










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Salud Uninorte
versão impressa ISSN 0120-5552versão On-line ISSN 2011-7531
Salud, Barranquilla v.27 n.1 Barranquilla jan./jun. 2011
Investigación científica sobre genotipificación y distribución de Giardia intestinalis en humanos y caninos de América
Scientific research about genotyping and distribution of Giardia intestinalis in humans and dogs from America
Giovanny Torres Lindarte1, Mario Zapata Tamayo2, Marcos Restrepo Isaza3, Leonardo Ríos Osorio4
1 Candidato a Maestría en Microbiologia y Bioanálisis. Grupo de Investigación Instituto Colombiano de Medicina Tropical. Universidad CES. Sabaneta (Antioquia). gtorres@ces.edu.co2 Magister en Ciencias Básicas Biomédicas. Especialista en Epidemiología. Grupo de Investigación Microbiología Veterinaria, Escuela de Microbiología Universidad de Antioquia. Medellín (Antioquia).
3 Especialista en Parasitología y en Medicina de laboratorio. Grupo de Investigación Instituto Colombiano de Medicina Tropical. Universidad CES. Sabaneta (Antioquia).
4 Bacteriólogo y Lab. Clínico. Especialista en CBB, Parasitología Humana. Ph.D. en Sostenibilidad. Grupo de Investigación Microbiología Veterinaria, Escuela de Microbiología, Universidad de Antioquia. Medellín (Colombia).
Correspondencia: Giovanny Torres Lindarte, Instituto Colombiano de Medicina Tropical- Universidad CES, carrera 43a 52 sur 99, Sabaneta (Antioquia), telefono +57(4)3053500. gtorres@ces.edu.co
Resumen
Objetivo: Describir la distribución geográfica en América de los genotipos A, B, C y D de Giardia intestinalis hallados en humanos y en perros, y los marcadores y técnicas usadas, de acuerdo con la literatura científica.
Materiales y métodos: Revisión sistemática en la que se seleccionaron 19 artículos, y en todos se caracterizaron aislados de humanos; solo 10 de estos caracterizaron aislados de perros.
Resultados: México fue el país que más estudios realizó (31,6%). El genotipo A fue reportado en el 89,5% de los trabajos que analizaron aislados de humanos y el genotipo B en el 52,3%. En los que analizaron aislados de perros, el genotipo A se informó en el 60%, el genotipo B en el 20%, el genotipo C en el 30% y el genotipo D en un 20%. La técnica y el marcador más usados fueron la PCR-RFLP y el gen de fi-Giardin.
Conclusiones: El genotipo A es el responsable de la mayoría de infecciones en humanos como perros de América, aunque los genotipos B, C y D también fueron identificados. El hallazgo de perros infectados con genotipos zoonóticos sustentan su papel como fuentes potenciales de infección de G. intestinalis para humanos.
Palabras clave: Giardia lamblia, genotipo, reacción en cadena de la polimerasa, zoonosis, localization geográfica de riesgo.
Abstract
Objetive: To describe America geographical distribution of A, B, C and D Giardia intestinalis genotypes founded in humans and dogs, and molecular markers and techniques used to detect them, accordingly to scientific literature.
Materials and methods: Systematic revision was made it and 19 scientific articles were selected in which Giardia intestinales human isolates were characterized; 10 of this articles reported dog isolates.
Results: 31,6% of the studies were made in Mexico. A genotype in 89,5% and B genotype in 52,3% were reported in studies that analyzed human isolates, in front of, A genotype in 60%, B genotype in 20%, C genotype in 30% and D genotype in 20% that were reported in analyzed dog studies. PCR-RFLP was the most used molecular technique, and fi-Giardin gen was the most used molecular marker.
Conclusions: A genotype was responsible of both humans and dogs Giardia intestinalis infections in America; and B, C and D genotype were identified in infections, too. Findings of infected dogs with zoonotic genotypes support their role as a potential source for human infection with G. intestinalis.
Key words: Giardia lamblia, Genotype, Polymerase Chain Reaction, Zoonoses, Geographical Localization of Risk.
introducción
Giardia intestinalis es un protozoo flagelado que infecta un amplio rango de hospederos vertebrados, entre los que se encuentran los humanos, perros y otros animales, causando en ellos la enfermedad conocida como giardiasis (1). Esta parasitosis es una de las principales causas de gastroenteritis epidemica a nivel mundial, considerada como una enfermedad infecciosa reemergente y un problema de salud pública (2,3)
En Asia, África y Latino América cerca de 200 millones de personas presentan síntomas y en los países desarrollados Giardia intestinalis es el parásito intestinal más común encontrado en humanos (4, 5). La prevalencia en países industrializados puede ir desde un 2% hasta un 5% y exceder el 30% en países en vía de desarrollo (6). La prevalencia reportada en Colombia esta alrededor del 12% en población general y del 28% en población de 1 a 4 anos (7). En los perros es el protozoo más común encontrado con prevalencias alrededor del 10% en perros adultos bien cuidados, del 36 al 50% en cachorros y hasta el 100% en animales de criaderos (8).
Giardia fue inicialmente descrita por van Leeuwenhoek en 1681 (9), pero fue hasta 1859 cuando Lambl describió con mayor detalle el microorganismo (10). Desde entonces se describieron más de 40 especies basándose en el huésped donde se encontró el parásito (11). Hacia 1952 Filice propuso tres especies apoyándose en las características morfológicas: G. lamblia (humanos y otros mamíferos), G. muris (roedores) y G. agilis (anfibios) (12). Entre las décadas de los anos ochenta y los noventa se describieron las otras tres especies reportadas basándose en criterios morfológicos detectados por microscopía electrónica: G. psittaci (pericos), G. ardae (garzas) y G. microti (roedores) (13-15).
Como G. intestinalis (sinónimo: G. lamblia o G. duodenalis) es la única especie capaz de infectar a los humanos, así como a una variedad de mamíferos, es la especie con mayor importancia dentro del género, a tal punto que la Organización Mundial de la Salud (OMS) desde los anos veinte le ha reconocido su potencial zoonótico (16). A través de una variedad de herramientas moleculares que incluyen el análisis de isoenzimas, Reacción en Cadena de la Polimerasa con análisis del polimorfismo de longitud de los fragmentos de restricción (PCR-RFLP), tipificación de cromosomas por electroforesis de campo pulsado y secuenciación de genes constitutivos, se ha mostrado que G. intestinalis es un complejo de especies que comprende al menos siete genotipos (17-19).
Los principales marcadores que han sido utilizados para la genotipificación son: la subunidad pequena del RNA ribosomal (SSu-rRNA), el gen |3-giardin (bg), la glutamato deshidrogenasa (gdh), la trifosfato isomerasa (tpi) y el factor de elongación 1-alfa (ef-1) (20). El análisis de estos marcadores les permitió a los investigadores clasificar inicialmente las poblaciones de G. intestinalis de la siguiente manera: en América como grupo 1, 2 y 3 (11,12), en Europa como Polish y Belgian (13,14) y en Australia como genotipos A y B (21). Otros estudios permitieron conocer genotipos adicionales que fueron clasificados como C, D, E, F y G (15, 22); además se logró identificar diferencias dentro de los genotipos A y B, que fueron clasificados posteriormente como subgenotipos: AI, AII, BIII y BIV (23).
Aunque toda esta clasificación puede llegar a ser confusa y algunas formas de clasificar pueden resultar limitadas y poco funcionales cuando son aplicadas en diferentes estudios, la literatura reciente demuestra que la clasificación que arroja más información acerca de las poblaciones de G. intestinalis, sobre todo si se quieren realizar estudios de epidemiología molecular, es: A (AI, AII), B (BIII, BIV), C, D, E, F y G (24). Como son los genotipos A y B los encontrados infectando humanos y un amplio rango de hospederos, entre los que se encuentran los perros, se han considerado como los genotipos potencialmente zoonóticos, en especial el subgenotipo AI (1). Los genotipos restantes (C-G) se consideran huésped-específicos (25).
La transmisión de G. intestinalis siempre se ha asociado a condiciones higiénicosanitarias deficientes en los países en vía de desarrollo (26) y en los países desarrollados, donde las condiciones higiénicas son favorables, la transmisión se ha relacionado principalmente con fuentes de agua no tratadas (27). Sin embargo, los investigadores que respaldan la transmisión zoonótica de G. intestinalis argumentan que los animales también juegan un papel importante en la propagación del parásito.
De acuerdo con lo anterior, se plantea una posible transmisión cruzada debido al estrecho contacto que puede existir entre animales y humanos, como las mascotas, especialmente los perros (28). Estos animales se han convertido en parte integral de la familia y usualmente son mantenidos dentro de la casa, viviendo en contacto frecuente con los humanos, tanto así, que algunos llegan a compartir la cama con sus amos (29).
Esta cercanía no está libre de riesgos, ya que un perro infectado por el parásito, sin importar las condiciones higiénicosanitarias, puede actuar como fuente de infección para el humano, contaminando con materia fecal, que pueda estar en su hocico y pelaje, objetos o superficies con las que estará en contacto el humano (1).
Cualquier persona es susceptible de infectarse, debido a que esta parasitosis es adquirida por vía fecal-oral, pero son los ninos, justamente por su naturaleza curiosa, los que están en mayor riesgo de infectarse (29, 30). Estudios realizados en diferentes partes del mundo han proporcionado evidencia acerca de la trasmisión zoonótica entre perros y humanos; es el caso de dos estudios realizados, uno en India y el otro en Tailandia, donde se encontró el mismo genotipo infectando tanto a perros como a humanos que convivían en una misma casa o área común (31-33). Así mismo, en un estudio reciente realizado en Alemania se encontró que de 60 perros pertenecientes al área urbana que resultaron positivos, el 60% estaba infectado con el genotipo A, 12% con el genotipo C o D (específicos del perro) y en el 28% restante se presentó una infección mixta (genotipos específicos de perro y zoonóticos) (34). Igualmente, en un estudio realizado en perros de ambientes urbanos en Australia se encontró que estaban infectados principalmente por los genotipos A y D (35).
Estudios como los anteriormente mencionados que aplican la epidemiología molecular permiten llegar a conocer la heterogeneidad genética de la G. intestinalis (genotipos), y a partir de esta información poder estudiar la dinámica de transmisión de los diferentes genotipos y su asociación con focos o fuentes de infección, vías de transmisión, sintomatología y respuesta a tratamientos, aunando evidencias para demostrar el posible carácter zoonótico de la giardiasis.
El objetivo de esta revisión fue determinar la distribución geográfica en Norte, Centro y Sur América de los genotipos de G. intestinalis, identificando huésped, marcador genético y técnica molecular utilizados en la caracterización genotípica de aislados de humanos y caninos, por medio de una revisión sistemática de la literatura científica disponible de los últimos 19 anos.
materiales y métodos
Tipo de estudio
Se disenó una revisión sistemática de la evidencia científica disponible, desde 1990 hasta 2009, relacionada con la distribución geográfica en América de los genotipos A, B, C y D de Giardia intestinalis, hallados en humanos y en perros.
Bases de datos y búsqueda bibliográfica
Para realizar esta revisión se consultó la literatura científica disponible en las bases de datos PubMed (Medline), Science-Direct, SpringerLink y Scielo. También se utilizaron los motores de búsqueda como Scholar Google, con opciones avanzadas; las palabras claves utilizadas en la búsqueda fueron: Giardia, Giardia PCR, Giardia genotypes, Giardia assamblages. Los artículos de interés inicialmente fueron seleccionados por título y contenido del resumen, teniendo en cuenta los criterios de inclusión establecidos: artículos originales; en idioma espanol, inglés o portugués; realizados en países de América; investigaciones realizadas con aislamientos de G. intestinalis procedentes de humanos y perros publicados entre 1990 y 2009. Posteriormente, cada artículo fue revisado completamente y solo se escogieron aquellos que contenían información en la que se describirá la presencia de los genotipos A, B, C y D de Giardia intestinalis en humanos y caninos.
Las publicaciones seleccionadas se recopilaron impresas en carpetas, así como en archivos magnéticos formato PDF (Adobe Reader 7.0). Se construyó una base de datos (Excel) en la cual se clasificaron los artículos de acuerdo con el ano, país, autor, genotipo encontrado, tipo de huésped, secuencia y técnica usada para genotipificar.
resultados
Con la búsqueda bibliográfica inicial se logró obtener un total de 5319 artículos en PubMed, 2001 en ScienceDirect, 1855 en SpringerLink y 2 en Scielo (revista Biomédica). Después de la primera selección (título y resumen) y de eliminar los duplicados quedaron 30 artículos. Sin embargo, la lectura completa de estos permitió descartar 11 referencias más, por no contar con la información de interés. Finalmente, solo 19 artículos fueron escogidos para realizar la revisión, ya que cumplieron con todos los criterios de selección establecidos.
Teniendo en cuenta el objetivo propuesto en esta investigación, los artículos que llegaron a discriminar subgenotipo (AI, AII, AIII, AIV) en el proceso de clasificación del aislado fueron clasificados de acuerdo con el genotipo mayor al que pertenecen (genotipo A) (24,36).
Objetivos de los estudios
Como resultado de la revisión se encontraron un total de 19 artículos publicados con la información de interés durante la última década, cuyo producto del trabajo de investigación reportó la presencia de algún genotipo de Giardia intestinalis en huéspedes humanos o caninos en países de Norte, Centro y Sur América. En el primer lustro (2000 a 2005) se encontraron siete artículos: el primero fue realizado en 2000 en Canadá; en éste++ el objetivo fue la biotipificación de muestras clínicas recolectadas durante un brote, en humanos, animales y fuentes de agua (37); en 2002 México aporta un estudio, en el cual el objetivo fue genotipificar algunos aislados del parásito procedentes de humanos, animales y cultivos axénicos del parásito (38); en 2003, fueron publicados dos trabajos más: uno hecho en México, donde describieron el análisis genético de un panel de 26 aislados axénicos de G. intestinalis adquiridos independientemente (39), y otro en Estados Unidos, que tuvo como objetivo el desarrollo de un protocolo de anidada para amplificar el gen triosafosfato isomerasa (tpi) de G. intestinalis y de G. microti, así como la caracterización de la secuencia nucleotidica del fragmento amplificado (40). En 2004 se publica un trabajo realizado en Canadá, que evaluó el uso de PCR en tiempo real utilizando sondas dirigidas contra el gen |-giardin para la detección de Giardia en muestras humanas y diferenciación, en un solo paso, de los genotipos mayores A y B (41); 2005 se dieron a conocer dos nuevos trabajos, ambos realizados en México: en uno se genotipificaron aislados del parásito obtenidos de ninos y de perros y se estudió la relación con el huésped (6); el otro genotipificó G. intestinalis de humanos y perros a través de una PCR anidada que amplifica el gen |3-giardin (42).
En el segundo periodo (2006 a 2009) se encontraron doce artículos; en 2007 solo se encontraron estudios hechos con aislados obtenidos de países suramericanos (Brasil, Perú, Argentina y Colombia); cinco en total. Los artículos de Brasil (dos) se realizaron en el estado de Sao Paulo y en Río de Janeiro; en el de Sao Paulo se estudió aislados de G. intestinalis obtenidos de humanos, perros, gatos y ganado vacuno que habitaban ciudades del estado, por medio de una PCR (43), mientras que el estudio realizado en Río de Janeiro caracterizó genéticamente (locus |-giardin) aislados del parásito adquiridos de humanos y animales domésticos (mascotas) con el fin de investigar la prevalencia, datos de ocurrencia, epidemio-logía y potencial de trasmisión zoonótica de los genotipos (44). El objetivo del trabajo argentino fue adaptar un protocolo de PCR para la amplificación del gen tpi de Giardia lamblia en materia fecal formolada (45); en el artículo de Colombia el objetivo fue determinar el nivel de polimorfismo y la complejidad de las cepas circulantes en áreas específicas del país (46). La investigación hecha en Estados Unidos con aislados de G. intestinalis procedentes de humanos peruanos buscó demostrar la recombinación genética del parásito por medio de la evidencia proveniente de poblaciones (47).
El ano 2008 es en el que se encuentra el mayor número de publicaciones con los resultados de interés (Cuba, México, Nicaragua, Perú, Argentina y Brasil); siete en total. En Cuba se estudió la infección por Giardia en ninos del país, incluidos los resultados de epidemiología molecular utilizando PCR y secuenciación de dos marcadores moleculares (48); uno de los dos estudios mexicanos analizó muestras de Giardia de humanos y perros con el fin de comparar los respectivos genotipos por restricción del gen |-giardin (49), y el otro estudio usó una PCR múltiple para clasificar aislados de G. intestinalis obtenidos de cultivos axénicos y materia fecal de humanos y perros (50); en Nicaragua se realizó una PCR-RFLP y secuenciación de aislados de Giardia recuperados de humanos y perros que vivían en una misma área de León (51); el estudio realizado en Perú buscaba hallar la prevalencia de parásitos intestinales en ninos desnutridos de tres distritos de Trujillo, utilizando PCR-RFLP de G intestinalis (52); el estudio argentino buscó determinar los genotipos del parásito aislados de heces humanas y de perros que habitaban La Plata y General Mansilla, utilizando la técnica molecular PCR-RFLP (53); y el de Brasil, relacionar el genotipo detectado por PCR multiple en ninos del noroeste del país, con la presentación clínica, así como con la inflamación intestinal (54).
Estudios en Aislados de Humanos
Los países donde más se ha publicado sobre genotipificación de Giardia intestinalis provenientes de aislados de seres humanos es México, donde se realizaron el 31,5% (6/19) de los estudios (6,38,39,42,49,50); seguido de Brasil, que aportó el 15,8% (3/19) (43,44,54). El número de muestras genotipificadas en todos los estudios varió con un rango de 117, una media muestral de 28 aislados, una mediana de 20 y una desviación estándar de 27 (Figura 1).
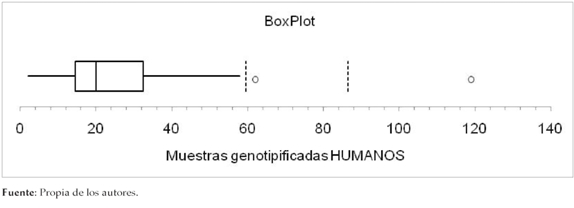
Figura 1. Muestras genotipificadas en estudios humanos
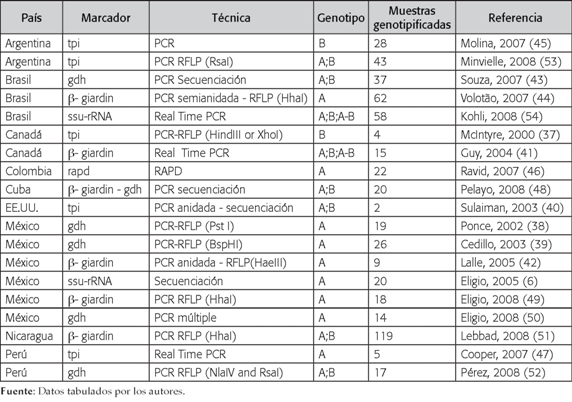
Los marcadores más utilizados para la ge-notipificación de los aislados de Giardia intestinalis fueron: el gen de |- giardin y el gen de triosafosfato isomerasa. Cada uno de estos se usó en el 26,3% (5/19) de los trabajos (37,40-42,44,45,47,49,51,53); con menor frecuencia se han utilizado el gen de la glutamato deshidrogenasa, la subunidad pequena del RNA ribosomal, el gen que codifica para la proteína de superfície variante-específica (6,38,39,43,47,54).
Las técnicas de biología molecular utilizadas con más frecuencia para genotipificar Giardia intestinalis en aislados de seres humanos fueron la PCR-RFLP, utilizada en el 47,4% (9/19), y la PCR y secuenciación en el 21% (4/19) de los estudios evaluados (6,37-40,42-44,47-49,51,53); también fueron utilizadas para la genotipificacion de Giardia intestinalis: PCR en tiempo real, PCR múltiple y Amplificación al Azar del DNA Polimórfico (RAPD) (41,46,47,50,54). El análisis del marcador |- giardin por medio de PCR-RFLP fue utilizado en el 21% (4/19) de los estudios (42,44,49,51); sin embargo, esta técnica también se utilizó para analizar el gen de triosafosfato isomerasa, el gen de glutamato deshidrogenasa y el gen de la proteína de superfície variante-específica (37-39,52,53).
La frecuencia de los genotipos de Giardia intestinalis evidenció que el genotipo A fue hallado en el 47,4% (9/19) de las investigaciones (6,38,39,42,49,50); en cambio, el genotipo B solo se encontró en el 10.5% (2/19) (37,45).
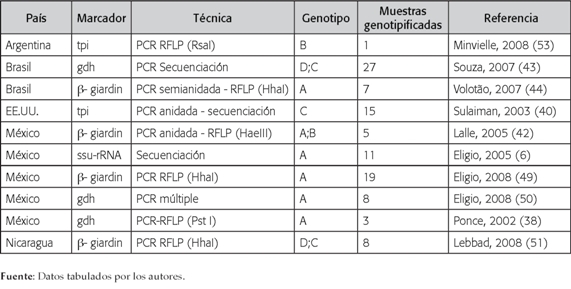
La distribución geográfica por países muestra que México y Colombia fueron los únicos países donde no se encontró el genotipo B infectando humanos, todos los aislados correspondieron al genotipo A (6,38,39,42,46,49,50), mientras que en Estados Unidos solo se detectó el genotipo B (40). En países como Canadá, Nicaragua, Cuba, Perú, Argentina y Brasil se encontraron ambos genotipos (A o B) entre los aislados analizados (41,44,48,51-53) (Figura 2) (Tabla 1).
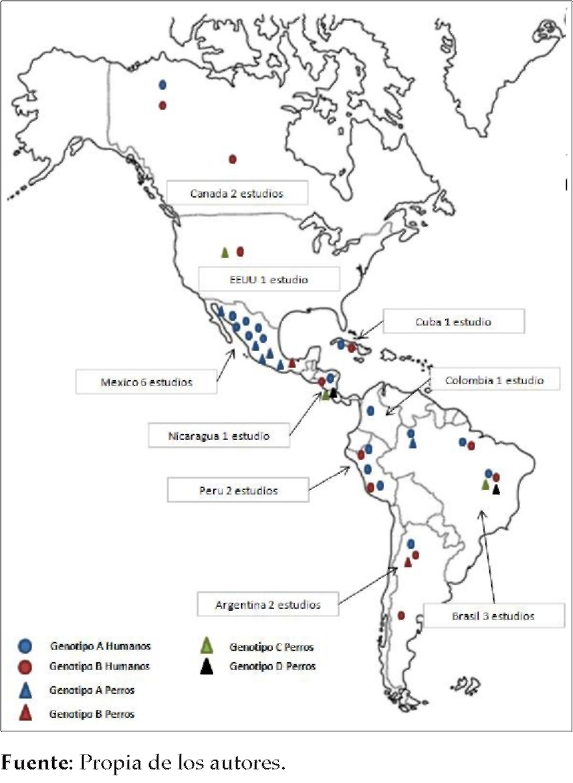
Figura 2. Mapa de distribución de los estudios sobre los cuatro genotipos de G. intestialis en humanos y caninos
Tabla 1
Estudios de genotipificación de Giardia intestinalis recuperada de humanos
Estudios en Aislados de Caninos
Los países donde más estudios de genopificación de Giardia intestinalis en aislados de caninos se han reportado son México,50% (5/10) (6,38,42,49,50), seguido de Brasil con un 20% (2/10) (43,44). El número de muestras genotipificadas en todos los estudios varió desde una muestra hasta 27, presentan una media muestral de 10 aislados, una mediana de 8 y una desviación están-dar de 8 (Figura 3).
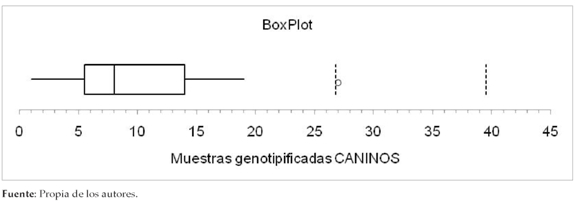
Figura 3. Muestras genotipificadas en estudios caninos
Los marcadores más utilizados para genotipificar fueron |3-giardin, utilizado en el 40% (4/10) de los estudios (42,44,49,51), seguido de los genes que codifican para triosafosfato isomerasa, y glutamato deshidrogenasa, cada uno con un 20% (2/10) (40,43,50,53).
Las técnicas de biología molecular reportadas con mayor frecuencia en los estudios fueron PCR-RFLP, con un 60% (6/10), y PCR-secuenciación, con un 30% (3/10) (6,38,40,4244,49,51,53) El marcador |3-giardin se utilizó en el 40% (4/10) de las publicaciones para genotipificar los aislados, y en todos los casos su análisis se hizo por medio de la técnica PCR-RFLP (42,44,49,51). El genotipo A fue el más encontrado en los aislados de caninos, con un 60% (6/10) (6,38,42,44,49,50); solo en un 10% (1/10) de los estudios se observó perros infectados solamente con el genotipo B (53), mientras que en un 30% (3/10) se reportaron la presencia en aislados de los genotipos huésped-específicos (C o D) (40,43,51). El genotipo A en caninos fue reportado en México y en Brasil (6,38,42,44,49,50), el genotipo B en caninos de Argentina, así como de México (42,53), y los genotipos huésped-específicos (C o D) se encontraron en perros de Estados Unidos, Nicaragua y Brasil (40,43,51) (Gráfica 2) (Tabla 2).
Tabla 2
Estudios de genotipificación aislados recuperados de caninos
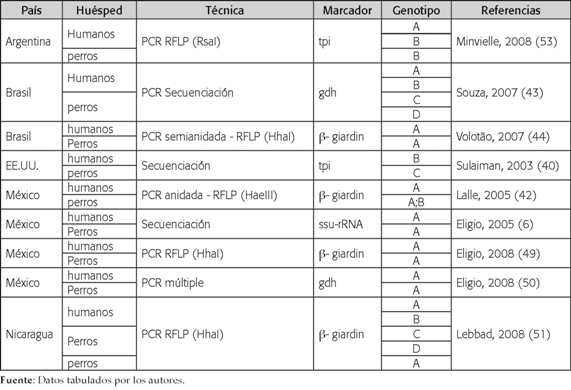
De los 19 artículos seleccionados, solo el 52,6% (10/19) genotipificó G. intestinalis recuperada tanto de humanos como de perros en el mismo estudio (6,38,40,42-44, 49-51,53). Las frecuencias encontradas con respecto a países donde se realizaron los trabajos, marcadores usados y técnicas empleadas son iguales a las informadas anteriormente en el análisis que se hizo a los estudios que involucraron aislados de caninos. Sin embargo, la proporción de los genotipos fue diferente, dado que el 50% (5/10) de las investigaciones demostró la presencia solo del genotipo A entre los humanos y perros muestreados (6,38,44,49,50), en un 20% (2/10) se hallaron los genotipos A y B infectando ambos huéspedes (42,53)(Tabla 3).
Tabla 3
Estudios de genotipificación de Giardia intestinalis recuperada de humanos y caninos
Conclusiones
La literatura científica reporta que los humanos sólo pueden ser infectados por los genotipos A o B (24,36), tal como se evidenció en esta revisión, pues todos los aislados recuperados de las personas correspondieron a estos genotipos, aunque el genotipo A fue el más frecuentemente detectado, lo cual contrasta con lo informado por Caccio et al. (2008), quienes proponen que el genotipo B es el mayor responsable de la infecciones en humanos (36).
En el 60% de los trabajos que genotipificaron aislados recuperados de ambos huéspedes se encontró que los caninos estaban infectados con los mismos genotipos detectados en los humanos (A-B); el hallazgo de estos genotipos, considerados zoonóticos en caninos y humanos, sugiere una posible transmisión zoonótica. Datos similares a los descritos en estos estudios han sido obtenidos en trabajos realizados en India (2004), Alemania (2007) y Tailandia (2009) (32-34); en estos casos, los caninos se podrían estar comportando como fuentes potenciales de infección para las personas, debido, hipotéticamente, a un proceso de desplazamiento de los genotipos específicos de especie, generado por los genotipos zoonóticos. En ese mismo sentido, se plantea la hipótesis según la cual en lugares geográficos donde se presente un fenómeno de alta transmisibilidad de genotipos específicos del parásito en seres humanos y baja transmisibilidad de genotipos específicos entre los caninos.
En comparación con los estudios realizados con otros microorganismos, las técnicas de genotipificación de G. intestinalis no son particularmente avanzadas, y gran parte de estas técnicas se han basado en el análisis de una variedad importante de marcadores. Quizás esta es una de las razones por las que todavía la transmisión zoonótica del parásito continua siendo tema de controversia, pues no todos los resultados obtenidos con estas técnicas son comparables. Este trabajo evidencia cómo, a través de los tiempos, se han venido empleado una cantidad de técnicas y marcadores para poder caracterizar aislados procedentes de humanos y de perros. Un total de cinco marcadores y de seis técnicas moleculares fueron utilizadas en los 19 estudios revisados; incluso, en alguno se utilizó más de un marcador (39,48).
Finalmente, se pudo observar que los genotipos A y B están ampliamente distribuidos entre personas de países de Norte, Centro y Sur América. El genotipo A en los perros, al igual que en los humanos, fue el más encontrado, lo que indica que estos animales pueden estar desempenando un papel importante en la transmisión zoonótica directa y posiblemente endémica de G. intestinalis. Sin embargo, es necesario proponer investigaciones que permitan conocer la compleja dinámica de transmisión del parásito entre ambos huéspedes.
Financiación: Los recursos para la realización del proyecto fueron aportados por el Grupo de Investigación del Instituto Colombiano de Medicina Tropical, Universidad CES y el Grupo de Investigación en Microbiología Veterinaria de la Universidad de Antioquia.
Conflicto de intereses: Los autores manifiestan que no se presentaron conflictos de intereses para la realización de este estudio en ninguna de sus fases.
REFERENCIAS
(1) Thompson R. The zoonotic significance and molecular epidemiology of Giardia and giardiasis. Vet Parasito! 2004; 126 (1-2): 15-35. [ Links ]
(2) Lane S, Lloyd D. Current trends in research into the waterborne parasite Giardia. Crit Rev Microbiol 2002; 28 (2): 123-47. [ Links ]
(3) Rivera M, La parte M, Hurtado P, Magaldi L, Collazo M. Giardiasis intestinal. Minirevision. Invest Clin 2002; 43: 119-28. [ Links ]
(4) Thompson R. Giardiasis as a re-emerging infectious disease and its zoonotic potential. Int J Parasito! 2000; 30 (12-13): 1259-67. [ Links ]
(5) WHO. Fighting Disease Fostering Development. Geneva: World Health Organizations;1996. [ Links ]
(6) Eligio-García L, Cortés-Campos A, Jiménez-Cardoso E. Genotype of Giardia e intestinalis isolates from children and dogs and its relationship to host origin. Parasitol Res 2005 Aug; 97 (1): 1-. [ Links ]
(7) Corredor A AE, Hernández CA. Parasitismo intestinal. Bogotá, D.C.: Instituto Nacional de Salud; 2000. [ Links ]
(8) Traub R, Robertson I, Irwin P, Mencke N, Thompson R. Canine gastrointestinal parasitic zoonoses in India. Trends in parasitology 2005; 21 (1): 42-8. [ Links ]
(9) Dobell C. The discovery of the intestinal protozoa of man. Proc R Soc Med (Sect Hist Med) 1920; 13: 1-15. [ Links ]
(10) Lambl W. Mikroskopische untersuchungen der Darmexcrete. Vierteljahrsschr Prakst Hei-kunde 1859; 61: 1-58. [ Links ]
(11) Nash T, McCutchan T, Keister D, Dame J, Conrad J, Gillin F. Restriction-endonuclea-se analysis of DNA from 15 Giardia isolates obtained from humans and animals. J Infect Dis 1985: 64-73. [ Links ]
(12) Nash T, Mowatt M. Identification and characterization of a Giardia lamblia group-specific gene. Exp Parasitol 1992; 75 (4): 36972. [ Links ]
(13) Keulen H, Homan WL, Erlandsend SL, Jarroll EL. A three nucleotide signature sequence in small subunit rRNA divides human Giardia in two different genotypes. J Eukaryot Microbiol 1995; 42 (4): 392-4. [ Links ]
(14) Homan W, Enckevort F, Limper L, Eys G, Schoone G, Kasprzak W et al. Comparison of Giardia isolates from different laboratories by isoenzyme analysis and recombinant DNA probes. Parasitol Res 1992; 78 (4): 316-23. [ Links ]
(15) Ey PL, Mansouri M, Kulda J, Nohynkova E, Monis PT, Andrews RH et al. Genetic analysis of Giardia from hoofed farm animals reveals artiodactyl-specific and potentially zoonotic genotypes. J Eukaryot Microbiol 1997; 44 (6): 626-35. [ Links ]
(16) WHO. Parasitic zoonoses. Geneva: World Health Organisation;1979. Contract n° 637. [ Links ]
(17) Meloni B, Thompson R, Strandén A, Kohler P, Eckert J. Critical comparison of Giardia duodenalis from Australia and Switzerland using isoenzyme electrophoresis. Acta Trop 1991 Dec; 50 (2): 115-24. [ Links ]
(18) Lu S, Li J, Zhang Y, Wen J, Wang F. The intraspecific difference of the triose phosphate isomerase (tim) gene from Giardia lamblia. Chin Med J (Engl) 2002 May; 115 (5): 763-6. [ Links ]
(19) Sarafis K, Isaac-Renton J. Pulsed-field gel electrophoresis as a method of biotyping of Giardia duodenalis. Am J Trop Med Hyg 1993 Jan; 48 (1): 134-44. [ Links ]
(20) Wielinga C, Thompson R. Comparative evaluation of Giardia duodenalis sequence data. Parasitology 2007 Nov; 134 (Pt 12): 1795-821. [ Links ]
(21) G, Andrews R, Ey P, Chilton N. Division of Giardia isolates from humans into two genetically distinct assemblages by electrophoretic analysis of enzymes encoded at 27 loci and comparison with Giardia muris.Parasitology 1995; 111 (1): 11-7. [ Links ]
(22) Hopkins R, Meloni B, Groth D, Wetherall J, Reynoldson J, Thompson R. Ribosomal RNA sequencing reveals differences between the genotypes of Giardia isolates recovered from humans and dogs living in the same locality. J Parasitol 1997: 44-51. [ Links ]
(23) Andrews RH, Adams M, Boreham PF, Myrhofer G, Meloni BP. Giardia intestinalis: electrophoretic evidence for species complex. Int J Parasitol 1989;19:183-91. [ Links ]
(24) Adam R. Biology of Giardia lamblia. Clin Microbiol Rev 2001; 14 (3): 447-75. [ Links ]
(25) Monis P, Andrews R, Mayrhofer G, Ey P. Genetic diversity within the morphological species Giardia intestinalis and its relationship to host origin. Infect Genet Evol 2003; 3(1): 29-38. [ Links ]
(26) Read C, Walters J, Robertson I, Thompson R. Correlation between genotype of Giardia duodenalis and diarrhoea. Int J Parasitol 2002; 32 (2): 229-31. [ Links ]
(27) Slifko T, Smith H, Rose J. Emerging parasite zoonoses associated with water and food. Int J Parasitol 2000; 30 (12-13): 1379-93. [ Links ]
(28) Ponce-Macotela M, Peralta-Abarca G, Mar-tínez-Gordillo M. Giardia intestinalis and other zoonotic parasites: prevalence in adult dogs from the southern part of Mexico City. Vet Parasitol 2005; 131 (1-2): 1-4. [ Links ]
(29) Pacheco R. Mascotas en los hogares: enfermedades de los ninos adquiridas por convivenda con animales. Enferm Infecc Microbiol Clin 2003; 23 (4): 137-48. [ Links ]
(30) Botero D. Parasitosis humanas. Medellín: Corporación para Investigaciones Biológicas; 2006. [ Links ]
(31) Traub R, Robertson I, Irwin P, Mencke N, Thompson R. The role of dogs in transmission of gastrointestinal parasites in a remote tea-growing community in northeastern India. Am J Trop Med Hyg 2002; 67 (5): 539-45. [ Links ]
(32) Traub R, Monis P, Robertson I, Irwin P, Mencke N, Thompson R. Epidemiological and molecular evidence supports the zoonotic transmission of Giardia among humans and dogs living in the same community. Parasitology 2004 Mar; 128 (Pt 3): 253-62. [ Links ]
(33) Traub R, Inpankaew T, Reid S, Sutthikorn-chai C, Sukthana Y, Robertson I et al. Transmission cycles of Giardia duodenalis in dogs and humans in Temple communities in Bangkok -a critical evaluation of its prevalence using three diagnostic tests in the field in the absence of a gold standard. Acta Trop 2009 Aug; 111 (2): 125-32. [ Links ]
(34) Leonhard S, Pfister K, Beelitz P, Wielinga C, Thompson R. The molecular characterisation of Giardia from dogs in southern Germany. Vet Parasitol 2007 Nov; 150 (1-2): 33-8. [ Links ]
(35) Thompson RCA, Morgan UM, Mellor KJ, Hopkins RM. Genotyping Giardia and Cryptosporidium. Today"s Life Sci 1999; 11: 80-5. [ Links ]
(36) Cacciò S, Ryan U. Molecular epidemiology of giardiasis. Mol Biochem Parasitol 2008; 160 (2): 75-80. [ Links ]
(37) McIntyre L, Hoang L, Ong C, Lee P, Isaac-Renton J. Evaluation of molecular techniques to biotype Giardia duodenalis collected during an outbreak. J Parasitol 2000 Feb; 86 (1): 172-7. [ Links ]
(38) Ponce-Macotela M, Martínez-Gordillo M, Bermúdez-Cruz R, Salazar-Schettino P, Or-tega-Pierres G, Ey P. Unusual prevalence of the Giardia intestinalis A-II subtype amongst isolates from humans and domestic animals in Mexico. Int J Parasitol 2002 Aug; 32 (9): 1201-2. [ Links ]
(39) Cedillo-Rivera R, Darby J, Enciso-Moreno J, Ortega-Pierres G, Ey P. Genetic homogeneity of axenic isolates of Giardia intestinalis derived from acute and chronically infected individuals in Mexico. Parasitol Res 2003 Jun; 90 (2): 119-23. [ Links ]
(40) Sulaiman I, Fayer R, Bern C, Gilman R, Trout J, Schantz P et al. Triosephosphate isomerase gene characterization and potential zoonotic transmission of Giardia duodenalis. Emerg Infect Dis 2003 Nov; 9 (11):1444-52. [ Links ]
(41) Guy R, Xiao C, Horgen P. Real-time PCR assay for detection and genotype differentiation of Giardia lamblia in stool specimens. J Clin Microbiol 2004 Jul; 42 (7): 3317-20. [ Links ]
(42) Lalle M, Jimenez-Cardosa E, Cacciò S, Pozio E. Genotyping of Giardia duodenalis from humans and dogs from Mexico using a beta-giardin nested polymerase chain reaction assay. J Parasitol 2005 Feb; 91 (1): 203-5. [ Links ]
(43) Souza S, Gennari S, Richtzenhain L, Pena H, Funada M, Cortez A et al. Molecular identification of Giardia duodenalis isolates from humans, dogs, cats and cattle from the state of São Paulo, Brazil, by sequence analysis of fragments of glutamate dehydrogenase (gdh) coding gene. Vet Parasitol 2007 Nov; 149 (3-4): 258-64. [ Links ]
(44) Volotão A, Costa-Macedo L, Haddad F, Brandão A, Peralta J, Fernandes O. Geno-typing of Giardia duodenalis from human and animal samples from Brazil using beta-giardin gene: a phylogenetic analysis. Acta Trop 2007 Apr; 102 (1): 10-9. [ Links ]
(45) Molina N, Polverino D, Minvielle M, Basualdo J. PCR amplification of triosephos-phate isomerase gene of Giardia lamblia in formalin-fixed feces. Rev Latinoam Microbiol 2007; 49 (1-2): 6-11. [ Links ]
(46) Ravid Z, Duque S, Arévalo A, Nicholls R, Wasserman M. Genetic diversity of Giardia intestinalis populations in Colombia. Biomédica 2007;27(1):34-41. [ Links ]
(47) Cooper M, Adam R, Worobey M, Sterling C. Population genetics provides evidence for recombination in Giardia. Curr Biol 2007 Nov; 17 (22): 1984-8. [ Links ]
(48) Pelayo L, Núnez F, Rojas L, Furuseth Hansen E, Gjerde B, Wilke H et al. Giardia infections in Cuban children: the genotypes circulating in a rural population. Ann Trop Med Parasitol 2008 Oct; 102 (7): 585-95. [ Links ]
(49) Eligio-García L, Cortes-Campos A, Cota-Guajardo S, Gaxiola S, Jiménez-Cardoso E. Frequency of Giardia intestinalis assemblages isolated from dogs and humans in a community from Culiacan, Sinaloa, Mexico using beta-giardin restriction gene. Vet Parasitol 2008 Oct; 156 (3-4): 205-9. [ Links ]
(50) Eligio-García L, Cortés-Campos A, Jimé-nez-Cardoso E. Classification of Giardia intestinalis isolates by multiple polymerase chain reaction (multiplex). Parasitol Res 2008 Sep; 103 (4): 797-800. [ Links ]
(51) Lebbad M, Ankarklev J, Tellez A, Leiva B, Andersson J, Svàrd S. Dominance of Giardia assemblage B in Leon, Nicaragua. Acta Trop 2008; 106 (1): 44-53. [ Links ]
(52) Peréz Cordón G, Cordova Paz Soldan O, Vargas Vasquez F, Velasco Soto J, Sempere Bordes L, Sánchez Moreno M et al. Prevalence of enteroparasites and genotyping of Giardia lamblia in Peruvian children. Parasitol Res 2008 Jul; 103 (2): 459-65. [ Links ]
(53) Minvielle M, Molina N, Polverino D, Basualdo J. First genotyping of Giardia lamblia from human and animal feces in Argentina, South America. Mem Inst Oswaldo Cruz 2008 Feb; 103 (1): 98-103. [ Links ]
(54) Kohli A, Bushen O, Pinkerton R, Houpt E,Newman R, Sears C et al. Giardia duodenalis assemblage, clinical presentation and markers of intestinal inflammation in Brazilian children.Trans R Soc Trop Med Hyg 2008 Jul; 102 (7): 718-25. [ Links ]

