











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Las plantas pertenecientes a la división Marchantiophyta, también conocidas como hepáticas, hacen parte del grupo monofilético Bryophyta (Zhang et al., 2020), plantas terrestres no vasculares ubicadas en el clado basal de las Embriophytas. Presentan una amplia distribución y se han registrado aproximadamente 7500 especies a nivel mundial en zonas con alta humedad y baja intensidad lumínica (Sõderstrõm et al., 2016). Colombia cuenta con 715 especies, siendo la región Andina la que presenta mayor riqueza, allí se registran130 géneros distribuidos en siete órdenes (Uribe y Gradstein, 1999; Gradstein, 2016a; Gradstein et al., 2018; Bernal et al., 2019).
Las hepáticas cumplen un papel importante en los ciclos hídricos de los ecosistemas, retienen grandes cantidades de agua y la liberan lentamente, a su vez presentan tolerancia a la desecación mediante la suspensión de su metabolismo en tiempos de sequía, y en tiempos húmedos retoman su actividad fotosintética y de crecimiento (Goffinet y Shaw, 2009). Dada la fisiología de las hepáticas basada en la ectohídria, estas plantas transportan el agua y los nutrientes dentro y fuera de la planta a través de espacios capilares, permitiendo así la regulación hídrica (Goffinet y Shaw, 2009; Vanderpoorten y Goffinet, 2009). Además, tienen la capacidad de almacenar y fijar grandes cantidades de CO2 (Goffinet y Shaw, 2009).
Uno de los servicios ecosistémicos que brindan las hepáticas es la formación de suelo, por su interacción con otros organismos como líquenes y hongos, esto mediante la aceleración de los procesos de meteorización, tanto física como química del material orgánico e inorgánico arrastrado por el viento y la escorrentía (Vanderpoorten y Goffinet, 2009).
Estas plantas se pueden ver afectadas de manera directa por cambios abruptos en el ecosistema, ya sea por actividades como la tala de árboles y/o la ganadería (Laurance, 1998), esto debido a que, la mayoría de las hepáticas son de hábito epifito, es decir, que se desarrollan sobre una planta hospedera o forófito (Barkman, 1958; Wolf, 1993). Otras causas de la afectación sobre las comunidades de briófitos están asociadas al cambio climático, dada la baja capacidad de resiliencia de estas especies (Holz y Gradstein, 2005), por otro lado, la fragmentación de los bosques genera pérdida de la diversidad y efectos negativos sobre los procesos de regulación hídrica en los ecosistemas (Vanderpoorten y Goffinet, 2009).
Varios estudios se han realizado en hepáticas en zona de páramo y bosque altoandino en Colombia (Wolf, 1995; Wolf, 2003; Orrego, 2005; Gil y Morales, 2014; Vargas y Morales, 2014; Feuillet y Torres, 2016; Cacua et al., 2018), estos estudios se han facilitado debido a las condiciones ambientales óptimas para el desarrollo y crecimiento de estas plantas; sin embargo, en el Parque Nacional Natural Chingaza solo se registran dos estudios, un estudio florístico (Madriñán, 2012), el cual arrojó poca representatividad de la diversidad que este lugar en realidad alberga solo se registran las familias Jubulaceae, Herbertaceae y Marchantiaceae, y un trabajo realizado por Santana y Aguirre (1986) donde se presentan unas claves preliminares a género para las hepáticas de este sector.
El sector Suasie en el PNN Chingaza cuenta con una amplia variedad de ecosistemas que van desde zonas de bosque andino hasta páramo, este sector es de gran importancia ecológica, al punto de ser considerado un "hotspot" dentro del Parque, dada la alta diversidad de plantas y la presencia asidua de especies sombrilla como el oso de anteojos (Parques Nacionales Naturales de Colombia [PNN], 2020). Esta investigación pretende contribuir al conocimiento del estado actual de las hepáticas en los ecosistemas andinos y de páramo de Colombia. El objetivo general de este proyecto fue determinar la diversidad de hepáticas epífitas del sector Suasie del Parque Nacional Natural Chingaza, reportando datos sobre la composición, riqueza y abundancia de la comunidad, además se evaluó la variación de la diversidad entre gradientes altitudinales, asociados a diferentes coberturas vegetales.
MATERIALES Y MÉTODOS
Área de estudio
El Parque Nacional Natural Chingaza se encuentra ubicado en la cordillera Oriental, en el rango altitudinal que va desde 800 hasta 4020 m (NO, m s n m) rango que permite la amplia distribución de hepáticas, puesto que presenta ecosistemas variados como humedales, páramos y bosques húmedos. Es un área protegida que cuenta con un complejo de 60 lagunas y un embalse, denominado "Chuza", el cual aporta el 80 % de agua potable a la ciudad de Bogotá, además de surtir a los 11 municipios que comprende el Parque y así brindar un gran servicio ambiental (PNN, 2020).
Los muestreos se hicieron en Suasie, ubicado en el municipio de Fómeque - Cundinamarca (Fig. 1). Chingaza presenta un régimen de lluvias monomodal en los meses de mayo a agosto (Morales et al., 2007), con una precipitación media anual de 4022 mm, presenta una alta nubosidad y un promedio de 3,5 horas de radiación directa con 85 % de humedad relativa anual, la temperatura promedio varia durante el año presentando 25 °C en los meses de diciembre a febrero y de 13 °C en los meses de junio y agosto (González et al., 2017). La flora se encuentra principalmente representada por angiospermas de la familia Asteraceae seguido de las familias Melastomataceae, Bromeliaceae y Ericaceae (Vargas y Pedraza, 2004).
Toma de muestras
Para la recolecta de hepáticas epifitas se realizó una salida de campo del 11 al 13 de junio del 2019 y se siguió la metodología propuesta por Wolf (1993), Orrego y Uribe (2004) y Campos et al. (2019). Se realizaron tres transectos de 2x50m, los cuales fueron ubicados en diferentes rangos altitudinales, en distintas unidades vegetales. En cada uno de los sitios de muestreo se escogieron al azar cuatro fustes de diferentes forófitos, para un total de 12 (Fig. 2), las muestras fueron recolectadas de manera oportunística y se tomaron desde la base hasta 2 m de altura del fuste, en especies arbóreas, y hasta 1 m de altura para especies de tipo arbustivo y achaparrado.
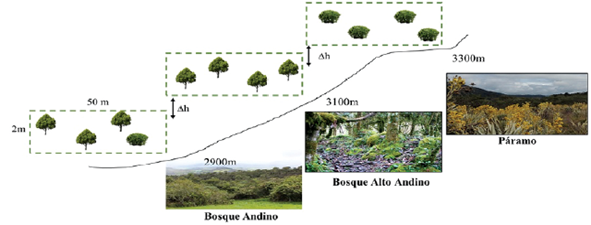
Figura 2 Esquema de las parcelas de muestreo de 2x50 m para cada localidad, con la variación altitudinal y el tipo de cobertura.
Las unidades vegetales tenidas en cuenta durante el estudio correspondieron a bosque andino, altoandino y páramo (Avellaneda et al., 2014; Sarmiento y León, 2015). A continuación se presenta una breve descripción de cada una de las unidades: 1) Bosque andino (BA) ubicado a una altura de 2900 m; 3100 m y 3300 m (NO m s n m) a 4°37'33,84" N y 73°43'36,72" W, caracterizado por vegetación arbórea y arbustiva, abundantes claros de bosque asociados a un cuerpo de agua y zonas de interacción antropogénica; 2) Bosque alto andino (AA) ubicado a una altura de 3100 m. s. n. m. a 4°37'24,18" N y 73°43'31,68" W, caracterizado por vegetación arbórea y arbustiva, en esta zona es evidente una estratificación que va desde un estrato rasante hasta dosel, y 3) páramo (P) a 3300 m. s. n. m. a 4°37'7,92" N y 73°43'36,24" W, caracterizado por vegetación achaparrada, arbustos y matorrales de especies nativas típicas de páramo (Fig. 2).
Los ejemplares se procesaron en los laboratorios de la Universidad de La Salle siguiendo las técnicas establecidas para las colecciones de briófitos (Gradstein et al., 2001). La identificación a especie de los ejemplares recolectados se realizó siguiendo las claves para géneros de hepáticas de Gradstein et al. (2001) y Gradstein (2016a) y otras específicas para cada uno de los géneros. Las muestras fueron depositadas en la colección del Museo La Salle. La nomenclatura empleada fue basada en el Catálogo de Plantas de Colombia (Gradstein y Uribe, 2016) y corroborada con el trabajo de Sõderstrõm et al. (2016).
Análisis de datos
Se tuvo en cuenta el número de especies de hepáticas encontradas en el estudio para el análisis de riqueza, una curva de acumulación de especies con el software EstimateS (Colwell, 2013) y los estimadores no paramétricos de ACE y Chao 1 (Magurran y McGill, 2011) fueron tenidos en cuenta en el análisis, las gráficas fueron realizadas en Graphpad Prism 5 (GraphPad, 2020). En el análisis de abundancia se realizó a partir del tratamiento de datos con logaritmos en Excel, las curvas rango - abundancia fueron elaboradas con Graphpad Prism 5.
Para el análisis de diversidad alfa, los datos se analizaron con el software Past (Hammer, 2020) donde se calcularon los índices de: Diversidad de Simpson (1-D), el cual establece que valores cercanos a la unidad presentan una diversidad mayor y valores cercanos a 0 presentan una gran dominancia y menor diversidad; Dominancia (D), el cual establece que valores cercano a la unidad presentan mayor dominancia y menor diversidad y valores cercanos a 0 una mayor diversidad; Shannon-Weiner (H'), el cual hace referencia a la uniformidad de las especies donde valores normales se encuentran en el rango de 2 y 3, en valores inferiores a 2 se habla de una baja diversidad y en valores superiores a 3 una alta diversidad de especies y finalmente, Equitatividad (J'), el cual mide la proporción de la diversidad observada con relación a la máxima diversidad esperada cuyo valor cercano a la unidad corresponde a situaciones donde todas las especies son igualmente abundantes (Magurran y McGill, 2011).
La diversidad beta se estimó con un análisis de similitud, teniendo en cuenta el índice de Bray-Curtis, este índice calcula la disimilitud entre dos zonas distintas, valores cercanos a 0 significan que dos lugares tienen la misma composición y valores cercanos a 1 los sitios no presentan especies compartidas (Magurran y McGill, 2011). Además, se realizó un CA (análisis de correspondencia por sus siglas en ingles), estos procedimientos se llevaron a cabo con el uso del software Past y RStudio (RStudio, 2020) respectivamente. En el caso de RStudio se hizo uso de las librerías "FactorMineR", "factoextra" y "ca".
RESULTADOS
Se evaluaron 12 forófitos en tres sitios de muestreo con diferente rango altitudinal y cobertura vegetal, fueron registrados 100 individuos pertenecientes a 47 especies, 21 géneros y 15 familias (Tabla 1). La flora de hepáticas fue dominada por la familia Plagiochilaceae (22,2 %), seguida por Lepidoziaceae (20,0 %), sólo 10,6 % de las especies registradas, correspondieron al grupo de las talosas, los géneros registrados fueron Riccardia, Metzgeria y Pallavicinia.
Tabla 1 Composición florística de hepáticas en el sector Suasie del PNN Chingaza, distribución taxonómica según especie y familia
| Especie | Andina | Alto Andina | Páramo |
|---|---|---|---|
| Acrobolbaceae | |||
| Acrobolbus wilsonii Nees. | x | ||
| Aneuraceae | |||
| Ricardia capillacea (Steph.) Meenks & C.De Jong. | x | ||
| Cephaloziellaceae | |||
| Kymatocalyx dominicensis (Spruce) Váña. | x | x | |
| Frullaniaceae | |||
| Frullania brasiliensis Raddi. | x | x | |
| Frullania densifolia Steph. | X | ||
| Frullania ecklonii (Spreng.) Spreng. | X | ||
| Frullania lobatohastata Steph. | X | ||
| Frullania pittieri Steph. | X | ||
| Frullania riojaneirensis (Raddi) Spruce. | x | ||
| Herbertaceae | |||
| Herbertus acanthelius Spruce. | X | ||
| Herbertus bivittatus Spruce. | X | ||
| Herbertus grossispinus Fulford. | X | X | X |
| Herbertus juniperoideus (Sw.) Grolle. | X | ||
| Lejeuneaceae | |||
| Blepharolejeunea securifolia (Steph.) R.M.Schust. | x | ||
| Cheilolejeunea fragrantissima (Spruce) R.M.Schust. | x | ||
| Cheilolejeunea revoluta (Herzog) Gradst. & Grolle. | x | ||
| Lejeunea debilis (Lehm. & Lindenb.) Nees & Mont. | X | ||
| Lejeunea elongella Gottsche. | X | ||
| Lejeunea. inflexilobaJ.B.Jack & Steph. | X | ||
| Lepidolejeunea involuta (Gottsche) Grolle. | X | ||
| Lepidolejeunea sullivantii (Gottsche) M.E.Reiner. | X | ||
| Marchesinia brachiata (Sw.) Schiffn. Lepicoleaceae | x | ||
| Lepicolea pruinosa (Taylor) Spruce. | x | ||
| Lepicolea ramentifissa Herzog | x | x | |
| Lepidoziaceae | |||
| Bazzania hookeri (Lindenb.) Trevis. | x | ||
| Bazzania jamaicensis (Lehm. & Lin-denb.) Trevis. | X | x | |
| Lepidozia brasiliensis Steph. | x | ||
| Pseudocephalozia quadriloba (Steph.) R.M.Schust. | x | x | |
| Lophocoleaceae | |||
| Leptoscyphus intermedius Grolle. | x | ||
| Metzgeriaceae | |||
| Metzgeria cleefii Kuwah. | x | ||
| Metzgeria consanguinea Schiffn. | x | ||
| Metzgeria leptoneura Spruce. | X | ||
| Pallaviciniaceae | |||
| Pallavicinia lyellii (Hook.) Gray. | x | ||
| Plagiochilaceae | |||
| Plagiochila adianthoides (Sw.) Lindenb. | x | ||
| Plagiochila bryhnii Steph. | x | x | x |
| Plagiochila exigua (Taylor) Taylor. | x | ||
| Plagiochila fuscolutea Taylor. | x | ||
| Plagiochila heteromalla (Lehm. & Lindenb.) Lindenb. | x | ||
| Plagiochila heterophylla Lehm. | x | ||
| Plagiochila ovata Lindenb. | X | ||
| Plagiochila pachyloma Taylor. | x | ||
| Plagiochila papillifolia Steph. | X | ||
| Plagiochila simplex (Sw.) Lindenb. | X | x | x |
| Plagiochila turgida Herzog. | X | ||
| Radulaceae | |||
| Radula voluta Taylor. | x | ||
| Scapaniaceae | |||
| Scapania portoricensis Hampe & Gottsche. | x | ||
| Trichocoleaceae | |||
| Trichocolea sp. | x | x |
El género más representativo en el área de estudio fue Plagiochila con un 22,2 % de las especies, seguido por Pseudocephalozia (13,3 %), Frullania (10,0 %), Herbertus (7,8 %) y Bazzania y Scapania (5,6 % ) respectivamente. Las familias Plagiochilaceae y Lepidoziaceae obtuvieron la mayor representatividad en el estudio con 42,2 % de las especies recolectadas.
Al estudiar la composición en las tres altitudes se encontró que las hepáticas presentes en el bosque andino (BA) se encuentran distribuidas en cuatro órdenes, 13 familias, 15 géneros y 32 especies, las presentes en el bosque altoandino (AA) se encuentran distribuidas en tres órdenes, ocho familias, nueve géneros y 15 especies, y finalmente las presentes en páramo (P) se encuentran distribuidas en tres órdenes, nueve familias, diez géneros y 13 especies.
Basado en evidencia estadística, no se encontró diferencia significativa en la riqueza de hepáticas entre los diferentes pisos altitudinales (Kruskal-Wallis, p= 0,1271). En BA se encontraron 31 especies, siendo esta zona la de mayor riqueza en el estudio, seguida por AA, y finalmente la zona de P. En la curva de acumulación de especies se observó un comportamiento asintótico del índice de representatividad (Fig. 3), lo que indica relación entre la riqueza observada y esperada (S), frente a los estimadores no paramétrico ACE (91,43 %) y Chao 1 (72,92 %).
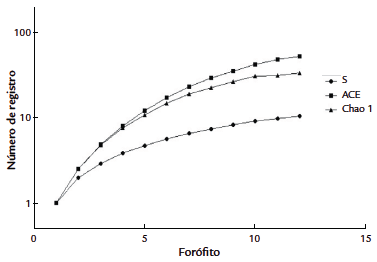
Figura 3 Curva de acumulación de especies de los datos recolectados en tres transectos por forófito según gradiente altitudinal.
La abundancia fue tomada a partir de la incidencia de los individuos en cada uno de los forófitos en los tres transectos establecidos, obteniendo un total de 100 registros. Se encontró diferencia significativa en la abundancia entre los gradientes (Kruskal-Wallis, p= 6,933e-05), BA presentó el mayor registro de abundancias con 32 especies en 44 registros, donde Plagiochila simplex (Sw) Lindenb fue la más representativa, en AA se registraron 15 especies en 31 registros con Pseudocephalozia quadriloba R.M Schust como la más abundante, tanto en esta zona como en las otras dos, y en P se registraron 13 especies en 15 registros con Pallavicinia lyellii (Hook.) Gray y P. simplex con la mayor representatividad.
Mediante la estimación de los índices de diversidad se obtuvo un valor alto para el índice de dominancia de Simpson (1-D) en las tres zonas para BA se presentó el mayor valor del índice (0,961), seguido de P (0,915) y finalmente AA (0,828); para el índice de dominancia (D) BA obtuvo el mayor valor de diversidad (0,038), seguido por P (0,084) y finalmente AA (0,171). Con el índice de Shannon-Weiner (H') se obtuvo para BA una alta diversidad de especies (3,375), mientras que para AA (2,234) y P (2,523) se presentó una uniformidad normal. Para la equitatividad (J') se encontró que para las tres zonas, tanto BA (0,973), AA (0,825) y P (0,983) presentan especies muy similares en cuanto a la abundancia.
Para la determinación de la diversidad beta se empleó el índice de Bray-Curtis entre BA y AA (0,128) y entre BA y P (0,267), los valores obtenidos permitieron establecer que dichas "parejas" presentan mayor similitud en su composición, mientras que entre P y AA (0,429) se da una menor similitud en su composición.
El dendrograma de similitud agrupó a AA y BA, ratificando lo obtenido por el índice de Bray-Curtis dando así la categoría de grupos con composición más similar, mientras P se comporta como un grupo aislado dado que presenta una composición disímil frente a BA y AA (Fig. 4a), esto posiblemente por la diferencia entre las unidades vegetales. Solo las especies de Herbertus grossispinus Fulford, Plagiochila bryhnii Steph y P. simplex se encontraron en las tres zonas muestreadas, siendo así especies compartidas en el estudio, las especies Kymatocalyx dominicensis (Spruce) Vána, Trichocolea sp y Frullania brasiliensis Raddi se encontraron compartidas en BA y P y Lepicolea ramentifissa Herzog, Bazzania Jamaicensis (Lehm. & Lindenb) y Pseudocephalozia quadriloba se encontraron compartidas en AA y P.
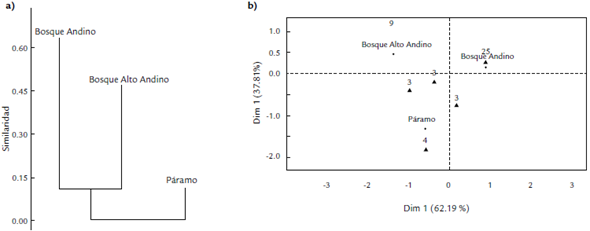
Figura 4 Análisis de Diversidad Beta a) Dendograma de similitud basado en el índice de Bray-Curtis para los diferentes gradientes altitudinales. b) Análisis de correspondencias para los diferentes gradientes altitudinales.
En el análisis de correspondencias (Fig. 4b) se obtuvo el 100 % de la representación de la información donde se definen tres grupos, de los cuales P presenta un agrupamiento separado de las especies y no tan cercano como lo presenta AA y BA; cerca al eje central aparecen las tres especies que se comparten en las tres zonas de estudio, y cerca a estas están las especies compartidas en dos de las tres zonas. AA y BA son el grupo de zonas más cercano y esto permite demostrar y corroborar la similitud entre ellas (Fig. 4a), mientras que P se encuentra a una mayor distancia frente a las otras zonas, evidenciando que esta es disímil.
DISCUSIÓN
La composición presente en el sendero Suasie del PNN Chingaza mostró a las familias Lejeuneaceae y Plagiochilaceae como los taxones de mayor representatividad, de la misma forma que lo exponen para estos ecosistemas Álvaro et al. (2007), Gradstein (2009) y Cacua et al. (2018). La familia Lejeuneaceae presenta una mayor riqueza en el territorio colombiano y más en la región Andina, en la cual representa el 90 % de las especies registradas (Uribe y Gradstein, 1999; Vargas y Morales, 2014; Cacua et al., 2018), debido al alto nivel de especialización fisiológico que presentan (Heinrichs et al., 2007; Wilson et al., 2007); en este estudio Plagiochilaceae tuvo la mayor riqueza, esto se debe al género Plagiochila que según Wolf (1993) y Álvaro et al. (2007) es el más rico en especies lo que permite un amplio rango de distribución, se obtuvieron 11 especies de este género, distribuidas en los tres rangos altitudinales, donde P. bryhnii y P. simplex fueron las únicas especies compartidas en las tres altitudes. Esta familia presenta una pigmentación que varía entre verde a marrón (Gradstein, 2016b) que le permite ventajas en procesos fisiológicos, desarrollándose en diferentes ambientes, desde húmedos hasta secos (Wolf, 1993). Es importante destacar la presencia de la especie P. brhynii como el primer registro para Cundinamarca a 2900 a 3300 m. s. n. m. (Bernal et al., 2019), se estima que esta especie es de amplia distribución en la región Andina.
Por otro lado, la familia Lejeuneaceae se encontró representada por cinco géneros Blepharolejeunea, Lejeunea, Lepidolejeunea, Marchesinia, con un total de siete especies en BA y Cheilolejeunea con dos especies en AA y P. La familia Herbertaceae con el género Herbertus también estuvo representada en las tres zonas estudiadas, teniendo a H. grossispinus como la única especie de este género en estar en los tres gradientes altitudinales, esto permite que estas dos familias también sean consideradas con un amplio rango de distribución y que por lo tanto sean asociadas a condiciones de especies generalistas o de especies con un amplio rango de tolerancia (Bernal et al., 2019).
El registro de Acrobolbus wilsonii Ness, es de gran relevancia, dado que esta especie sólo se conocía por una cita de Stephani (1917) sin localidad, de esta manera sería el primer registro con localidad precisa para Colombia a una altura de 2900 m. s. n. m. Acrobolbus es un género que en nuestro país tiene una distribución netamente andina (Bernal et al., 2019).
Este estudio abarcó las hepáticas epífitas del sector Suasie, sin embargo, llama la atención la presencia de la especie talosa Pallavicinia lyellii una especie de hábito principalmente terrestre, P. lyellii fue recolectada en la base del fuste, esto se debe al continuo en la brioflora que se presenta entre el suelo y la primera zona del fuste (a 1m de altura de la base del forófito) ya que las condiciones de humedad, luminosidad y temperatura son muy similares, en este continuo la composición de las especies se comparte encontrándose elementos típicos de zonas terrestres en la base del fuste o viceversa (Campos et al., 2015).
Estudios realizados por Wolf (2003), Feuillet y Torres (2016) y Cacua et al. (2018) muestran que a 2900 m se presentan mayores valores de riqueza y que a mayor altitud este valor disminuye como se observa en este estudio, que aunque la riqueza no presentó diferencias significativas entre las tres zonas estudiadas, BA obtuvo el mayor valor de riqueza mientras que en P fue el valor más bajo de las tres zonas, y esto podría deberse en como lo muestra Campos et al. (2015) y Cataño et al. (2021), a las condiciones microclimáticas presente en cada una de las zonas y el tipo de cobertura vegetal presente.
Según Campos et al. (2015) la riqueza se muestra en una alta proporción de familias con pocos géneros y pocas especies como se observa en este estudio. Pseudocephalozia quadriloba es la única especie con la mayor abundancia la cual estuvo presente en zonas de AA y P, dado que esta especie tiene un buen desarrollo en ambientes descubiertos, como los de Páramo y espacios húmedos, como los de Bosque Alto Andino (Álvaro et al., 2007).
Wolf (2003) evidencia en su estudio patrones de diversidad a lo largo de transectos altitudinales que para rangos entre 2550 y 3190 m. s. n. m. la diversidad de briófitos es mayor, como se demuestra en los resultados, en donde la diversidad alfa de las tres altitudes fue alta y se podría considerar que solo en el sector de Suasie del PNN Chingaza la presencia de hepáticas representa el estado de los ecosistemas del sector.
En la diversidad beta se tiene que P se diferenció de manera significativa frente aBAy AA posiblemente por condiciones de luminosidad, humedad y velocidad del viento (Campos et al., 2015), las cuales pueden afectar de manera estructural y fisiológica las especies presentes en las tres altitudes, sumado a esto están los diferentes tipos de coberturas vegetales que según lo observado permite justificar la relación existente entre Bosque Andino y Bosque Alto Andino, ya que en estos se comparten especies arbóreas totalmente diferentes a las que están presentes en el Páramo.
CONCLUSIONES
La composición en la comunidad varía en el gradiente altitudinal del sector de Suasie del PNN Chingaza, debido a las diferencias en las características microambientales determinadas por la estructura y composición vegetal de cada zona. BA presentó la mayor riqueza de especies, la heterogeneidad del hábitat que presenta este tipo de cobertura, además de la humedad, brindan las condiciones óptimas para el desarrollo de estas plantas de tipo poiquilohídrico. La vegetación influye directamente sobre la distribución de las hepáticas, encontrándose que, de 47 especies registradas en el estudio, solo tres especies son compartidas entre los gradientes con cobertura vegetal diferenciada. Por la distribución de las especies se evidencia la preferencia de estas por zonas umbrías, es decir, zonas con menos radiación solar y mayor humedad, lo que se relaciona perfectamente con la preferencia de los epifitos a estos tipos de hábitat. La riqueza y la diversidad alfa no presentaron variación en el gradiente altitudinal mientras que, para la diversidad beta, si existe variación teniendo mayor similitud BA con AA, diferenciándose así de P. La zona de páramo exige mayor especialización fisiológica de las especies al tener que sortear cambios bruscos en las condiciones climáticas razón por la que presentan el menor número de especies compartidas con las otras zonas o coberturas.

