











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Los Triatominae son insectos hematófagos pertenecientes a la familia Reduviidae (Heteroptera). Esta subfamilia cuenta con 151 especies divididas en cinco tribus y siete géneros (Justi y Galvão, 2017). La mayoría de estas especies han demostrado poder infectarse de forma natural o experimental con Trypanosoma cruzi Chagas, 1909, agente etiológico causante de la enfermedad de Chagas (Schofield y Galvão, 2009). Triatoma infestans Klug, 1834 se distribuye en Argentina, Bolivia, Brasil, Chile, Ecuador, Paraguay, Perú y Uruguay es una especie con importancia epidemiológica primaria que coloniza tanto domicilios como anexos peridomiciliarios (Gürtler et al., 2014; Belliard et al., 2019). Por otro lado, Triatoma platensis Neiva, 1913, de importancia epidemiológica secundaria (Schofield y Galvão, 2009), se distribuye en Argentina, Bolivia, Brasil, Paraguay y Uruguay (Salvatella et al., 1991; Galvão et al., 2003; Coscarón, 2017). En ambientes silvestres se encuentra asociada a nidos de Furnariidae y Psitaciidae, mientras que en el ambiente humano se halla en corrales de aves domésticas o bien, vuela hacia la vivienda atraída por la luz (Ferrero et al., 1999; Martí et al., 2014; Cavallo et al., 2016).
Estas especies, que pertenecen al subcomplejo Infestans, son interfértiles produciendo descendencia híbrida fértil en condiciones experimentales (Pérez et al., 2005) y en ambientes silvestres, donde Martí et al. (2014) registraron colonias mixtas con la presencia de individuos híbridos. Esta capacidad de entrecruzamiento interespecífico permitiría la producción de nuevos genotipos híbridos resultando en adaptaciones evolutivas que podrían permitir la colonización de nuevos hábitats (Pérez et al., 2005). Por otro lado, las intervenciones humanas en los ambientes naturales pueden aumentar el riesgo epidemiológico, facilitar la emergencia de enfermedades endémicas y crear nuevos entornos adecuados para la integración y cruzamiento entre especies (Correia et al., 2013). Es por ello que estudiar aspectos relacionados con la biología reproductiva de híbridos contribuye al conocimiento de la dinámica poblacional, dispersión y potencial adaptación a diferentes hábitats (Gurevitz et al., 2006).
Un aspecto relevante, en la biología reproductiva de las hembras híbridas es la oogénesis, proceso que se desarrolla en los folículos ováricos, es un período de rápido crecimiento y diferenciación celular que ocurre en tres estadios: previtelogénesis que implica la diferenciación del ovocito, vitelogénesis donde se acumula vitelo y por último coriogénesis donde se depositan las capas del corion (Fruttero et al., 2011; Lynch y Roth, 2011).
En este estudio se caracterizaron aspectos histológicos de la biología reproductiva de híbridos experimentales de T. infestans y T. platensis enfocados en el desarrollo folicular.
Los híbridos se obtuvieron de una colonia de 20 hembras adultas de T. infestans silvestres y 20 machos adultos de T. platensis de primera generación de laboratorio. Los triatominos fueron mantenidos bajo condiciones controladas de temperatura (27 ± 1 °C), humedad (60 ± 10 %) y fotoperíodo (12: 12 luz-oscuridad). Cada 15 días se les ofreció alimento sobre paloma (Columba livia, Gmelin, 1789) con movimiento restringido y sin anestesiar, hasta la obtención de 35 hembras adultas. Estas fueron alimentadas a repleción al séptimo día posterior a la muda, y se emparejaron con machos híbrido bien alimentados. Una vez comprobada al menos una cópula, a través de la presencia del espermatóforo, el macho fue retirado y a la hembra se le ofreció nuevamente comida. Aquellas alimentadas a repleción fueron asignadas a distintos grupos de sacrificio, según el número de días transcurridos después de la última ingesta (0, 3, 5, 7, 10, 14 y 16 días post ingesta), a fin de obtener folículos basales en sus distintos estadios. Transcurrido el tiempo estimado se extrajeron los ovarios de las hembras sacrificadas y se fijaron en buffer fosfato salino (PBS) a pH = 7. Luego se deshidrataron en soluciones de etanol de graduación creciente, aclararon en xilol e incluyeron en parafina. Posteriormente se realizaron cortes a 7 um y colorearon con hematoxilina-eosina (Tolosa et al., 2003).
En relación a la histología del proceso de desarrollo folicular, este inicia con ovocitos en previtelogénesis temprana, de estructura fibrosa y homogénea, con volumen reducido, alimentados por cordones tróficos de similares características, típicos de ovarios telotróficos meroísticos, los que conectan las haplocélulas con el estroma del germario (Fig. 1a, b). Durante la previtelogénesis tardía, los ovocitos manifiestan un importante crecimiento en volumen, el citoplasma aumenta considerablemente de tamaño, por la acumulación de nutrientes provenientes del germario conservando la estructura fibrosa (Fig. 1c). La fase de vitelogénesis temprana se caracteriza por la degeneración de los cordones tróficos y el ingreso de vitelogeninas desde la periferia formando gotas de vitelo con la consecuente reclusión del ooplasma hacia el centro de la célula (Fig. 1d). Una vez finalizado el proceso de acumulación de vitelo, el interior de la gameta se encuentra repleto de grânulos y sustancias de reserva, lo que aumenta rápidamente su volumen en vitelogénesis tardía (Fig. 1e).
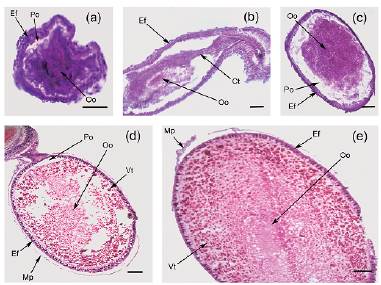
Figura 1: Secuencia de desarrollo folicular en ovarios de hembras híbridos entre T. infestans y T. platensis (a-e). a) Ovocito en previtelogénesis temprana (corte transversal) (200X HE, barra de escala = 300 μm). b: Ovocito en previtelogénesis temprana (corte longitudinal) (100X HE, barra de escala = 300 μm). c) Ovocito en previtelogénesis tardía (100X HE, barra de escala = 300 μm). d) Ovocito en vitelogénesis temprana (100X HE, barra de escala = 300 μm). e) Ovocito en vitelogénesis tardía (100X HE, barra de escala = 300 μm). Ct: cordón trófico; Ef: epitelio folicular; Mp: membrana peritoneal; Oo: ooplasma; Po: espacio perioocítico.
En relación al desarrollo del epitelio folicular se advierten cambios en los diferentes estadios. En previtelogénesis temprana, el folículo basal está rodeado por un epitelio simple con células pequeñas y fusiformes, con un núcleo central y cromatina condensada, lo que denota poca actividad (Fig. 2a). En previtelogénesis tardía las células del tejido epitelial adquieren formas cilíndricas, con núcleos centrales redondeados y cromatina laxa haciendose notoria la presencia del nucléolo. Este cambio a nivel nuclear indica un aumento de la actividad metabólica (Fig. 2b). En la vitelogénesis temprana, a nivel del epitelio, ocurre divisiones mitóticas sin cariocinesis, de modo que todas las células se observan binucleadas, voluminosas y cilíndricas, presentando el extremo basal en contacto con el espacio perioocítico y el apical con la membrana peritoneal que separa al ovocito de la hemolinfa, además el volumen de los núcleos aumenta notándose la cromatina aún más laxa que en la fase anterior. Cabe destacar la presencia de un nucléolo en cada núcleo (Fig. 2c). En la fase de vitelogénesis tardía, se produce un estiramiento del epitelio folicular como consecuencia del aumento rápido de volumen que sufre la gameta por el ingreso de sustancias de reserva y agua generando un achatamiento del tejido, en esta situación las células se observan con forma cúbica, de menor tamaño y manteniendo el carácter binucleado con núcleos pequeños (Fig. 2d).
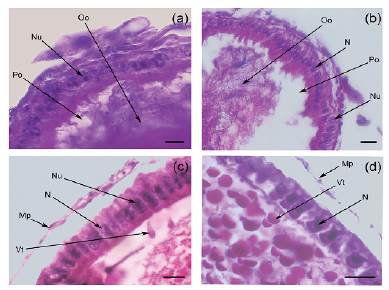
Figura 2: Secuencia de desarrollo de las células del epitelio del folículo basal en ovarios de hembras híbridos entre T. infestans y T. platensis (a-d). a) Epitelio folicular de ovocito en previtelogénesis temprana (800X HE, barra de escala = 100 μm). b) Epitelio folicular de ovocito en previtelogénesis tardía (400X HE, barra de escala = 100 μm). c) Epitelio folicular binucleado de ovocito en vitelogénesis temprana (400X HE, barra de escala = 100 μm). d) Epitelio folicular de ovocito en vitelogénesis tardía (400X HE, barra de escala = 100 μm). Mp: membrana peritoneal; N: núcleo; Nu: nucléolo; Oo: ooplasma; Po: espacio perioocitico; Vt: vitelo.
Los resultados evidencian que las fases del desarrollo folicular en hembras híbrido se encuentran bien definidas produciendo huevos viables con un microambiente apto para dar lugar a la embriogénesis y que los cambios ocurridos a nivel histológico en los folículos basales se asemejan a los registrados en T. infestans (Barth, 1973). Estos resultados corroboran lo propuesto por Pérez et al. (2005), sobre la existencia de híbridos fértiles, del subcomplejo Infestans, con capacidad de generar descendencia viable, lo cual sumado a la invasión de ambientes humanos por especies silvestres podría sugerir la existencia de colonias naturales híbridas asociadas a ecotopos domésticos, que funcionen como refugio para estos híbridos y que ocasionalmente puedan dispersarse hacia la vivienda humana convirtiéndose en un riesgo para la salud de sus ocupantes, en este sentido, López et al. (2020) describen y comparan el patrón de ingesta y defecación de híbridos con T. infestans, señalando que adultos y ninfas híbridos poseen comportamientos de alimentación y defecación similares a T. infestans concluyendo que estos organismos podrían tener un potencial relativamente alto de transmisión de T. cruzi convirtiéndose en un riesgo epidemiológico de importancia. Por ello, indagar con mayor profundidad en el estudio de la eficiencia reproductiva del híbrido cobra importancia desde el punto de vista de la capacidad de colonización de la vivienda, de la capacidad vectorial y del control de triatominos.

