










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Salud Uninorte
Print version ISSN 0120-5552On-line version ISSN 2011-7531
Salud, Barranquilla vol.24 no.2 Barranquilla July/Dec. 2008
ARTÍCULO DE REVISIÓN / REVIEW ARTICLE
REVISIONES BÁSICAS / BASIC REVIEW
Las proteínas alergénicas: un novedoso blanco para el desarrollo de estudios en proteomica funcional
The allergenic proteins: a novel target for the studies' development in functional proteomica.
Elkin Navarro1, Gloria Garavito2, Luis Alejandro Barrera3, Leonardo Lareo4, Eduardo Egea5
1 Biólogo. Joven Investigador Colciencias, 2007 (Convenio 090-2006) Grupo de investigación en Inmunología y Biología Molecular, Universidad del Norte. equiroz@uninorte.edu.co
2 Médico Inmunólogo, PhD. Profesor - investigador Grupo de investigación en Inmunología y Biología Molecular, Universidad del Norte. ggaravit@uninorte.edu.co
3 Biólogo, PhD. Director Instituto de Errores Innatos del Metabolismo. Pontificia Universidad Javeriana. abarrera@javeriana.edu.co
4 Biólogo, PhD. Director Grupo de investigación en Bioquímica computacional y estructural y bioinformática. Pontificia Universidad Javariana. llareo@javeriana.edu.co
5 Médico Inmunólogo, MSc. Director Grupo de investigación en Inmunología y Biología Molecular, Universidad del Norte. eegea@uninorte.edu.co
Correspondencia: Universidad del Norte, Km 5 vía a Puerto Colombia, A.A. 1569, Barranquilla (Colombia).
Fecha de recepción: 2 de septiembre de 2008
Fecha de aceptación: 25 de octubre de 2008
Resumen
En la patogénesis de las enfermedades alérgicas están involucrados el ambiente, la carga genética y la inmunocompetencia del individuo. Continuamente nuestro sistema inmune está expuesto a numerosas proteínas, sin embargo, solo unas pocas inducen una respuesta inmune alérgica. El potencial intrínsico de una proteína alergénica para inducir sensibilización solo se manifiesta en individuos susceptibles, genéticamente condicionados a presentar respuestas atópicas. Muchas de estas proteínas alergénicas comparten alguna homología en su secuencia de aminoácidos. Estos alérgenos poseen un amplio rango de características moleculares, ninguna de las cuales es única solo para estas proteínas alergénicas. A pesar de esto, algunas de estas características son más comunes entre algunos alérgenos que con otras proteínas. Se ha demostrado que algunas proteínas con actividad enzimática inducen reacciones alérgicas y que esta propiedad biológica está asociada con su actividad catalítica. En la presente revisión se describen las principales características moleculares de las proteínas alergénicas, y se hace énfasis en la cistein proteasas de los ácaros intradomiciliarios, en razón a que ellas son un factor de riesgo en el desarrollo de una respuesta inmune alérgica en individuos susceptibles y se constituyen en factores desencadenantes de respuestas inflamatorias en la fisiopatología de las enfermedades alérgicas respiratorias.
Palabras claves: Ácaros, alérgenos cistein proteasas, inflamación, biología computacional.
Abstract
The components involved in the pathogenesis of disorder allergic include the environment, the genetics and the immune competence. Continuously we are exposed to thousands of different proteins, but a single few of them induce an allergic immune response. The unique potential of an allergic protein to induce sensitization, only produce a response in susceptible individuals. Many of those allergic proteins share some homology in their sequence of amino acids, nevertheless, allergens have an ample rank of characteristic, no of which is unique for allergenic proteins. In spite of this, some characteristics are more common between allergens than in other proteins. It has been demonstrated that some proteins with enzymatic activity have a predisposition to induce allergic reactions and that its biological characteristics is associated with its catalytic activity. Those characteristic could be contribute directly to the allergenicity of this protein. In the present review we presented/displayed the main intrinsic characteristics of allergenic proteins associated with an allergic immune response, making emphasis in their Properties as activity cystein protease how and this biological faculty determining a factor in the development of the inflammatory cascade as an allergic immune response in susceptible individuals.
Key words: House dust mite, allergens, cystein protease, inflammation, computational biology.
INTRODUCCIÓN
La capacidad de ciertas proteínas para inducir respuestas alérgicas mediadas por Inmunoglobulina E en individuos susceptibles, las definen como alérgenos. Los síntomas inducidos por estas proteínas pueden ir desde una rinitis alérgica hasta un choque anafiláctico. Estas respuestas alérgicas son el resultado de complejas interacciones entre estas proteínas alergénicas y el sistema inmune, y existe una enorme dificultad para predecirlas. Sin embargo, está claro que algunas proteínas son mas alergénicas que otras. Esto ha generado un gran desafío entre los inmunólogos, al tratar de identificar las características que confieren a dichas proteínas el potencial para inducir una respuesta alérgica. (1)
La bioquímica de los alérgenos ha abierto una nueva era en el estudio 1de la biología estructural y de la proteomica, y se requieren herramientas sofisticadas para el procesamiento de los datos, debido a que cada vez más la abundancia de información estructural está permitiendo esclarecer la función biológica de los alérgenos y la asignación y clasificación de familias de proteínas. (2)
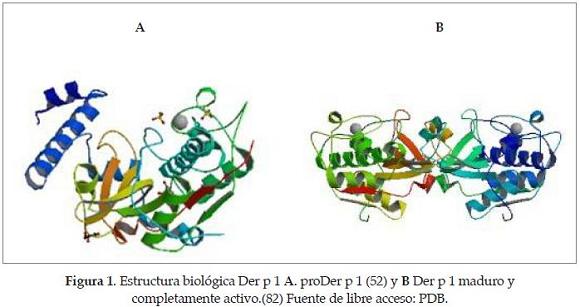
En los ácaros intradomiciliarios se han podido clasificar 21 familias de estas proteínas alergénicas, y las del grupo I son las más estudiadas hasta el momento. A ellas pertenece Der p 1, la cual es una glicoproteína de 25 kd (88). Ésta presenta una alta homología en su secuencia aminoacídica, con enzimas proteolíticas, cuya función depende de residuos de cisteína para su actividad catalítica. Se ha demostrado que las características moleculares de las cistein peptidasas están asociadas no solo a su estructura tridimensional, sino también a su estructura secundaria. La presencia de dominios altamente conservados en los miembros de esta gran familia puede hacer que estas proteínas alergénicas sean reconocidas por anticuerpos IgE de pacientes sensibilizados contra las cistein proteasas. Estos anticuerpos IgE específicos no solo nreconocen estos inmunógenos en múltiples especies de ácaros, sino también en distintas proteínas de orígenes muy diversos como kiwi, camarones y otros crustáceos, lo cual se debe, en parte, a la reactividad cruzada de determinantes alergénicos comunes entre las proteínas presentes en las especies comprometidas en la respuesta alérgica (89). En el presente trabajo presentamos las principales características moleculares y bioquímicas asociadas con la inducción de una respuesta inmune alérgica, y se hace énfasis en la función biológica del grupo de proteínas con actividad cistein proteasa que influencia directamente el desarrollo de las mismas.
En general, las proteínas alergénicas exhiben dos o tres propiedades moleculares distintas: la propiedad de sensibilizar, es decir, inducir al sistema inmune a producir una respuesta inmune adaptativa de anticuerpos IgE de alta afinidad; y la capacidad para provocar una respuesta alérgica, es decir, desencadenar síntomas y signos característicos de las enfermedades alérgicas en individuos previamente sensibilizados. Adicionalmente, un alérgeno tiene la propiedad de unir anticuerpos IgE específicos. Así un alérgeno es una proteína que comúnmente induce una respuesta inmune exagerada mediada por anticuerpos IgE en individuos genéticamente predispuestos. (3)
Los análisis de las bases de datos de proteínas sugieren que el universo de los alérgenos abarca más de 120 familias distintas de proteína (2). Muchas proteínas pueden tener actividad biológica que influencia la respuesta inmune. Sin embargo, la mayoría de estos alérgenos purificados no tienen efectos sobre la piel, tracto digestivo, nariz, etc. en individuos no alérgicos (8). Los alérgenos se derivan de proteínas con una variedad de funciones biológicas, incluyendo proteasas, proteínas de unión, proteínas estructurales, proteínas de transferencia de lípidos, profilinas, proteínas de unión a calcio, y otros. La función biológica de las enzimas proteolíticas de los ácaros del polvo intradomiciliario, influencian directamente el desarrollo de las respuestas IgE y pueden iniciar respuestas inflamatorias en el pulmón y otros órganos. Las características estructurales o biológicas pueden también influenciar el tiempo en el cual los alérgenos persisten en los espacios interiores o en el ambiente del exterior, así como en que conserven su alergenicidad en el tracto digestivo (15).
CARACTERÍSTICAS MOLECULARES DE LAS PROTEÍNAS ALERGÉNICAS
En la estructura molecular de las proteínas alergénicas se presentan regiones inmunodominantes, denominadas epítopes, las cuales interactúan con los fragmentos de unión al antígeno (Fab) de los anticuerpos IgE específicos. Los complejos inmunes Fab-alérgeno, tienen entre 15 a 22 residuos de aminoacídicos. De éstos sólo de 3 a 5 residuos contribuyen al proceso de unión a través de múltiples enlaces complementarios de tipo no covalente, originados por fuerzas electrostáticas, principalmente de tipo van der Waals. (11-13)
A la fecha se investiga ampliamente para determinar si las características intrínsecas de los epítopes se encuentran relacionadas con su capacidad alergénica. Es decir, si su complejidad molecular (secuencia de aminoácidos, estructura secundaria y tipo de plegamiento), así como su solubilidad, la estabilidad, el tamaño y la actividad bioquímica de un alérgeno, promueven las condiciones inmunológicas necesarias para la sensibilización del sistema inmunológico del huésped, la interacción con anticuerpos IgE y la inducción de las reacciones alérgicas (14).
Algunas de las relaciones establecidas hasta el momento entre las características moleculares de este tipo de proteínas y su capacidad alergénica, son las siguientes:
- Modificaciones postraduccionales
- Estabilidad de los alérgenos
- Alérgenos con actividad enzimática
- Transportadores de Ca+
- Alérgenos transportadores
- Profilinas
- Lectinas
1) Modificaciones postraduccionales
La mayoría de los alérgenos son proteínas extracelulares, que suelen sufrir modificaciones postraduccionales, y la principal es la glicosilación. Las glicoproteínas se forman en el retículo endoplásmico, donde se unen de manera covalente oligosacáridos, especialmente residuos de asparaginas (Nglicanos) de serinas/ treoninas (O-glicanos), o bien de prolinas y lisinas en las proteínas de origen vegetales. Estas reacciones se llevan a cabo gracias a la acción de glicosiltransferasas y de algunas glicosidasas. Las glicoproteínas presentan cambios en la estabilidad, la solubilidad, la hidrofobicidad y en la carga eléctrica, y ocasionan que los sitios glicosilados sean más visibles al sistema inmune del huésped (15).
Algunos alérgenos, N-glicosilados de plantas, son altamente inmunogénicos, poseen azúcares como manosa, fucosa, xilosa y Nacetilglucosamina, en un orden que no se encuentra en los glicanos de mamíferos, lo que podría ser un indicio de que este orden estructural y biológico precisamente es la clave de la alergenicidad (16-17).
2) Estabilidad de los alérgenos
Los principales alérgenos son proteínas termoestables y mantienen su alergenicidad después de ser sometidos a efectos físicos, tales como calentamiento. Otros son considerados incompletos por ser susceptibles al efecto de pH's bajos y a la acción de enzimas proteolíticas (18).
Los puentes disulfuro intramoleculares tienen un papel importante en la conservación de la alergenicidad. Trabajos realizados en Der f 1, Der f 2, Lep d 2 (alérgenos de ácaros del polvo casero) (19-20) muestran que al reducir estos puentes, o bien eliminarlos por mutagénesis dirigida, los alérgenos pierden su capacidad de unión a las moléculas de IgE específicas para alérgeno. Este detrimento en la alergenicidad se podría explicar esencialmente por la pérdida de la conformación original del alérgeno y por lo tanto de los epítopes conformacionales; o bien, a una menor estabilidad de los mismos, siendo más susceptibles a la digestión enzimática.
3) Alérgenos con actividad enzimática
Muchas de las proteínas alergénicas ejercen su función biológica a través de actividad enzimática y en su mayoría ellas son proteasas y nucleasas. Aunque no se ha comprobado que la alergenicidad de una proteína pueda ser atribuida únicamente a sus propiedades enzimáticas, se ha especulado que la actividad bioquímica de los alérgenos puede aumentar su inmunogenicidad. Es poco probable que su alergenicidad sea originada debido únicamente por esta actividad biológica (21-24).
Un ejemplo de lo anterior lo constituyen las proteasas del ácaro y la fosfolipasa A2 (principal alérgeno del veneno de abeja), las cuales inducen y modulan la síntesis de citocinas pro-inflamatorias al estimular directamente a células epiteliales bronquiales, mastocitos y basófilos, aumentando la cascada inflamatoria alérgica (25).
4) Transportadores de Ca+
Algunos alérgenos originarios de plantas y animales unen y transportan Ca2+. Estos presentan de 2 a 8 dominios denominados EF-hands o plegamientos tipo calmodulina, los cuales interactúan con diferentes ligandos dependientes de calcio (26). Se ha propuesto que una proteína alergénica puede asumir de manera reversible diferentes conformaciones, exponiendo epítopes variables que ocasionan interacciones diferentes con IgE de suero de pacientes, puesto a que el alérgeno unido al Ca2+ presenta una conformación abierta, haciendo que los residuos expuestos en la superficie sean accesibles a los anticuerpos; mientras que cuando el alérgeno está libre de calcio, éste presenta una conformación cerrada que impide la interacción con estas inmunoglobulinas (27).
5) Alérgenos transportadores
En esta categoría se encuentran las lipocalinas, proteínas altamente solubles que transportan retinol, esteroides, lípidos y ferohormonas (28).
Los principales alérgenos de este grupo provienen de animales domésticos comunes en ciudades industrializadas y en animales de granja; destacándose como aeroalérgenos Can f 1 y Can f 2 (perro), Mus m 1 (ratón), Rat n 1 (rata), Equ c 1 y Equ c 2 (caballo) y Bos d 2 (vaca), así como Bla g 4 (cucaracha) y Bos d 5 (beta-lactoglobulina de leche de vaca). Este último, presenta seis regiones de unión a los anticuerpos IgE específicos de suero de pacientes sensibilizados, y se destaca una secuencia altamente conservada en las lipocalinas rica en ácido aspártico (29).
6) Profilinas
Son proteínas de unión a la actina, aunque también interactúan con cadenas de poli L-prolina y con algunos fosfolípidos. Se ha encontrado que las áreas superficiales que contienen los epítopes de unión a IgE de este tipo de alérgenos, son las mismas que interactúan con actina, lo cual indica que las áreas expuestas a la superficie, que muestran afinidad por ligandos naturales, son también reconocidas por las regiones variables de estos anticuerpos (28).
7) Lectinas
Algunos alérgenos interactúan con carbohidratos. Ejemplos de esto lo encontramos en los alérgenos más importantes del látex del árbol del caucho, Hevea brasiliensis, tales como Hev b 6.01, Hev b 6.02 y Hev b 11, los cuales unen oligosacáridos de N-acetilglucosamina, gracias a un dominio de heveína. También se ha descrito este tipo de dominios en otras plantas, destacando Pers a 1 (aguacate), y algunos otros presentes en el plátano, kiwi y castaña, lo cual podría explicar el hecho personas sensibles a látex lo son también a estas frutas (30).
Alérgenos de los ácaros del polvo intradomiciliarios
Los ácaros son pequeños artrópodos pertenecientes a la clase Arácnida, poseen cuatro pares de patas, ausencia de segmentación del cuerpo y carencia de antenas. Se encuentran ditribuidos por todo el mundo, adaptados a vivir en diversos medios. Entre las numerosas especies se encuentran aquellos que viven en el polvo de las casas y basan su nutrición en detritus humano y animal (31).
Dermatophagoides pteronyssinus es la especie de ácaros intradomiciliarios más importante en Europa Occidental, Australia, Inglaterra y Nueva Zelanda. Dermatophagoides farinae prevalece en los Estados Unidos y Japón, aunque D. pteronyssinus prevalece en algunas regiones de estos países (76-77). La infestación con una u otra especie de ácaro depende no solo de la ubicación geográfica, sino también de las condiciones ambientales de cada hogar. El ácaro Blomia tropicalis es importante en regiones tropicales y subtropicales, particularmente en el cinturón del Trópico, Suramérica y el Sureste Asiático (78). Las especies del género Dermatophagoides y del género Blomia son evolutivamente dispares y presentan alguna reactividad cruzada en sus proteínas en razón a que tienen secuencias aminoacídicas disímiles (79).
Estos artrópodos son organismos complejos que producen miles de proteínas diferentes y otras macromoléculas. Se han caracterizado hasta el momento más de 21 grupos de alérgenos en nueve especies acarinas diferentes, y se han clasificado de acuerdo con su identidad bioquímica (32-33).
El primer alérgeno de ácaros descrito fue Der p 1 de Dermatophagoides pteronyssinus (16). Der f 1 fue después caracterizado por diversos investigadores (34-35). Der p 1 fue también el primer alérgeno con el que se desarrolló un análisis con cDNA (36). El análisis del cDNA reveló que ellas eran cistein proteasas de la misma familia de la papaína y actinidina (37-38).
El grupo 1 de alérgenos ha sido identificado como alérgenos mayores a partir de estudios con extractos alergénicos. Estos unen IgE de sujetos alérg6icos a los ácaros con alta frecuencia (16). Estas proteínas están presente en las heces de los ácaros en altas concentraciones. La actividad proteolítica de estas cistein proteasas se ha propuesto que aumentan su capacidad de sensibilizar seres humanos (39). La IgE dirigida contra Der p 1 se da en un rango del 50 - 70% en el suero de estos mismos pacientes alérgicos (40). Se ha confirmado una reactividad similar para Der f 1 (43).
La IgE específica anti - alérgenos de Dermatophagoides pteronyssinus representa entre el 6-21% del total de la IgE sérica polivalente (41-42).
Las cistein proteasas de ácaros intradomiciliarios y su potencial alergénico
Las proteasas se agrupan en clanes y familias. Los clanes son grupos de familias para quienes hay evidencia de ascendencia común. Las familias son agrupadas por su tipo catalítico: A, aspartato; C, cisteína; G, ácido glutámico; S, serina; T, treonina; y U, desconocido. Las serin, treonin y cistein proteasas utilizan en su sito activo un aminoácido nucleófilo y forman un estado intermediario acil y pueden también actuar fácilmente como transferasas (86).
La cistein proteasa del ácaro D. pteronyssinus (Der p 1) es una proteína de 25 Kda, codificado por un único gen que presenta frecuentes polimorfismos, los cuales han sido ya secuenciados (44). Tiene una variación prevalente de la secuencia alanin - valina en la posición 124 y aparecen variaciones esporádicas que difieren en 2 - 3 residuos (80-81).
El marco abierto de lectura codifica un péptido señal de 18 aa, un pro-péptido de 80 aa, y la región de la proteína madura que abarca 222 aa. La secuencia incluye cuatro sitios de potencial N-glicosilación, tres en la secuencia madura y uno en el pro-péptido. Der p 1 es producida como una enzima inactiva, que se activa después de la separación del propéptido. Aparte de inhibir la actividad de la pro-enzima, el pro-péptido puede también actuar como plataforma de plegamiento para la maduración de Der p 1, según lo sugerido para otras proteasas. Cuando Der p 1 se extrae de las heces del ácaro, está presente en la forma madura (nDer p 1) (45-46).
Las cistein proteasas se dividen en cinco clanes, cada una contiene a un número de familias, agrupadas con base en la arquitectura de su díada o triada catalítica. Der p 1 pertenece al clan CA, a la familia C1, que también incluye a la papaína y a sus parientes. Las estructuras cristalinas de la papaína, y de varias proteasas estrechamente relacionadas de esta misma familia, han sido determinadas, y los residuos catalíticos se han identificado como Cys, His, y Asn. Además, un residuo conservado de Gln es esencial para la actividad catalítica y se cree que ayuda a formar el agujero del oxianionico, el cual estabiliza el estado transitorio durante la catálisis (47).
Los residuos catalíticos Cys e His se piensa forman un Ion thiolato-imidazolium, estabilizado por un puente de hidrógeno directo entre las cadenas laterales de los residuos catalíticos His y Asn (48).
La mayoría de los miembros de la familia C1 tienen pro-péptidos homólogos al de la papaína (115 aa), aunque la longitud puede variar. Se estima que el pro-péptido actúa ocultando el sitio activo y blindándolo del acceso a los substratos, de tal modo que inhibe la actividad enzimática. La proteolisis del mismo es evitada ligando el pro-péptido en la orientación reversa comparada con la de un substrato. El pro-péptido de Der p 1 tiene solo 17 residuos idénticos al ser alineada su secuencia con el pro-péptido de la papaína (49).
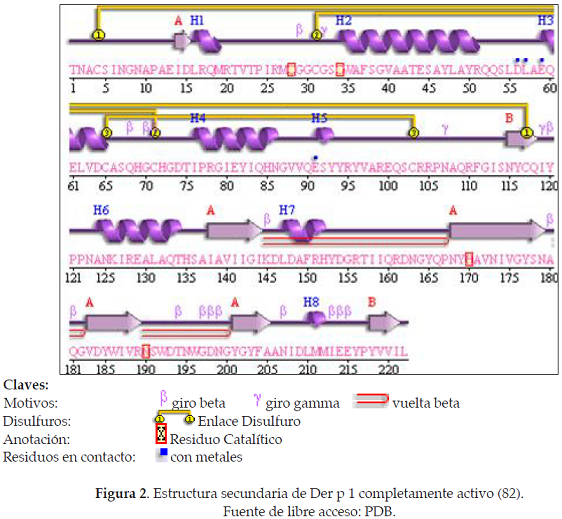
La estructura secundaria de la cistein proteasa del ácaro Dermatophagoides pteronyssinus está constituida en un 28% por hélices alfa (8 hélices; 63 residuos) y el 23% por láminas beta (18 hojas; 53 residuos) con diversos enlaces disulfuro que estabilizan la proteína (Ver Figura 2).
Epítopes moleculares de la proteína DER P 1
La región entre los aminoácidos 15-33 de la proteína madura es accesible a la interacción con paratopes de anticuerpos IgE, pero es relativamente extendida y podría constituir epítopes discontinuos con partes de la región entre los aminoácidos 188-199. La región 34-47, la cual contiene residuos de cisteína catalíticos, es una hélice enterrada en el interior de la molécula. El segmento comprende los residuos 52-111, está bien expuesta sobre la superficie y podría ser subdivida en pequeños segmento: 52-56, 60-80, 81-94, y 101-111, correspondiendo a epítopes lineales o epítopes discontinuos. El epítope 81-94 consiste de dos epítopes continuos separados por los aminoácidos 101-111, y este mismo es también expuesto a la superficie y constituye un epítope lineal ideal, al igual que el epítope 117-133. La región 155-175 está parcialmente enterrada en la interface de un dímero y puede ser parte de un epítope discontinuo con el segmento adyacente 176-187, o con parte del segmento 60-80. Finalmente, la región 188-199 es suficientemente compacta para constituir un epítope lineal o combinado con 15-33. Hay variaciones en todos los epítopes descritos comparados con Der f 1 y Eur m 1 con la excepción de la región 60-80, la cual está completamente conservada con respecto a Der f 1 (82).
Se han reportado por lo menos 12 isoformas de Der p 1 que han sido identificadas (90). La estructura tridimensional de esta proteína permite predecir el efecto de cada una de estas mutaciones sobre la estructura y los epítopes accesibles. La mayoría de los residuos específicos de cada isoforma son expuestos en la superficie de la proteína lejos del dímero de interface y sin consecuencias para la estructura de la proteína, con la excepción del Glu 44, uno de los residuos coordinados al ion de magnesio. Dos residuos, Ala 124 e Ile138, son más enterrados, pero el C-ß de Ala 124 está en ambiente relativamente hidrofóbico y el cambio por una Val podría favorecer la estabilidad de la proteína. Similarmente, el cambio de Ile por Met en la posición 138 es probablemente bien tolerada y/o incrementa la estabilidad de la proteína (82).
Los residuos aminoacídicos dentro del propéptido median su asociación a la membrana, y desempeñan un papel en el transporte de la proenzima a los lisosomas (50-51).
El grupo 1 de ácaros intradomiciliarios está constituido por las cistein proteasas de cada una de las diferentes especies de ácaros; estas proteínas provocan síntomas clínicos claros en >80% pacientes alérgicos a los ácaros. Se han caracterizado hasta el momento las cistein proteasas de Dermatophagoides pteronyssinus (Der p 1), Dermatophagoides farinae (Der f 1), Blomia tropicalis (Blo t 1), Dermatophagoides microceras (Der m 1) y Euroglyphus maynei (Eur m 1) (53). Entre ellas se destacan la cistein proteasa Der p 1, la cual es positiva en el 92% de 35 pacientes alérgicos a los ácaros por RAST (54), y Der f 1, que en 63 sueros de pacientes alérgicos a los ácaros, 55 (87%) tienen anticuerpos IgE contra Der f 1 (55).
Algunas cistein proteasas han sido implicadas en enfermedades humanas como cáncer,artritis reumatoide, osteoporosis y enfermedad de Alzheimer's (56). Por esta razón consideramos a las cistein proteasas de ácaros intradomiciliarios como moléculas blanco para el diseño de estrategias encaminadas al diagnóstico, tratamiento y control de enfermedades alérgicas (57).
Reactividad cruzada entre cistein proteasas
El fenómeno de reactividad cruzada ocurre cuando anticuerpos IgE, originalmente producidos para un alérgeno, se unen o reconocen un epítope en una proteína similar de un recurso diferente. La interacción con tales proteínas homólogas puede desencadenar reacciones alérgicas o puede ser completamente irrelevante para el paciente (58-60).
Las proteínas del grupo 1 de ácaros intradomiciliarios comparten características estructurales, que permite agruparlas dentro del clan CA, en la familia C1, que también incluye a la papaína, y catepsinas de mamíferos (B, C, F, H, L, K, O, S, V, X y W). Estas proteínas poseen dominios muy conservados (ver Figura 3) que hacen que estas proteínas alergénicas sean reconocidas por anticuerpos IgE de pacientes sensibilizados contra cistein proteasas de un recurso diferente. Es decir, estas proteínas presentan reactividad cruzada entre si, por lo que un grupo muy diverso de péptidos puede producir síntomas alérgicos comunes (61-63). Se ha comprobado que Der p I y Der f I tienen reactividad cruzada superior al 80 % (64-66).
El análisis de la secuencia aminoácidica de pro Der p 1 con la herramienta bioinformática WU-Blastp nos muestra una identidad con el alérgeno Eur m 1 del 83 %; con Der f 1 del 82%; con las cistein proteasa de Pseroptes ovis del 61%; Sarcoptes scabiei del 44%; con Blo t 1 del 38%; con Sui m 1 del 34%; con la Catepsina L de Leishmania brazilensis 32%; Catepsina L2 de Fasciola hepatica del 31%; con una proteína no caracterizada del arroz, Oriza sativa Subsp, del 32%; con el camarón rojo del norte, 33%, etc. Esto podría explicar por qué pacientes con alergia a ácaros pueden presentar y/o aumentar la severidad a la alergia al camarón y caracol por reactividad cruzada.
Entre otros péptidos relevantes con reactividad cruzada con el grupo 1 de ácaros intradomiciliarios tenemos a las catepsinas, un grupo de exopeptidasas lisosomales presentes en todos los tejidos humanos involucrada en la degradación de proteínas, procesamiento de antígenos, en activación de proenzimas y en procesos apoptóticos (67-68).
y las cathepsinas de plantas, las cuales son utilizadas en la movilización de proteínas de reserva (69-71).

Respuesta inmune a las cistein proteasa
Aunque hay una comprensión creciente de los mecanismos implicados en el desarrollo de la inflamación alérgica, una vez la sensibilización ha ocurrido, los mecanismos de la interacción entre Der p 1 y las poblaciones celulares de la vía aérea y su papel en el proceso de sensibilización siguen siendo confusos (83).
El epitelio bronquial es la primera barrera que encuentran los antígenos inhalados; ésta proporciona un acoplamiento importante entre el medio externo y el interior del cuerpo. La activación de células dendríticas por los antígenos inhalados conduce normalmente a la inducción de la tolerancia de la inhalación más que a la inflamación alérgica. Los mecanismos implicados en la interrupción de esta tolerancia en individuos alérgicos se desconocen. Se ha demostrado que una corta exposición a altas concentraciones de Der p 1, o la exposición prolongada a bajas concentraciones, causó degradación de las uniones entre células epiteliales bronquiales en pacientes asmáticos y también en las células de individuos normales. Esto incrementa la permeabilidad del epitelio, lo cual podría favorecer la penetración en la mucosa de la vía aérea, no solo de Der p 1, sino también de otros antígenos desprovistos de actividad proteolítica, de tal modo que aumenta la probabilidad de encontrar células presentadora de antígenos y causar sensibilización. Tal fenómeno explicaría, por qué la sensibilización contra los antígenos del ácaro del polvo doméstico se asocia con frecuencia a la sensibilización con múltiples antígenos (84).
La exposición de las células epiteliales de la vía respiratoria a Der p1 induce la secreción de citocinas pro inflamatorias, particularmente GM-CSF, e IL -6. La secreción de estos mediadores de las células epiteliales bronquiales de pacientes asmáticos se asocia al transporte intracelular del alérgeno y ocurre después de una breve exposición a concentraciones bajas de Der p1 (85). De esta forma la actividad catalítica cistein-proteasa del alérgeno de ácaro de polvo casero Der p 1, se asocia con el incremento de la permeabilidad en el epitelio bronquial, facilitando así su propio procesamiento.
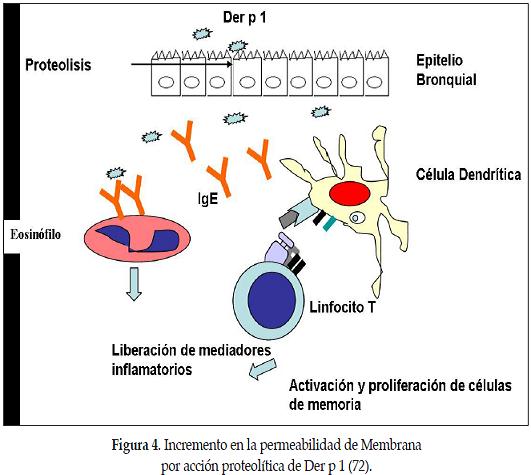
Este alérgeno es capaz también de clivar a CD23 (receptor de baja afinidad para IgE, el cual regula la síntesis de estas inmunoglobulinas), y a CD25 (la subunidad del receptor para interleucina-2). Como resultado del clivage del CD25 de superficie, los linfocitos T de sangre periférica disminuyen su proliferación y la secreción IFN-γ como respuesta a una estimulación. Las poblaciones de la célula Th1 y Th2 promueven el desarrollo de células del mismo subconjunto, mientras que suprime la propagación de los del otro subconjunto. Por lo tanto, el clivaje inducido por Der p 1 de CD25, probablemente conduce al crecimiento deteriorado de las células del subconjunto Th1 y al consiguiente aumento de los del subconjunto Th2 (73). Esta subpoblación de células T produce un perfil de citoquinas que incluyen IL-4, IL-5, IL-6, e IL-13, las cuales ejercen sus efectos sobre los mismos linfocitos T, pero también sobre otro tipo de células, incluyendo linfocitos B y células presentadoras de antígenos, tales como monocitos y células dendríticas. La IL-4 sirve como factor de crecimiento de los linfocitos B e induce la producción de los isótipos IgE e IgG4. En la exposición inicial al alérgeno, los linfocitos T son estimulados, y la producción de IgE es inducida. Estos anticuerpos se unen a los receptores de la superficie mastócitos (74-75).
REFERENCIAS
1. Russell DJ, et al. Why are some proteins allergens? Toxicological Sciences 2000; 55: 235-246. Abstract/Free Full Text [ Links ]
2. Chapman MD, et al. Nomenclature and structural biology of allergens J Allergy Clin Immunol 2007;119:414-20. [ Links ]
3. Alberse RC. Structural biology of allergens. J Allergy Clin Immunol 2000;106:228-238. Abstract [ Links ]
4. Marsh DG, Goodfriend L, King TP, et al. Allergen nomenclature. Bull WHO 1986; 64:767. Abstract [ Links ]
5. King TP, Hoffman D, Lowenstein H, et al. allergen nomenclature. WHO/IUIS Allergen Nomenclature Committee. Int Arch Allergy Immunol 1994; 105: 224. Abstract [ Links ]
6. Ishizaka K, Ishizaka T, Honbrook MM. Physico-chemical properties of human reaginic antibody. IV. Presence of a unique immunoglobulin as a carrier of reaginic activity. J Immunol 1966; 97: 75. Abstract [ Links ]
7. Ishizaka K, Ishizaka T. Mechanisms of reaginic hypersensitivity and IgE antibody response. Immunol Rev 1978; 41: 109. Abstract [ Links ]
8. Marsh DG, Goodfriend L, King TP, et al. Allergen nomenclature. Bull WHO 1986; 64:767. Abstract [ Links ]
9. Wide L, Bennich H, Johansson SGO. Diagnosis of allergy by an in vitro test for allergen antibodies. Lancet 1967; 2: 1105. Abstract [ Links ]
10. Platts-Mills TAE, Snajdr MJ, Ishizaka K, et al. measurement of IgE antibody by an antigenbinding assay: correlation with PK activity and IgG and IgA antibodies to allergens. J Immunol 1978; 120: 1201. Abstract [ Links ]
11. Woodfolk JA, Wheatley LM, Piyasena RV, et al. Trichophyton antigens associated with IgE antibodies and delayed type hypersensitivity: sequence homology to two families of serine proteinases. J Biol Chem 1998; 273: 29489. Abstract/Free Full Text [ Links ]
12. Schellenberg RR, Adkinson NF. Measurement of absolute amounts of antigen-specific human IgE by a radioallergosorbent test (RAST) elution technique. J Immunol 1975; 115: 1577. Abstract [ Links ]
13. Bohle B, Schwihla H, Hu HZ, et al. long lived Th2 clones specific for seasonal and perennial allergens can be detected in blood an skin by their TCR-hypervariable regions. J Immunol 1998; 160: 2020. Abstract/Free Full Text [ Links ]
14. Ring J, Kramer U, Schafer T et al. Why are allergies increasing? Curr Opin Immunol 2001; 13: 701-708. Abstract [ Links ]
15. Pómes A. Intrinsic properties of allergens and environmental exposure as determinants of allergenicity. Allergy 2002; 57: 673-679. Abstract [ Links ]
16. Fötisch F, Vieths S. N-and O-linked oligosaccharides of allergenic glycoproteins. Glycoconj J 2001; 18: 373-390. Abstract [ Links ]
17. Van Ree R. Carbohydrate epitopes and their relevance for the diagnosis and treatment of allergic diseases. Int Arch Allergy Immunol 2002; 129: 189-97. Abstract [ Links ]
18. González EM, Villalba M, Lombardero M et al. Influence of the 3D-conformation, glycan component and micro-heterogeneity on the epitope structures of Ole e 1 the major olive allergen. Use of the recombinant isoforms and specific monoclonal antibodies as immunological tools. Mol Immunol 2002; 39: 93-101. Abstract [ Links ]
19. Taylor SB, Lenher SL. Principles and characteristics of food allergens. Crit Rev Food Sci Nutr 1996; 36: S91-S118. Abstract [ Links ]
20. Brederhost R, Kerstin D. What establishes a protein as an allergen? J Chromatogr B Biomed Sci Appl 2001; 756: 33-40. Abstract [ Links ]
21. Roitt I, Brostoff J, Male D. Immunology. 5th edition. UK: Mosby International; 1998: 107- 119. [ Links ]
22. Holgate S, Church M, Lichtenstein L. Alergia. 2a edición. España: Editorial Harcout; 2001: 3-16. [ Links ]
23. Sela M, Pecht I. The nature of the antigen. Adv Protein Chem 1996; 49: 289-328. Abstract [ Links ]
24. Valenta R. The future of antigen-specific immunotherapy of allergy. Nat Rev Immunol 2002; 2: 446-453. Abstract [ Links ]
25. Buchanan BB, Adamidi C, Lozano MR, et al. Thioredexinlinked mitigation of allergic response to wheat. Proc Natl Acad Sci 1997; 94: 5372-77. Abstract [ Links ]
26. Valenta R. Recombinant allergen molecules: tools to study effector cell activation Immunol Rev 2001; 179: 119-127. Abstract. [ Links ]
27. Virtanen T. Lipocalis allergens. Allergy 2001; 56: 48-51. Abstract [ Links ]
28. Tinghino R, Twardosz A, Barletta B, et al. Molecular, structural, and immunologic relationships between different families of recombinant calcium-binding pollen allergens. J Allergy Clin Immunol 2002; 109: 314-20. Abstract [ Links ]
29. Wal JM. Structure and function of milk allergens. Allergy 2001; 56: 35-38. Abstract [ Links ]
30. Breiteneder H, Ebner C. Molecular and biochemical classification of plant-derived food allergens. J Allergy Clin Immunol 2000; 106: 27-36. Abstract [ Links ]
31. Sánchez-De la Vega W, Sánchez-Medina M, Lucena G, Vanella L, Moreno G, Albornoz A, et al. Manual Latinoamericano de Alergia e Inmunología. Bogotá (Colombia): Colciencias; 1987: 39-60. [ Links ]
32. Fernández-Caldas E, Puerta L, et al. Mite allergens. Clin Allergy Immunol. 2004; 18:251- 70. Abstract [ Links ]
33. International Union of Immunological societies (IUIS) Allergen Nomenclature Subcommittee. Allergen Nomenclature. www.allergen.org [ Links ]
34. Heymann PW, Chapman MD, Platts-Mills TAE. Antigen Der f I from the house dust mite Dermatophagoides farinae. In structural comparison with Der p I from Dermatophagoides pteronyssinus and human IgE epitope specificity of murine IgG and human IgE antibodies. J Immunol 1986; 137:2841- 2847. Abstract [ Links ]
35. Yasueda H, et al. Isolation and characterization of two allergens from Dermatophagoides farinae. Int Arch Allergy Appl Immunol 1986; 81: 214- 223. Abstract [ Links ]
36. Thoma WR, et al. Cloning and expression of DNA coding for the major house dust mite allergen Der p 1 in E. coli. Int Arch Allergy Appl Immunol 1986; 85:127-129. Abstract [ Links ]
37. Chua KY, et al. Sequence analysis of cDNA coding for a major house dust mite allergen, Der p I. Homology with cystine protease. J Exp Med 1988;167:175-182. Abstract [ Links ]
38. Dilworth RJ, et al. Sequence analysis of cDNA coding for a major house dust mite allergen. Der f I. Clin Exp Allergy 1991; 21:25-32. Abstract [ Links ]
39. Schulz O, H. F. Sewell, F. Shakib. The interaction between the dust mite antigen Der p 1 and cell-signalling molecules in amplifying allergic disease. Clin Exp Allergy 1999; 29:439.- 444. Medline [ Links ]
40. Lind P, Lowenstein H. Identification of allergens in Dermatophagoides pteronssinus mite body extract by crossed radioimmunoelectr ophoresis with two different rabbit antibody pools. Scand J immunol 1983; 17:263-273. Abstract [ Links ]
41. Lind WR, et al. Purification and partial characterization of two major allergens from the house dust mite Dermatophagoides pteronyssinus. J Allergy Clin Immunol 1988; 76:753-761. Abstract [ Links ]
42. Vand der ee JS, et al. Skin tests and histamine release with P1-depleted Dermatophagoides pteronyssinus body extracts and purified P1. J Allergy Clin Immunol 1988; 81:884-895. Abstract [ Links ]
43. Lind P, Hansen OC, Horn N. The binding of mouse hybridoma and human IgE antibodies to the major fecal allergen, Der p 1, of Dermatophagoides pteronyssinus. Relative binding site location and species specificity studied by solid phase inhibiton assays with radiolabeled antigen. J Immunol 1988; 140: 4256-62. Abstract [ Links ]
44. Chua K Y, Stewart G A, Thomas W R, Simpson R J, Dilworth R J, Plozza T M, Turner K J. Sequence analysis of cDNA coding for a major house dust mite allergen, Der p 1: homology with cysteine proteases. J Exp Med 1988; 167:175.-182. Abstract [ Links ]
45. Vernet T, H E Khouri, P Laflamme, D C Tessier, R Musil, B J Gour-Salin, A C Storer, D Y Thomas. Processing of the papain precursor: purification of the zymogen and characterization of its mechanism of processing. J Biol Chem 1991; 266:21451.-21457. Abstract/Free Full Text [ Links ]
46. Ikemura, H, H Takagi, M Inouye. Requirement of pro-sequence for the production of active subtilisin E in Escherichia coli. J Biol Chem 1987; 262:7859.-7864. Abstract/Free Full Text [ Links ]
47. Menard, R, J Carriere, P Laflamme, C Plouffe, H E Khouri, T Vernet, D C Tessier, D Y Thomas, A C Storer. Contribution of the glutamine 19 side chain to transition-state stabilization in the oxyanion hole of papain. Biochemistry 1991; 30:8924.-8928. Medline [ Links ]
48. Storer A C, Menard R. Catalytic mechanism in papain family of cysteine peptidases. Methods Enzymol 1994; 244:486.-500. Medline [ Links ]
49. Barrett A J, N D Rawlings, J F Woesser. Cysteine peptidases. Handbook of Proteolytic Enzymes. London: Academic Press; 1998. P. 543-798. [ Links ]
50. Schultz, A. et al. SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci 1998; USA 95, 5857-5864. Abstract [ Links ]
51. Letunic et al. SMART 5: domains in the context of genomes and networks Nucleic Acids Res 2006; 34, D257-D260. Abstract [ Links ]
52. Meno K, Thorsted PB, Ipsen H, Kristensen O, Larsen JN, Spangfort MD, Gajhede M, Lund K. The crystal structure of recombinant proDer p 1, a Major house dust mite proteolytic. J Immunol 2005; 175: 3835-3845. Abstract [ Links ]
53. International Union of Immunological societies (IUIS) Allergen Nomenclature Subcommittee. Allergen Nomenclature.http://www.allergen.org [ Links ]
54. Chapman MD, Platts-Mills TA. Purification and characterization of the major allergen from Dermatophagoides pteronyssinus-antigen P1. J Immunol 1980 Aug; 125(2):587-92. Abstract [ Links ]
55. Heymann PW, Chapman MD, Platts-Mills TA. Antigen Der f I from the dust mite Dermatophagoides farinae: structural comparison with Der p I from Dermatophagoides pteronyssinus and epitope specificity of murine IgG and human IgE antibodies. J Immunol 1986 Nov 1;137(9):2841-7. Abstract [ Links ]
56. Furmonaviciene R, Shakib F. The molecular basis of allergenicity: comparative analysis of the three dimensional structures of diverse allergens reveals a common structural motif. Mol Pathol 2001 Jun;54(3):155-9. Abstract [ Links ]
57. Furmonaviciene R, Sewell HF, Shakib F. Comparative molecular modelling identifies a common putative IgE epitope on cysteine protease allergens of diverse sources. Clin Exp Allergy 2000 Sep;30(9):1307-13. Abstract [ Links ]
58. Woodfolk JA, Wheatley LM, Piyasena RV, et al. Trichophyton antigens associated with IgE antibodies and delayed type hypersensitivity: sequence homology to two families of serine proteinases. J Biol Chem 1998; 273: 29489. Abstract [ Links ]
59. Schellenberg RR, Adkinson NF. Measurement of absolute amounts of antigen-specific human IgE by a radioallergosorbent test (RAST) elution technique. J Immunol 1975; 115: 1577. Abstract [ Links ]
60. Bohle B, Schwihla H, Hu HZ, et al. long lived Th2 clones specific for seasonal and perennial allergens can be detected in blood an skin by their TCR-hypervariable regions. J Immunol 1998; 160: 2020. Abstract [ Links ]
61. Goodman RE. Practical and predictive bioinformatics methods for the identification of potentially cross-reactive protein matches. Mol Nutr Food Res 2006 Jul;50(7):655-60. Abstract [ Links ]
62. Thomas WR, Smith WA, Hales BJ. The allergenic specificities of the house dust mite. Chang Gung Med J 2004 Aug;27(8):563-9. Abstract [ Links ]
63. Simpson A, Green R, Custovic A, Woodcock A, Arruda LK, Chapman MD. Skin test reactivity to natural and recombinant Blomia and Dermatophagoides spp. allergens among mite allergic patients in the UK. Allergy 2003; Jan;58(1):53-6. Abstract [ Links ]
64. Pauli G, Bessot JC. The fight against mites. Allergy Today 1998;2(7):7-9. Abstract [ Links ]
65. Fernández E, Puerta L, Mercado D, Lockey RF, Caraballo L. Mite fauna Der p1, Der f1 and Blomia tropicalis allergen in a tropical environment. Clin Exp Allergy 1993; 23(4):2992-7. Abstract [ Links ]
66. Carswell F. House dust allergy. ACI Int 1996;8(5-6):169-71. Abstract [ Links ]
67. Thomas WR, Hales BJ, Smith WA. Recombinant allergens for analysing T-cell responses. Methods. 2004 Mar;32(3):255-64. Abstract [ Links ]
68. Aalberse RC. Structural features of allergenic molecules. Chem Immunol Allergy. 2006;91:134-46. Abstract [ Links ]
69. Raina OK, Yadav SC, Sriveny D, Gupta SC. Immuno-diagnosis of bubaline fasciolosis with Fasciola gigantica cathepsin-L and recombinant cathepsin L 1-D proteases. Acta Trop 2006 May;98(2):145-51. Abstract [ Links ]
70. Collazo ME, Diaz AM. Monoclonal antibodies against whole body extract of the dust mite Blomia tropicalis. P R Health Sci J. 2003 Dec; 22(4):345-51. Abstract [ Links ]
71. Rokni MB, Massoud J, Hanilo A. Comparison of adult somatic and cysteine proteinase antigens of Fasciola gigantica in enzyme linked immunosorbent assay for serodiagnosis of human fasciolosis. Acta Trop. 2003 Sep;88(1):69-75. Abstract [ Links ]
72. Shakib F, Schulz O, Sewell HF. A mite subversive: cleavage of CD23 and CD25 by Der p 1 enhances allergenicity. Immunol Today 1998; 19:313. Abstract [ Links ]
73. Abbas, A.K., Murphy K.M., Sher A. Functional diversity of helper T lymphocytes. Nature 1996;383:787-793. Abstract [ Links ]
74. Schulz O, Sewell HF, Shakib F. Proteolytic cleavage of CD25, the a subunit of the human T cell Interleukin 2 receptor, by Der p 1, a major mite allergen with cysteine protease activity. J Exp Med 1998; 187:271-5. Abstract [ Links ]
75. Machado DC, Horton D, Harrop R et al. Potential allergens stimulate the release of mediators of the allergic response from cells of mast cell lineage in the absence of sensitization with antigen-specific IgE. Eur J Immunol 1996; 26:2972-80. Abstract [ Links ]
76. Weghofer M, Thomas WR, Pittner G, Horak F, Valenta R, Vrtala S. Comparison of purified Dermatophagoides pteronyssinus allergens and extract by two-dimensional immunoblotting and quantitative immunoglobulin E inhibitions. Clin Exp Allergy 2005 Oct;35(10):1384-91. Abstract [ Links ]
77. Arlian LG, Bernstein D, Bernstein IL, Friedman S, Grant A, Lieberman P, Lopez M, Metzger J, Platts-Mills T, Schatz M, et al. Prevalence of dust mites in the homes of people with asthma living in eight different geographic areas of the United States. J Allergy Clin Immunol 1992 Sep;90(3):292-300. Abstract [ Links ]
78. Fernández-Caldas E, Lockey RF. Blomia tropicalis, a mite whose time has come. Allergy 2004 Nov;59(11):1161-4. Abstract [ Links ]
79. Cheong N, et al. Lack of human IgE crossreactivity between mite allergens Blo t 1 and Der p 1. Allergy 2003; 58:912-920. Abstract [ Links ]
80. Smith WA, et al. Allergens of wild house dust mites: environmental Der p 1 and Der p 2 sequence polymorphisms. J Allergy Clin Immunol 2001; 107:285-992. Abstract [ Links ]
81. Jarnicki AG, et al. Stimulatory and inhibitory epitopes in the T cell responses of mice to Der p 1. Clin Exp Allergy 2002; 32:942-950. Abstract [ Links ]
82. De Halleux S, Stura E, VanderElst L, Carlier V, Jacquemin M, Saint-Remy JM. Threedimensional structure and IgE-binding properties of mature fully active Der p 1, a clinically relevant major allergen J.Allergy Clin Immunol 2006; 117: 571-576. Abstract [ Links ]
83. Hong W, Winton HL, Soeller C, Tovey E, Gruenert DC, Thompson PJ, Stewart GA, Taylor GW, Garrod DR, Cannell MB, Robinson C. Der p 1 facilitates transepithelial allergen delivery by disruption of tight junctions. J Clin Invest 1999 jul;104:123-133. Full Text [ Links ]
84. Winton W, Soeller, Gruenert, Thompson, Cannell, Stewart, Garrod, Robinson. Quantitative structural and biochemical analyses of tight junction dynamics following exposure of epithelial cells to house dust mite allergen Der p 1. Clinical & Experimental Allergy 2000;30 (5): 685 698. Abstract [ Links ]
85. Tomee JFC, van Weissenbruch R, de Monchy JGR, Kauffman HF. Interactions between inhalant allergen extracts and airway epithelial cells: effect on cytokine production and cell detachment. J Allergy Clin Immunol 1998; 102:75-85. [ Links ]
86. Barrett AJ, Rawlings ND. Evolutionary lines of cysteine peptidases. Biol Chem 2001 May;382(5):727-33. [ Links ]
87. Marchler-Bauer A, Bryant SH. CD-Search: protein domain annotations on the fly. Nucleic Acids Res 2004; 32: 27-331. Abstract [ Links ]
88. Chua KY, Stewart GA, Thomas WR, Simpson RJ, Dilworth RJ, Plozza TM, et al. Sequence analysis of cDNA coding for a major house dust mite allergen, Der p 1: homology with cysteine proteases. J Exp Med1988;167:175-82. [ Links ]
89. Simpson A, Green R, Custovic A, Woodcock A, Arruda LK, Chapman MD. Skin test reactivity to natural and recombinant Blomia and Dermatophagoides spp. allergens among mite allergic patients in the UK. Allergy. 2003 Jan;58(1):53-6. [ Links ]
90. Smith WA, Hales BJ, Jarnicki AG, Thomas WR. Allergens of wild house dust mites: environmental Der p 1 and Der p 2 sequence polymorphisms. J Allergy Clin Immunol 2001;107:985-92. [ Links ]

