número de semillas por fruto (NSF) y peso de semilla por fruto (PSF). Lo que puede obedecer al tamaño de la muestra usada en el estudio. Esto pudo repercutir en el índice de variabilidad genética (b) y la heredabilidad, dado que esta predice el potencial de transmisión de genes con acción aditiva a la siguiente generación, haciendo menor el progreso genético, ya que la parte ambiental no es heredable (Maia et al., 2016; Lopes et al., 2017).
Los coeficientes de variación fenotípica y variación genética otorgan información sobre la naturaleza y magnitud de la variación, resaltando la participación de los efectos ambientales en la variación fenotípica. Altos valores se presentaron para el número de semillas por fruto (NSF) y peso de semillas por fruto (PSF), lo cual denota la presencia de variabilidad genética en ellos y posibilidades de éxito a través de la selección de los árboles con mayores ventajas para la silvicultura (Tabla 3). Esta situación concuerda con los resultados de Moraes et al. (2015) en Eucalyptus grandis W. Hill., quienes sostienen que ello permite una ganancia significativa a través de la selección, lo que se corrobora en la Tabla 3.
La probabilidad de lograr avances importantes por selección fenotípica para los caracteres PFR y LFR, es concordante con los expresado por Correa et al. (2013) y Mohamad et al. (2017), quienes sostienen que la superioridad de la variación genética sucede cuando los efectos ambientales son menos influyentes en la medición de los valores fenotípicos. Maia et al. (2016), en trabajos de mejoramiento por selección, resaltaron que la heredabilidad en sentido estricto (h2 E) es de mayor importancia, debido a que determina cuánto de la apariencia fenotípica de las plantas es el reflejo exacto del valor genotípico aditivo y de la acción de los genes que responden a la selección.
Los altos valores para los parámetros genéticos VG, VGA, CVG e índice de variabilidad genética b, determinaron la alta heredabilidad media familiar (h2 Fam) y estricto (h2 E) detectada en las variables peso de fruto (PFR), longitud del fruto (LFR) y número de semillas por fruto (NSF) (Tabla 3). Los índices de variabilidad en la medida que se aproximen a valores iguales o mayores que uno, expresan una condición favorable para la selección (Moraes et al., 2015) y brindan mayor confiabilidad al describir el comportamiento de la heredabilidad en función de los componentes de la varianza (VF). Resultados similares han sido reportados en diferentes variables de interés en varias especies forestales (Batista et al., 2012; Moraes et al., 2015; Dias et al., 2015).
Los efectos genéticos aditivos que representan la varianza genética aditiva (VGA), no desaparecen fácilmente con la recombinación genética o caracteres superiores, ya que son heredables y permiten incrementar la frecuencia de alelos favorables en los caracteres de interés en la siguiente generación, conforman nuevas combinaciones genéticamente superiores y, por consiguiente, sustentan el progreso genético por selección en posteriores generaciones (Pérez et al., 2018; Farias-Neto et al., 2018).
Los caracteres NSF y PSF expresaron los mayores valores de CVG (23,59 % y 24,74 %; respectivamente) y ganancia genética esperada (GGE) de 27,31 % y 27,87 %, en su orden (Tabla 3). Estos valores son similares (CV = 24 % y GGE = 26 %) a los reportados en diferentes variables de interés en Pachira aquatica Aubl. (Espitia-Camacho et al., 2018a), Platonia insignis Mart. (Maia et al., 2016) y Euterpe oleracea Engel. Las magnitudes de estos parámetros indican que, en futuras evaluaciones en campo, podría haber mayor expresión de variación genética para otros caracteres asociados, con buenas perspectivas de variabilidad, para ser aprovechada a lo largo de un programa de mejoramiento genético (Correa et al., 2013).
La moderada y alta GGE detectada en este estudio sustenta la variación fenotípica significativa encontrada entre árboles para la mayoría de las variables (Tabla 3), todo ello en función de las divergencias genéticas generada en poblaciones aisladas de C. ebano, producto de la alogamia entre árboles, similar a la detectada por Diallo et al. (2008) en la subfamilia Caesalpinioideae.
La GGE obtenida para PFR, GFR, NSF, PSF y D1S, puede incrementarse en un posterior paso cuando se considere clonar los árboles seleccionados como superiores, en comparación a cuando se use solo semilla sexual de libre polinización. De acuerdo con lo documentado por varios autores (Murillo et al., 2012; Espitia et al., 2016), esto se explica porque en la clonación se captura toda la acción génica aditiva y no aditiva de los árboles superiores seleccionados, mientras que cuando se utiliza semilla sexual originada de polinización libre, sólo se aprovecha variación genética aditiva.
En general, los valores de la GGE acusaron un potencial de progreso genético relativamente alto en el mejoramiento de C. ebano en Córdoba. De acuerdo con Batista et al. (2012) y Ali et al. (2016), este avance integrado con el proceso de clonación de los mejores árboles puede constituir el complemento ideal de un programa de mejoramiento genético para cualquier especie forestal y/o frutal.
CONCLUSIONES
Se detectó variabilidad genética entre árboles para los caracteres: peso, largo, grosor y número de semillas por fruto, volumen y densidad de una semilla; la cual puede ser aprovechable por la magnitud de la varianza genética aditiva. Los valores más altos de heredabilidad media familiar (h2 Fam) y en sentido estricto (h2 E) se presentaron para las variables peso y largo del fruto, con magnitudes de h2 Fam > 82 % y h2 E > 30 %, por lo que todavía la selección y la clonación de árboles superiores basada en el fenotipo es suficiente para lograr progreso genético. La ganancia genética esperada expresada como porcentaje de la media, fue mayor al 10 % en cinco de las nueve variables evaluadas, lo que representa un potencial de progreso genético relativamente importante en el mejoramiento de los caracteres biométricos del fruto y la semilla de C. ebano. La estimación de los parámetros genéticos de los caracteres peso de fruto, largo de fruto y peso de semillas por fruto, indican que son los más importantes para el mejoramiento genético de C. ebano, de modo que la selección de frutos largos y pesados con semillas grandes y pesadas permitiría la obtención de plántulas vigorosas, tanto para la restauración de ambientes, como material de propagación para otros usos.

