










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Salud Uninorte
Print version ISSN 0120-5552
Salud, Barranquilla vol.28 no.1 Barranquilla Jan./June 2012
ARTÍCULO DE REVISIÓN / REVIEW ARTICLE
Procesos embrionarios y malformaciones congénitas. Revisión con reporte de casos
Embryonic processes and congenital malformations. Review with case reports
Luis Cervantes Parra1, Alfonso Londoño Orozco2, Marco Nieto García3, Ricardo Gutiérrez De Aguas4
1 Docente investigador, Universidad del Quindío. Armenia (Colombia).
2 Docente, Facultad Ciencias de la Salud, Universidad del Quindío. Armenia (Colombia).
3 Docente, Facultad Ciencias de la Salud, Universidad del Quindío. Armenia (Colombia).
4 Profesor asociado, Departamento de Química y Biología, Universidad del Norte. Barranquilla (Colombia).
Correspondencia: Luis Cervantes Parra. Universidad del Norte, Km 5, vía a Puerto Colombia. Barranquilla (Colombia). Fax 5-3598852. luiscerva7@gmail.com
Fecha de recepción: 20 de junio de 2011
Fecha de aceptación: 12 de enero de 2012
Resumen
Los procesos fundamentales de la dinámica embrionaria en el reino animal, y particularmente en los vertebrados, obedecen a un programa genético que solo ha comenzado a comprenderse en los últimos años. Uno de los principales objetivos de los biólogos del desarrollo es develar el misterio de cómo un oocito, después de ser fecundado, se transforma en un organismo multicelular. Esta dinámica requiere la activación de un complejo programa de desarrollo en el que genes específicos se expresan en una secuencia temporal precisa, y en la ubicación correcta, para dar origen a diferentes tipos de tejidos como la piel, músculos y nervios, entre otros. Mediante el estudio de mutantes en Drosophila se han identificado genes que participan en la organización del patrón de desarrollo del embrión, los cuales son activos en las hembras. Asimismo, en los últimos años, gracias a las técnicas moleculares, se han realizado grandes avances en el conocimiento de los mecanismos que controlan este intrincado proceso. Este trabajo plantea diversos aspectos relacionados con la teratogenia asociada a la gastrulación, periodo que es muy sensible a las agresiones, así como el aporte de casos y un análisis de los avances en el conocimiento de los procesos moleculares implicados en la dinámica embrionaria.
Palabras clave: Desarrollo embrionario, malformaciones congénitas, dinámica embrionaria, procesos moleculares.
Abstract
The fundamental processes of embryonic dynamics in the animal kingdom and particularly in vertebrates are due to a genetic program that has begun to be understood in recent years. One of the developmental biologists' main goals is to unravel the mystery of how after being fertilized an oocyte becomes a multicellular organism. This dynamic requires activation of a complex development program in which genes are expressed in a temporal, precise sequence and in the correct location to give rise to different types of tissues such as skin, muscles and nerves among others. The genes involved in the embryo development pattern, active in females, have been identified by studying Drosophila mutants. Also, thanks to molecular techniques, advances in understanding the intricate mechanism that control this process have been significant in recent years. This work raises several issues related to teratogenicity associated with gastrulation, embryonic stage sensitive to damages. Also, this work provides study cases and a review of the progress in the understanding of molecular processes involved in embryonic dynamics.
Keywords: Embryonic development, congenital malformations, embryonic dynamic, molecular process.
INTRODUCCIÓN
Los procesos embrionarios en la escala animal, y particularmente en la de los cordados vertebrados, obedecen a un plan genético que solo ha comenzado a comprenderse en los últimos años. Esta comprensión del desarrollo ha progresado sobre todo en un invertebrado como es la mosca de la fruta, Drosophila melanogaster, en la que se ha estudiado el mecanismo de determinación de los dos ejes principales del embrión: antero- posterior y dorso-ventral, el mecanismo de asignación del tipo de estructura segmentaria y se ha llegado a identificar una serie de genes con homeobox y el plan corporal de los vertebrados (1).
En épocas recientes, los embriólogos han realizado grandes avances en el conocimiento de los mecanismos moleculares que controlan este proceso, otrora misterioso. Gracias a las poderosas técnicas de la biología molecular que han permitido aislar y caracterizar genes particulares que intervienen en algunas de las decisiones relativas al establecimiento del esquema corporal embrionario; cuya clave reside en una familia de genes conocidos como genes con homeobox (caja homeótica), que subdividen al embrión temprano en campos celulares dotados de la capacidad de transformar en tejidos y órganos específicos. El proceso de desarrollo de Xenopus laevis (una rana sudafricana) (1); Bufo arenarum en Argentina (2, 3, 4) y Bufo marinus en Colombia (5, 6) constituyen ejemplos de cómo toma forma un vertebrado durante su formación embrionaria y posterior desarrollo. Sin embargo, y a pesar de la serie de adelantos moleculares y sus resultados en Drosophila (7), estos no son fácilmente extrapolables a los cordados, dadas las diferencias en el proceso de desarrollo entre un protostómado como la mosca y un deuterostómado, incluido en este grupo al humano. Pero a pesar de las grandes diferencias entre la mosca y los vertebrados, en este último grupo se encuentran también los mismos genes homeóticos de la mosca, con una disposición en familia génica; ejemplos de estos genes son HOX y PAX (7). De igual manera, los genes HOM-C y el gen de segmentación "apareado" se ha constituido en tema central de la embriogénesis, y a partir de él se han dado los primeros pasos para entender las perturbaciones tempranas del desarrollo humano, por posibles alteraciones de genes reguladores, algunos de los cuales funcionan o se expresan de manera normal durante la formación embrionaria, como los que se describen más adelante en este trabajo.
Uno de los conceptos más sugestivos e importantes que han servido y sirven de base en la investigación embriológica actual es el concepto de "inducción embrionaria". Concepto que ha surgido como corolario de una extensa serie de investigaciones que sintetizan el resultado de acciones bioquímicas que un esbozo embrionario ejerce sobre otro esbozo todavía no determinado y al que orienta hacia una muy precisa diferenciación. En este proceso, al primer esbozo se considera como "inductor" y al segundo como "territorio competente" (8). Por otra parte, conocer cómo funciona un "inductor" significa conocer el funcionamiento del embrión organizado, debido a que las posibilidades de que este organice a un embrión normal, o uno anormal, cuando está implantado representan el resultado de una suma de acciones inductoras sinérgicas y consecuenciales (8).
TERATOGENIA ASOCIADA CON LA GASTRULACIÓN
El comienzo de la tercera semana del desarrollo en humano es un periodo muy sensible a las agresiones teratógenas (9). Este periodo equivale al inicio de la gastrulación, momento en el que se puede trazar el "mapa" del destino final de los diferentes sistemas del organismo, como los ojos, esbozo encefálico, etc., cuyas poblaciones celulares pueden ser dañadas por sustancias teratógenas. Por ejemplo, durante este período cantidades excesivas de alcohol destruyen las células de la línea media, por ende, con compromiso o daño en las estructuras craneocefálicas de la línea media, y provoca holoprosencefalia (9).
La gastrulación por sí misma puede interrumpirse por anomalías genéticas o por agentes tóxicos (9). Este es el caso de la disgenesia caudal (sirenomelia), en la que no se forma suficiente mesodermo en la región más caudal del embrión y conlleva a la aparición de anomalías de estas estructuras (10 - 12). El mesodermo contribuye a la formación de las extremidades inferiores, el sistema urogenital (mesodermo intermedio) y las vertebras lumbosacras.
Otras de las patologías asociadas con la teratogenia y con la gastrulación es la transposición visceral, malformación caracterizada por la trasposición de las vísceras en el tórax y el abdomen (9). Además pueden aparecer tumores asociados con la gastrulación, como los teratomas sacrococcígeos, que suelen contener tejidos derivados de las tres capas embrionarias (ectodermo, endodermo y mesodermo). Estos tumores son los más comunes en los recién nacidos, y su frecuencia es de un caso por cada 37 000 nacimientos. Estos tumores también pueden originarse a partir de células germinales primigenias que no logran migrar hacia la cresta gonadal (9).
MALFORMACIONES CONGÉNITAS Y REPORTE DE CASOS
Teratoma de la línea media craneal
Los tumores de la cavidad oral y de la nasofaringe son poco comunes en el recién nacido. El epignato es un teratoma de la región orofaríngea compuesto de células del ectodermo, mesodermo y endodermo que generalmente se extiende a través de la boca como una masa evidente. Los teratomas son los tumores de células germinales extragonadales más comunes en la infancia, pues llegan a afectar a uno por cada 40 000 nacidos vivos. Estos tumores están bien circunscritos y contienen una diversidad de tejido maduro e inmaduro, entre los que se identifica: epitelio escamoso, respiratorio, glandular y gastrointestinal; tejido de la glándula tiroides, del páncreas e hígado; tejido nervioso, cartílago, hueso, músculo y grasa.
Los teratomas más frecuentes son los sacrococcígeos (45 a 65%), de las gónadas (10 a 35%), del mediastino anterior (10 a 12%), retroperitoneal (3 a5%), cervical (3 a 6%), presacral (3 a 5%), del sistema nervioso (2 a 4%) y menos del 2% son orofaríngeos. El epignato es un teratoma congénito del paladar duro, localizado en la región de la bolsa de Rathke. La mayor parte se desarrolla de manera unidireccional, dentro de la cavidad oral, y provoca obstrucción de la vía aérea, pero cuando el crecimiento es bidireccional, afecta las estructuras intracraneales.
La incidencia de epignato varía de uno por cada 35 000 a uno por cada 200 000 nacidos vivos, y suele afectar con mayor predominio a las mujeres, en una relación de tres a uno (13).
En Armenia (Quindío, Colombia) se reportó un caso excepcional, producto de una multigestante de 28 años remitida al servicio de gineco-obstetricia del Hospital Universitario San Juan de Dios (HUSD), que correspondió a un teratoma de la línea media craneal, diagnosticado prenatalmente mediante ultrasonido, junto con la confirmación y correlación anatomopatológica, y el análisis embriogenético (14).
En el estudio se practicó una ecografía de nivel III, que reveló presentación cefálica enclavada en el tercio inferior, por lo que fue difícil la identificación de la línea media. A nivel del cavum del septúm pelúcidum, en la fosa media izquierda, se observó una zona anecoica quística irregular bilobulada sugestiva de lesión vascular. Sobre el parietal izquierdo y la sutura coronaria izquierda se observó una línea de continuidad. La órbita izquierda se encontró invadida por una gran masa redondeada y lobulada, que medía 10 x 10 cm, la cual desplazaba el globo ocular y el pabellón auricular izquierdo; además ocupaba la región frontoparietal izquierda. El globo oscular izquierdo se observó aumentado y mostraba conformaciones anormales con lesión hiperecoica sugestiva de catarata (figura 1).
Por los hallazgos descritos se propusieron los siguientes diagnósticos: embarazo único vivo con edad gestacional de 40 semanas; polihidramnios severo a tensión; malformación facial por teratoma y malformación cerebrovascular. La revisión de la literatura confirma la rareza de este tipo de tumores (15-19).

Extrofia cloacal por bridas amnióticas
El síndrome de bridas amnióticas (SBA) abarca un amplio espectro de alteraciones congénitas que se caracteriza por anillos de constricción, pseudosindactilia, amputaciones, y menos frecuente, múltiples defectos craneofaciales, viscerales y alteraciones en la pared abdomino-torácica (20). Su incidencia varía entre 1 en 12 000 a 1 en 15 000 recién nacidos vivos, y en abortos espontáneos puede llegar a ser de 178 en 10 000. Afecta a ambos sexos por igual, con una incidencia ligeramente elevada en descendientes afrocaribeños. Su presentación tiene un carácter esporádico.
Herman y Opitz en 1974 (21) propusieron el término ADAM para referirse a esta entidad conocida también como síndrome de bridas amnióticas o constricciones anulares y amputaciones congénitas. Acrónimo que describe la combinación más importante de anomalías en este trastorno: Amniótica, Deformidades, Adhesiones y Mutilaciones. Por ser una secuencia disruptiva, hoy se le conoce como complejo de disrupción por bandas amnióticas. Si bien las extrofias cloacales son defectos poco comunes, se produce uno de cada 30 000 nacimientos, mientras que la frecuencia de epispadias es de uno en 120 000 y la extrofia cloacal, de uno en 400 000. En todas estas anomalías existe preponderancia del varón sobre la mujer.
En el departamento del Quindío se reportó un caso de extrofia cloacal por bridas amnióticas con compromiso de musculatura perineal y defectos congénitos del área perineal que derivan del desarrollo defectuoso de la membrana cloacal (22). El caso fue un feto de sexo femenino, obtenido de parto por cesárea en el HUSD de Armenia (Quindío), que midió 32 cm de la corona al talón, pesó 1075 g y presentaba múltiples malformaciones congénitas.
Al examen externo, la cabeza presentó una circunferencia de 34 cm, cabello de color negro de implantación normal, pabellones auriculares de implantación baja y diámetro torácico de 26 cm. A nivel abdominal presentaba bridas amnióticas en hemiabdomen derecho y defecto de cierre de la pared abdominal y perineal, con exposición de estómago y transposición visceral. Los genitales externos eran de aspecto normal. Se observaron múltiples malformaciones, cráneo de aspecto dolicocefálico alargado (escafocefalia), como resultado de una craneosinostosis severa, con imágenes de plata martillada. Alteraciones en MS y MI, pelvis, a nivel de columna lumbar, sacro, coxígea y abdominal (figura 2).

Amputaciones congénitas por bridas amnióticas
Entre 1986 y 1990, como parte del Programa de prevención de las malformaciones congénitas de las facultades de Medicina de la Universidad Tecnológica de Pereira y del Quindío, se estudiaron siete pacientes, de ambos sexos, diagnosticados con el Complejo de disrupción por bandas amnióticas o Complejo de ADAM, desde el periodo prenatal hasta la edad de seis años (23).
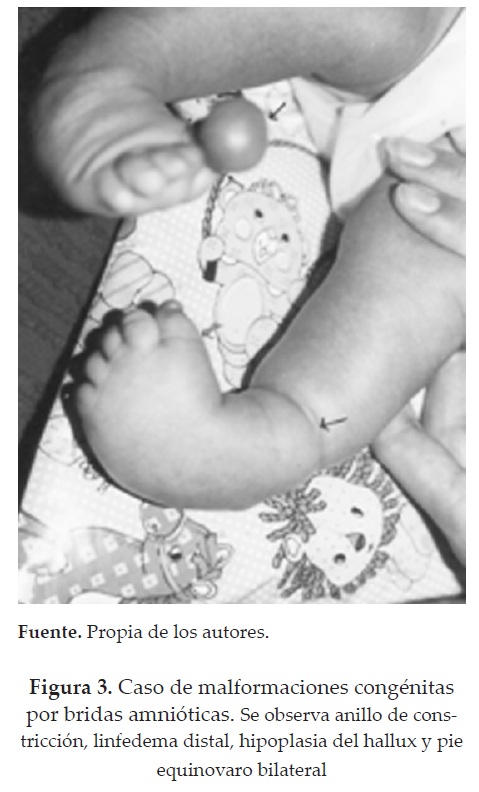
El estudio se realizó con base en la historia clínica, estudio radiológico y ecográfico. En los casos analizados estaban afectadas principalmente las extremidades (figura 3). La deformación predominante fue el pie bilateral. En uno de los casos se encontró ingesta de medicamentos antieméticos, considerados sin efecto teratogénico, como el clorofenotiazonil-6-copina.
Si bien el Complejo de disrupción por bandas amnióticas es una patología frecuente y una de las causas más importantes de amputaciones congénitas e hipoplasia de las extremidades, su diagnóstico temprano orienta hacia un manejo adecuado.
La revisión de la literatura muestra que los teratomas más frecuentes son los de la región sacrococcígea y en menor proporción los de la región craneofacial. El teratoma orofaríngeo, con una incidencia de 1:35.000 a 1:200.000 nacidos vivos, surge de la base del cráneo o de la parte posterior de la faringe y se extiende interiormente, causando obstrucción parcial o completa del tracto aéreo y digestivo (13), de manera similar a lo ocurrido en el caso reportado. En La Paz (Bolivia) se reportó también un caso relacionado con un teratoma maduro craneofacial derecho (19). El epignato, o teratoma orofaríngeo, es un teratoma congénito del paladar duro, localizado en la región de la bolsa de Rathke; la mayor parte se desarrolla de manera unidireccional, dentro de la cavidad oral, y provoca obstrucción de la vía aérea, pero cuando el crecimiento es bidireccional afecta las estructuras intracraneales (24). Con relación a este tipo de malformación se ha reportado un caso clínico de un nacido a través de cesárea, con 35 semanas, Apgar 2-4, con hipotonía, y con una masa sólida de 5 cm, quien falleció a las tres semanas (25). Los epignatos gigantes que ocupan toda la orofaringe pueden ocasionar acumulación de líquido amniótico en el feto y secundariamente polihidramnios (26).
Con relación al síndrome de bridas amnióticas, este es un proceso que se inicia 28 días después de la concepción y durante las 12 primeras semanas del embarazo (etapa de embriogénesis). Es conocido también como secuencia de bandas amnióticas, complejo disrupción amniótico, amputación congénita, bandas congénitas de constricción, entre otras, e incluye un amplio espectro de deformaciones, con una incidencia que se estima en 7.7 por 10 000 nacidos vivos y se considera implicada en 178 por 10 000 abortos espontáneos (20). En este trabajo se presenta un reporte de casos en el que se describen diferentes manifestaciones clínicas de esta entidad y se hace una revisión de diversas hipótesis etiológicas.
En Brasil se identificaron 270 casos con algunas evidencias sobre pie de constricción y que presentaban, además, defectos sugestivos de la secuencia de ADAM (27). El estudio se realizó entre 1982 y 1998, con hallazgos en los pacientes estudiados de acrania, encefalocele, hendiduras faciales, coloboma del parpado, entre otros. Tres casos clínicos de pacientes, recién nacidos masculinos, con síndrome de bridas amnióticas han sido reportados en Chile (28), así como se ha hecho una revisión de las principales características: bridas amnióticas, adherencias intrauterinas que condicionan la llamada "sabana amniótica", y tabiques uterinos secundarios a anomalías mullerianas (29). En Murcia (España), la revisión anual de la Sociedad Española de Urgencias de Pediatría reportó un caso clínico de un recién nacido con anillo de constricción, en unión 1/3 medio y distal del antebrazo derecho, compatible con bridas amnióticas (30). En Argentina se ha reportado el caso clínico de una niña de 8 meses de edad, con hallazgos típicos del síndrome de bridas amnióticas en mano derecha; los dedos 2º, 4º y 5º unidos, configurando una pseudosindactilia; además con amputaciones distales de los dedos 2º y 3º, y en el tercio inferior de la pierna izquierda existía un anillo de constricción circunferencial con edema (31). En México se ha reportado el manejo anestesiológico de una adolescente de 17 años de edad con síndrome de bridas amnióticas que requirió tratamiento odontológico extensivo bajo anestesia general, la cual presentaba hundimiento parietal, exoftalmos, hipertelorismo, implantación baja de los pabellones auriculares, puente nasal aplanado y desviado, obstrucción parcial de las narinas, hipoplasia malar bilateral, micrognatia, paladar ojival alto, protrusión dental y edema de las encías (32). En Barranquilla (Colombia) se ha reportado un caso de síndrome de bridas amnióticas por exposición a misoprostol (33). En el estudio se describe a un paciente masculino de dos meses de edad; hijo de padres sanos; con amputaciones y oligodactilia a nivel de miembros inferiores y superiores, y a nivel de miembros inferiores con anillos de constricción.
En general, las corrientes etiológicas del síndrome de bridas amnióticas apoyan, por un lado, a las malformaciones debido a defectos citogenéticos (34) y, por otra parte, los productos pretérmino, la ruptura prematura de membranas corio-amnióticas, la incompetencia cervical, cirugía del cérvix, amniocentesis, gestación múltiple como factores importantes que se asocian a este síndrome (35, 36). Aunque, al parecer, la teoría más aceptada para explicar las malformaciones en estos pacientes fue la propuesta por Torpin en 1965, quien mencionó que la ruptura prematura del amnios, según la etapa de la gestación, produce efectos disruptivos en el producto, y cuando la ruptura es muy temprana, el defecto se propaga rápido y puede llegar a la zona donde el cordón umbilical se inserta en la placenta (37).
IMPORTANCIA DE LOS PROCESOS MOLECULARES EN LA DINÁMICA EMBRIONARIA
Los recientes avances en el conocimiento de los mecanismos moleculares que controlan la dinámica embrionaria aportan sobre cuestiones cruciales en la biología del desarrollo. Se ha venido planteando en este trabajo el interés, en el campo de la biología del desarrollo, por develar el misterio de cómo un huevo fertilizado, que inicia al poco tiempo su segmentación dentro de la dinámica embrionaria, empieza a transformarse en un organismo multicelular bien sea un pez, anfibio, reptil, ave o mamífero, incluyendo en este grupo al humano. Esto lleva a plantearse algunas preguntas sobre este mecanismo tan complejo, entre las que se pueden mencionar: ¿Cómo se coordinan las células del embrión para formar, por ejemplo, una aleta, ala, brazo, etc., en los cordados? ¿Cuál es el mecanismo causante de la disposición precisa en la formación de huesos, músculos, nervios, etc.?
Según Pisanó (8), a lo largo de la historia del siglo XX estas mismas preguntas han sido planteadas por investigadores como Spemann en 1935, Seidel en 1952, Chang en 1954, Dalc en 1967 y His en 1984. De acuerdo con esto, Bostuoabad y Pisanó (2) postulan la "teoría de las localizaciones germinales", según la cual, las futuras partes del embrión existirían en estadios muy precoces como territorios órgano-formativos y como prueba de la totipotencia ovular en mamíferos. En este sentido, Seidel en 1952 (8) realizó un estudio interesante, cuyos resultados mostraron cómo los dos primeros blastómeros de conejo, luego de haber sido separados, dieron cada uno origen a un embrión normal. Un resultado similar se obtuvo con el desarrollo del primero de los dos blastómeros irregulares, mantenidos en explante, en Bufo arenarum (38).
El desarrollo de la proteína verde fluorescente (GFP, por sus siglas en inglés), como un marcador para observar la expresión de los genes, es un avance científico que ha sido ampliamente utilizado en el campo de la biología del desarrollo y las ciencias biomédicas en general (39 - 41). Hace más de una década que se demostró su aplicación en el marcaje de estructuras biológicas, como la identificación de neuronas en el nemátodo Caenorhabditis elegans (39), por lo que esta proteína se ha utilizado también en el estudio de diferentes procesos a nivel celular (42), para observar cómo migran las células cancerosas hasta llegar a hacer metástasis (43), así como para analizar la actividad de las células embrionarias y observar cómo conquistan nuevos espacios hasta lograr su diferenciación en órganos específicos (44).
Los estudios de la genética clásica y molecular han permitido el descubrimiento de muchos genes que participan en los bien regulados programas del desarrollo y que dan origen a organismos tan complejos como los vegetales y animales (45). Así que la comprensión de la base molecular para la acción de estos genes es una de las metas más perseguidas por la biología tanto en el pasado siglo como en el presente.
Los genes Hox, descubiertos en la mosca de la fruta Drosophila melanogaster (46), constituyen una familia de genes que codifican factores de transcripción que regulan muchos procesos del desarrollo, a menudo de una manera combinatoria. Determinadas mutaciones en esos genes afectan, por ejemplo, la ubicación de los primordios de las extremidades en cordados; así como mutantes para el gen Hoxb5 presentan desplazamiento de ciertas estructuras de las extremidades respecto de las posiciones normales en el flanco embrionario. El estudio de los genes Hox en muchas especies de los diversos grupos animales ha establecido que el complejo homeótico es una característica universal de los organismos multicelulares y que este complejo existía ya hace unos 540 millones de años durante el Cámbrico inferior, por lo que se plantea una relación estrecha causal entre la explosión del Cámbrico y la aparición del complejo Hox (47).
Se ha observado que los genes nodal juegan un papel importante en la regulación de la dinámica embrionaria. Estos genes se encuentran en todos los vertebrados y su expresión regula el patrón de diferenciación del cigoto mediante interacciones moleculares complejas. Las proteínas nodal, una subclase dentro de la superfamilia de proteínas conocidas como TGF-ß, están relacionadas con la inducción del mesodermo, el desarrollo de la porción rostral del SNC y la determinación de la asimetría derecha- izquierda del corazón y de otros órganos de los vertebrados (48, 49). De manera parecida, el HNF-3ß (Factor Nuclear Hepático) mantiene el nódulo primitivo y, más tarde, induce la especificidad regional en las áreas del prosencéfalo y mesencéfalo. Sin HNF- 3B, los embriones no consiguen gastrular adecuadamente y carecen de estructuras prosencefálicas y mesencefálicas (9). Con respecto a la región central y caudal del embrión, la formación del mesodermo dorsal está regulada por el gen brachyury (T), que se expresa en el "nódulo", las células precursoras de la notocorda. Este gen es esencial para la migración de las células a través de la línea primitiva (50).
Por otro lado, el estudio de mutaciones en humanos y en ratones confirma que Pax6 juega un papel crítico en la formación del ojo y que la haploinsuficiencia para Pax6 en humanos causa aniridia, desorden caracterizado por hipoplasia del iris y de la fóvea, acompañado de catarata, incapacidad corneal y glaucoma progresivo (51).
Finalmente, entender los mecanismos por los cuales se generan los distintos tipos celulares durante el desarrollo embrionario representa uno de los aspectos más fascinantes de la biología del desarrollo. Se ha establecido que dos mecanismos principales son los responsables de establecer la organización básica del embrión: la localización de determinantes citoplasmáticos y la señalización entre células (52). Con relación a los casos expuestos en este trabajo, la no expresión de algunos genes, entre los que estarían los genes nodal, podría ser la causa de los diversos tipos de malformaciones presentadas.
Financiación: Universidad del Norte.
Conflicto de interés: Ninguno.
REFERENCIAS
(1) De Robertis EM, Oliver G, Wright CV. Genes con homeobox y el plan corporal de los vertebrados. Temas "Investigación y Ciencia": construcción de un ser vivo. Investigación y Ciencia 1998; 92-94. [ Links ]
(2) Bostuoabad O, Pisanó A. A explant of the first four micromeres of Bufo arenarum eggs. Acta Embryologiae Experimentalis 1971; 1: 225-37. [ Links ]
(3) Bostuoabad O, Pisanó A. Explante de los primeros macrómeros en huevos de Bufo arenarum. Acta Embryologiae Experimentalis 1974; 1: 77-86. [ Links ]
(4) Bostuoabad O, Pisanó A. Diferenciación de áreas blastulares explantadas de Bufo arenarum. Acta Embryologiae Experimentalis 1974; 1: 67-75. [ Links ]
(5) Falco P, Restrepo S. Desarrollo embrionario de Bufo marinus (Amphibia Anura): Tabla de desarrollo normal. Anuario Científico Universidad del Norte 1983; 2: 109-19. [ Links ]
(6) Cervantes LE, Falco GP. Tabla de desarrollo larvario normal de Bufo marinus (Amphibia Anura). Anuario Científico Universidad del Norte 1984; 3: 153-65. [ Links ]
(7) Solari AJ. Genética del desarrollo normal y del desarrollo de tumores. Genética Humana. 2ª ed. Buenos Aires: Editorial Médica Panamericana; 2000. p. 251-252. [ Links ]
(8) Pisanó A. Tópicos de Embriología. Buenos Aires: FECC editores; 1977. [ Links ]
(9) Sadler TW. Tercera semana del desarrollo. Lagman Embriología Médica. 11ª ed. Baltimore: Lippincott Wiliams & Wilkins; 2009. p. 61- 63. [ Links ]
(10) Lugones B, Pichis G, Ramírez B, Miyiar P. Sirenomelia. Rev. Cubana Med Gen Integr 2006; 22 (2): 37. [ Links ]
(11) Rodríguez MA, Carmona ML, Chacin PB, Tovitto G, Colina A, Moreno F. Sirenomelia. Rev Obst Ginecol 2007; 67 (3): 1-11. [ Links ]
(12) Solano AF, Saldarriaga W, Isaza C, Mastroiacovo P, Castilla E. Foco epidémico de sirenomelia en Cali (Colombia). Informe de 4 casos en el Hospital Universitario del Valle en 54 días. Colombia Médica 2006; 37 (3): 1-18. [ Links ]
(13) Flórez CC, Villagomez GE, Soria JA, Asombriz R, Iglesias JL. Teratoma Orofaríngeo. Comunicación de un caso. Rev Med Univ (México) 2009; 11 (43): 129-33. [ Links ]
(14) Nieto MA, Meneses MA, Cervantes LE. Teratoma de la línea a media craneal. Reporte de un caso. Revista Universidad del Quindío 2004; 10: 79-85. [ Links ]
(15) Saiga T, Osasa H, Hatamaya H, Miyamoto T, Ono H, Mikami T. The origin of extragonadal teratoma: case report of an inmature teratoma ocurring in a prenatal brain. Fetal Pediatr Pathol 1991; 11 (5): 759-70. [ Links ]
(16) Shere DM, Abramowics JS, Eggers PC, Metlay LA, Sinkin RA, Woods JR. Prenatal ultrasonigraphic diagnosis of intracranial teratoma and massive craniomegaly with associated higr-out put cardiac failure. Rev Am J Obstet Gynecol 1993; 168 (1): 97-9. [ Links ]
(17) Digiovanni LM, Sheik Z. Prenatal diagnosis clinical significance and magnagement of fetal intracranial teratoma: a case report and literature review. Rev Ann J Perinatal 1994; 11 (69): 420-22. [ Links ]
(18) Kuller JA, Laifer SA, Marti JG, MacPherson TA, Hill LM. Unusual presentations of fetal teratoma. Rev J Perinatal 1991; 113: 294-6. [ Links ]
(19) Brun J, Portugal O, Tamayo L, Fernández R, Moreno B. Teratoma maduro cráneofacial derecho: Presentación de un caso clínico inusual. Cuad Hosp Clin 2007; 52 (1): 64-68. [ Links ]
(20) Islas L, García S, Palma E, Cruz J. Amputación fetal por bandas amnióticas de una de las extremidades. Rev Mex Pediatr 2010; 77 (3): 119-22. [ Links ]
(21) Hermann J, Optiz JM. Naming and nomenclature of syndromes. In: Birth Defects, Original Article Series. The National Fundation March of dimes 1974; 10 (7): 82. [ Links ]
(22) Cervantes LE, Ruiz O, Nieto MA, Rengifo RL, Meza OF. Extrofia Cloacal por Bridas Amnióticas: reporte de un caso en el departamento del Quindío. Algunas consideraciones embriológicas. Revista Investigaciones Universidad del Quindío 1994; 5: 17-26. [ Links ]
(23) Rengifo RL, Ochoa LM, Meza OF, Cervantes LE, Castaño JJ. Amputaciones congénitas por bridas amnióticas. Presentación de siete casos. Revista Española de Pediatría 1991; 47 (5): 441-46. [ Links ]
(24) Witters I, Moerman P, Louwagie D, Van Assche FA, Migeon BR, Fryng JP. Second trimester prenatal diagnosis of epignatus teratoma in ring X chromosome mosaicism with inactive ring X chromosome. Ann Genet 2001; 44: 179-82. [ Links ]
(25) Pino RV, Mogollón CT, Pardo JM, Rojas UE, Blanco HA. Teratoma. Acta Otorrinolaringológica 2003; 54: 305-8. [ Links ]
(26) Haghighi K, Miller M, Cleveland D, Ziccardi V. Epignathus teratoma with bifid tongue and median glossal salivary mase. Report of a case. J. Oral Maxillofac Surg 2004; 62: 379- 83. [ Links ]
(27) Orioli IM, Ribeiro MG, Castilla EE. Clinical and epidemiological studies of amniotic deformity adhesion and mutilation (ADAM) sequence in a South American (ECLAMC) population. Am J Med.Genet 2003; 118-35. [ Links ]
(28) Da Silva G, Cammarata SF, Gonzalez CM, La Cruz MA. Síndrome de bridas amnióticas. Revista Chilena de Pediatría 2008; 79 (2): 172-180. [ Links ]
(29) Pons GA, Saez AR, Sepúlveda LW. Bridas amnióticas, sinequia intrauterina y tabique mulleriano. Etiopatogenia, diagnóstico diferencial y pronóstico. Rev Chil Ultrasonog 2005; 8: 51-8. [ Links ]
(30) De la Peña A, Pérez JD, Farrases O, Rubio LM, Marín J, González D. Bridas amnióticas. A propósito de un caso. Revisión Anual de la Sociedad Española de Urgencias de Pediatría, Murcia, 18 de abril de 2008. Disponible en: www.codem.es/Docs/Noticias/poster_bridas_amnioticas.pdf [ Links ]
(31) Bibas BH, Atar FM, Espíndola E. Síndrome de bridas amnióticas. Arch Argent Pediatr 2002; 100: 240-5 [ Links ]
(32) Reyes-Aveleyra M, Borunda D, Whizar-Lugo VM, Anzorena-Vallarino J. Síndrome de Bridas Congénitas. Consideraciones Anestésicas. Rev Anestesia Méx 2008; 20 (3). Disponible en: http://www.fmca.org.mx/art/art.php?id=584 [ Links ]
(33) Hernández E, Silvera LA, Garavito P, Rolong G, Ordóñez J, Barrios R, Silvera C. Síndrome de bridas amnióticas por exposición a misoprostol: presentación de un caso clínico. Rev Asoc Col Ciencias Biol 2010; 22 (1): 163-4. [ Links ]
(34) Philipp T, Philipp K, Reiner A, Beer F, Kalousek DK. Embryoscopic and cytogenetic analysis of 233 missed abortions: factors involved in the pathogenesis of developmental defects of early failed pregnancies. Hum Reprod 2003; 18: 1724-32. [ Links ]
(35) Corona-Rivera JR, Corona Rivera E, Navarro- Ramírez P, Castro-Hernández JF, Acosta- León J, Corona-Rivera A. Lumbopedal skin pedicle in an infant with the amniotic band syndrome: a disorganization-like defect. Clin Dysmorphol 2003; 12: 51-4. [ Links ]
(36) Craven C, Ward K. Placental causes of fetal malformation. Clin Obstet Gynecol 1996; 3: 588-606. [ Links ]
(37) Torpin R. Amniochorionic mesoblastic fibrous strings and amniotic bands associated constricting fetal malformations of fetal death. Am J Obstet Gynecol 1965; 91:65-75. [ Links ]
(38) Cervantes LE, Paz AD, Pisanó A. Developmental of the first two irregular blastomeres of Bufo arenarum maintained in explante. Comunicaciones Biológicas 1990; 8 (3): 271-82. [ Links ]
(39) Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC. Green Fluorescent protein as a marker for gene expression. Science 1994; 263: 802-5. [ Links ]
(40) Shaner NC, Patterson GH, Davidson MW. Advances in fluorescent protein technology. J Cell Science 2007; 120 (24): 4247-60. [ Links ]
(41) Yeh E, Gustafson K, Boulianne G. Green fluorescent protein as a vital marker and reporter of gene expression in Drosophila. Proc Natl Acad Sci USA 1995; 92: 7036-40. [ Links ]
(42) Gerdes H, Kaether C. Green fluorescent protein: applications in cell biology. FEBS Letters 1997; 389: 44-7. [ Links ]
(43) Hoffman RM. In vivo imaging of metastatic cancer with fluorescent proteins. Cell Death and Diferentiation 2002; 9: 786-789. [ Links ]
(44) Takeuchi Y, Yoshizaki G, Kobayashi T, Takeuchi T. Mass isolation of primordial germ cells from transgenic rainbow trout carrying the green fluorescent protein gene driven by the vasa gene promoter. Biol Reprod 2002; 67: 1087-92. [ Links ]
(45) Lodish H, Berk A, Zipursky SW, Matsudaira P. Baltimor, Darnell J. Biología Celular y Molecular. 4ª ed. Buenos Aires: Editorial Médica Panamericana; 2002. p. 537-538. [ Links ]
(46) Capdevila J, Rodríguez Esteban C, Izpisúa Belmonte JC. Arquitectura Molecular de la Extremidad de Vertebrados. Revista Investigación y Ciencia 1998; 26-37. [ Links ]
(47) Morata G. La historia de los genes homeóticos. Arbor CLXVIII 2001; 662: 229-46. [ Links ]
(48) Sampath K, Rubinstein AL, Cheng AM, Liang JO, Fekany K, Solnica-Krezel L et al. Induction of the zebrafish ventral brain and floorplate requires cyclops/nodal signaling. Nature 1998; 395 (6698): 185-9. [ Links ]
(49) Dougan ST, Warga RM, Kane DA, Schier AF, Talbot WS. The role of the zebrafish nodal related genes squint and cyclops in patterning of mesendoderm. Development 2003; 130 (9): 1837-51. [ Links ]
(50) Beddington RSP, Rashbass P, Wilson V. Brachyury - a gene affecting mouse gastrulation and early organogenesis. Development 1992; 157-165. [ Links ]
(51) Wawersik S, Maas RL. Vertebrate eye development as modeled in Drosophila. Human Molecular Genetics 2000; 9 (6): 917-25. [ Links ]
(52) Larrain J, Brown C, De Robertis EM. Integrin- a3 mediates binding of Chordin to the cell surface and promotes its endocytosis. EMBO Reports 2003; 4: 813-8. [ Links ]
