










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Salud Uninorte
versão impressa ISSN 0120-5552
Salud, Barranquilla vol.31 no.1 Barranquilla jan./avr. 2015
Antígenos asociados a tumores y su potencial uso en el tratamiento del cáncer
Tumor associated antigens and its potential use in cancer therapy
Luz Adriana Sarmiento-Rubiano1
1 Bacterióloga, Ph.D. Investigadora, Laboratorio de Investigación Universidad Metropolitana de Barranquilla - Fundación Hospital Universitario Metropolitano.
Correspondencia: Luz Adriana Sarmiento-Rubiano. Cra. 42F n° 75 B -18, piso 12. Barranquilla (Colombia). lusarru@hotmail.com
Fecha de recepción: 9 de agosto de 2014
Fecha de aceptación: 7 de noviembre de 2014
Resumen
De acuerdo con cifras de la Organización Mundial de la Salud (OMS) en su reporte mundial de cáncer 2008, la incidencia de esta enfermedad se duplicó en las últimas tres décadas del siglo XX, y a pesar de los importantes esfuerzos de la ciencia por combatir este flagelo y de la creación de nuevas estrategias terapéuticas, se calcula que para 2050 la mitad de la población estará en riesgo de desarrollar algún tipo de cáncer. El rápido desarrollo de la tecnología en las últimas décadas ha permitido dilucidar algunos de los mecanismos moleculares y de las interacciones de la célula tumoral con el sistema inmunológico, que conllevan finalmente al desarrollo o eliminación del tumor y en los que la estructura antigénica de este juega un papel fundamental. El descubrimiento de una gran variedad de antígenos asociados a tumores ha permitido el desarrollo de inmunoterapias que mediante el uso de anticuerpos monoclonales o de vacunas para inmunización activa se han convertido en una nueva estrategia con interesantes y alentadoras perspectivas en el tratamiento de esta enfermedad. En este artículo se describen las características básicas de algunos de estos antígenos, su influencia en el desarrollo tumoral y su actual aplicación en inmunoterapias, con el fin de generar en el lector interés y expectativas acerca de este promisorio futuro terapéutico contra el cáncer.
Palabras clave: cáncer, antígenos, inmunoterapia, marcadores tumorales, vacuna.
Abstract
According to the World Health Organization data in the World Cancer Report 2008, the cancer incidence was duplicated in the last three decades of the 20th century and in spite of the important efforts of the science for attacking this scourge and of the creation of new therapeutic strategies, it is calculated that for the year 2050, the half of the population will be in risk of developing some cancer type. The rapid development of the technology in the last decades has allowed explaining some of the molecular mechanisms and the interactions of the tumor cell with the immune system, that finally produces the tumor development or elimination and on the tumor antigenic structure plays a fundamental role. The discovery of variety of tumors associated antigens has allowed the immunotherapy development with the use of monoclonal antibodies or vaccines for active immunization, these news strategies represent important trends in cancer treatment. In this paper there are described some of these antigens in its basic characteristics, influence in the tumor development and current application in immunotherapy, in order to generate in the reader expectations about this future cancer therapy.
Keywords: cancer, antigens, immunotherapy, tumoral markers, vaccine.
Introducción
Cuando se habla de cáncer se hace referencia a un grupo de enfermedades caracterizadas por la proliferación rápida de células anormales con capacidad de invadir cualquier órgano y en algunos casos propagarse, generando nuevos centros de proliferación denominados "metástasis".
De acuerdo con cifras de la Organización Mundial de la Salud (OMS) en su reporte mundial de cáncer de 2008, de 12,66 millones de personas diagnosticadas con cáncer en el mundo, 7,6 millones murieron, y de estas, el 70 % habitaba en países de ingresos bajos y medios. La OMS destaca que la prevención, detección oportuna y adecuado manejo son fundamentales para reducir y controlar este importante problema de salud pública, y debe ser un compromiso de los gobiernos y la sociedad aunar esfuerzos para lograr este propósito (1).
Desde los descubrimientos de madame Curie y los inicios de la radioterapia hasta la actual terapia génica han sido muchos y muy diversos los esfuerzos de la humanidad por estudiar, controlar y tratar de curar el cáncer. La estrecha relación que hoy se conoce entre el tumor y el sistema inmunológico fue ini-cialmente planteada en los años 20 por Paul Erlich y posteriormente confirmada experimentalmente por Gross Foley y Prehn en los años 50, con sus experimentos de resistencia al tumor en ratones previamente inmunizados, y por Burnet y su teoría de la supervivencia inmune de los tumores (2).
Actualmente se sabe que existen moléculas que son expresadas en las células cancerígenas de forma selectiva, aberrante o en cantidades muy superiores en relación con las células normales y que muchas de estas moléculas tienen capacidad inmunogénica, convirtiéndose en interesantes blancos inmu-nológicos capaces de generar una respuesta que permita la eliminación del tumor. Estos antígenos, denominados comúnmente Antí-genos Asociados a Tumores (AAT), tienen en la mayoría de los casos actividades biológicas relacionadas con la proliferación celular, inhibición de la apoptosis, adherencia de las células tumorales y metástasis, por lo que una respuesta inmunológica dirigida contra ellos podría generar la destrucción tumoral o tener un efecto inhibitorio de su actividad mediante el bloqueo por anticuerpos de sitios biológicamente activos.
Muchos de estos antígenos han sido descubiertos gracias a la presencia espontánea de anticuerpos en pacientes con tumores que los expresan, indicando que han dejado de ser reconocidos como propios, por ser lo suficientemente diferentes antigénicamente, porque se expresan en cantidades significativamente mayores en las células tumorales o porque están ausentes en los tejidos normales. Una de las técnicas más utilizada para la identificación de estos antígenos es la llamada SEREX (Serological Screening of cDNA Expression Library); este método consiste en descubrir anticuerpos en sueros de pacientes con cáncer, al enfrentarlos con proteínas recombinantes obtenidas a partir del cDNA de sus propios tumores (3).
En los últimos veinte años y gracias a los avances en las técnicas de investigación y en el conocimiento del sistema inmunológico, son muchos los AAT que han sido descubiertos, y más diversas aun son las terapias que han sido planteadas para su utilización como blanco inmunológico en el tratamiento del cáncer.
En este artículo se pretende describir las características de algunos de estos antígenos, por considerarlos relevantes por su universalidad, ya que están presentes en un alto porcentaje de tumores; porque han sido ampliamente estudiados y las experiencias terapéuticas llevadas a cabo en diferentes estudios clínicos con ellos nos pueden permitir una adecuada visualización del panorama actual en el desarrollo de vacunas y otras inmunoterapias para el tratamiento de variados tipos de cáncer.
Receptores de la familia tirosina quinasa
La familia de receptores tirosina quinasa son un grupo de proteínas que se expresan normalmente en algunos tipos celulares y poseen la capacidad de transferir un grupo fosfato a partir de ATP a grupos hidroxilo de proteínas presentes en el citoplasma celular, y así intervenir en diferentes vías de señalización relacionadas con la proliferación,supervivencia y metabolismo celular. En general, estas proteínas están compuestas por tres dominios: (i) dominio extracelular, donde se encuentran los ligandos que interaccionan con diversas moléculas como factores de crecimiento, citoquinas u hormonas, entre otras; (ii) dominio transmembrana, encargado de la transmisión de la señal cuando sucede la dimerización del receptor, y (iii) domino citoplasmático, en el cual se activa uno o varios residuos quinasa que interaccionan con moléculas que intervienen en diferentes vías de señalización celular (4).
Los genes que codifican para miembros de la familia tirosina quinasa en humanos pueden clasificarse en 20 subfamilias basados en la secuencia del dominio quinasa; una de estas subfamilias es la EGFR (Epidermal Growth Factor Receptor), que incluye los receptores EGFR/HER1, ErbB-2/HER2, ErbB-3/HER3 y ErbB-4/HER4 (5), involucrados principalmente en las vías de señalización MAPK (proteínas qui-nasas activadas por mitógenos), la ruta PI3K (fosfatidilinositol- 3-kinasa), que contribuye a la progresión del ciclo celular, disminuye la apoptosis y promueve la metástasis de células cancerígenas, y las vías JAK/STAT y C gamma-1 fosfolipasa (PLCyl), relacionadas con la proliferación, diferenciación, migración y apoptosis celular (6). Los receptores de esta familia se encuentran frecuentemente implicados en neoplasias de animales y humanos y una sola célula tumoral puede sobre expresar más de un tipo de estos receptores.
El receptor HER2 ha sido uno de los AAT más estudiados; se encuentra sobreexpresado hasta en el 60 % de los cánceres de pulmón, 39 % de mama, 40 % de estómago, 20 % de colon, 64 % de esófago, 80 % de próstata, 50 % de vejiga, 32 % de ovario, entre otros (7). La sobreexpresión de HER2 en las células cancerígenas ha sido asociada a mayor agresividad del tumor, aumento de metástasis y disminución en la supervivencia del paciente, principalmente en cáncer de mama (8). También se ha asociado a la resistencia al tratamiento con medicamentos como el Tamoxifen (9) y a menor supervivencia postquirúrgica en pacientes con cáncer co-lorrectal (10). Sin embargo, no en todos los cánceres que sobreexpresan este receptor se ha logrado establecer una relación directa con pronósticos negativos, más aun cuando se evalúa como único indicador (11), por ello cada día se generan más controversias ante el valor pronóstico de HER2.
Aunque HER2 está sobreexpresado en variados tipos de cáncer (hasta 2 000 000 de copias en la superficie celular), se sabe que también se encuentra normalmente en células de las membranas epiteliales gastrointestinal, respiratoria, urinaria, reproductiva, seno y placenta, en mínimas cantidades en el adulto (menos de 20 000 copias por célula) y en mayor proporción en el mismo tejido durante el periodo fetal.
La presencia de HER2 en tejido normal supone una tolerancia y reconocimiento como propio de esta proteína a nivel inmunológico, sin embargo, en sueros de pacientes con cáncer de mama HER2 positivo se encontraron anticuerpos contra epítopes específicos de HER2 tanto del dominio extracelular como intracelular (12), lo que supone una diferencia inmuno-lógicamente reconocible del receptor sobre expresado en las células cancerígenas, que permite su uso como blanco inmunológico en la eliminación de tumores, sin un efecto autoinmune contra el tejido normal.
En 1998 la Agencia de Alimentos y Medicamentos de los Estados Unidos de América(FDA) aprobó el uso de Transtuzumab (Her-ceptin), un anticuerpo monoclonal humanizado tipo IgG1 producido por tecnología de DNA recombinante, para ser utilizado en el tratamiento de pacientes con cáncer de mama metastásico HER2 positivo (13). Este anticuerpo se une a dos sitios específicos del dominio extracelular de HER2, inhibiendo su dimerización y estimulando la lisis celular mediada por anticuerpos (14). Posteriormente, y en respuesta a la resistencia al tratamiento de algunos pacientes, fueron creados Pertuzumab (15), Cetuximab y Matuzumab(16) , anticuerpos dirigidos contra epítopes diferentes pero con similar actividad.
Una segunda estrategia terapéutica basada en HER-2 consiste en el diseño de moléculas capaces de bloquear su actividad fosforila-sa; el Imatinib, por ejemplo, es un inhibidor competitivo de los receptores tirosina qui-nasa PDGF (factor de crecimiento derivado de plaquetas) y c-Kit, que regulan el crecimiento y división celular y cuyo aumento es fundamental en el desarrollo del cáncer(17) ; Nilotinib, Desatinib y Lapatinib son bloqueadores de segunda generación de los receptores tirosina kinasa (18). In vitro se ha demostrado que el uso simultáneo de Lapa-tinib y transtuzumab aumenta la capacidad de destrucción en diferentes líneas celulares de cáncer de mama (19).
Mucina asociada a la superficie celular MUC
Las mucinas asociadas a la superficie celular (MUC) son una familia de glicoproteínas expresadas normalmente en muchos tipos de células epiteliales y cuya función es promover la supervivencia celular y la homeostasis con su medio ambiente local, permitir la interacción celular con diferentes moléculas de su entorno y activar algunas vías de señalización celular. La alteración en la expresión o glicosilación de estas proteínas ha sido observada en muchos tipos de cáncer, en los que se cree favorece la proliferación tumoral y metástasis (20). Las proteínas de esta familia han sido clasificadas en dos grupos: las que están unidas a la superficie celular y las que son secretadas y se encuentran libres en el espacio intercelular (21). De esta familia, MUC1, MUC4 y MUC16 son las proteínas que han sido mejor caracterizadas; MUC1 es una glicoproteína transmembrana de 500 a 1000 kDa, su base peptídica tiene dos subunida-des, la subunidad C terminal, que incluye un pequeño dominio extracelular, un dominio transmembrana y un dominio citoplasmático, del cual se sabe que es sitio de unión para diferentes moléculas (22), y la segunda, una subunidad extracelular N terminal altamente glicosilada, que consiste en la repetición de secuencias de 20 aminoácidos con cinco posibles sitios de O-glicosilación, con un valor de repetición entre 25 y 125 veces (23). La proteína MUC1 está sobreexpresada y abe-rrantemente glicosilada en más del 90 % de los canceres de mama y en un alto porcentaje de otros tipos de cáncer.
Aunque no se han dilucidado completamente las funciones de esta familia de proteínas, se sabe que la fosforilación del dominio cito-plasmático de MUC1 activa diferentes vías de señalización al interactuar con factores como: (i) beta-catenina, molécula involucrada en la capacidad de adhesión celular y en la activación transcripcional de genes relacionados con la proliferación celular (24); (ii) P53, activador transcripcional de genes pro-apoptosis y del que MUC1 es un importante represor (25); (iii) HER2, receptor tirosina quinasa que al dimerizar con MUC1 permite la activación de la vía MAPK, regulando funciones de proliferación y diferenciación celular (26). Por su parte, MUC4 actúa como ligando en la formación de heterodímeros HER2/HER3, con la consecuente fosforilación y activación de las vías de señalización (27). MUC16 se encuentra principalmente en el epitelio de la superficie ocular, tracto respiratorio superior, mesotelio de algunas cavidades y en los órganos reproductores, cumpliendo funciones de protección y lubricación de estas superficies; MUC16, conocido también como CA125, es utilizado como marcador celular en cáncer de ovario, teniendo su elevación en sangre un importante valor en el seguimiento terapéutico de estos pacientes (28). Aunque no se conoce la actividad de la región citoplasmática de MUC16, sí se sabe que su región extracelular es capaz de bloquear la formación de la sinapsis inmunitaria entre los linfocitos natural killer y la célula cancerígena de ovario, impidiendo la lisis de la célula tumoral (29).
La glicoproteína MUC-1 es sintetizada como una cadena polipeptídica simple, a la que se le unen N-glicanos en la región transmembrana durante su paso por el retículo endoplasmáti-co y es altamente O-glicocilada y parcialmente sializada a su paso por el aparato de Golgi, la proteína finalmente es anclada en la superficie celular, gracias a la acetilación de los ácidos grasos de membrana por las cisteínas presentes en la región extracelular de MUC1 (30). La O-glicosilación es catalizada por las N-acetilgalactosamina (GalNAc)-transferasas; anormalidades en estas enzimas están asociadas a la formación de glicoproteínas aberrantes presentes en diferentes enfermedades, y es posible que este mecanismo también ocurra en el caso del cáncer (31).
Se ha observado la presencia de MUC1 soluble en el sobrenadante de cultivos de células cancerígenas in vitro y en algunos fluidos corporales in vivo, por lo cual se ha planteado la posibilidad de que el dominio extracelular de esta glicoproteína sea enzimáticamente liberado por algún mecanismo proteolítico, y se ha considerado a TACE (Tumor necrosis factor a converting enzyme) como el posible responsable de esta actividad (32).
El gen que codifica para la proteína MUC1 (cromosoma 1q21-24) presenta un promotor con un alto nivel de complejidad en el que existen variados elementos cis de regulación transcripcional controlados por diversas moléculas; esta variedad en la regulación es la responsable de las diferentes isoformas de MUC1 presentes en un mismo tipo celular (33).
MUC1 ha sido utilizado como marcador tumoral en seguimiento terapéutico en pacientes con cáncer de mama, conocido como el marcador CA15.3; se encuentra elevado 10 a 15 veces en pacientes con cáncer metastásico, aunque su valor pronóstico solo es importante cuando se analiza junto con otros parámetros (34).
Jerome y colaboradores observaron en los ganglios linfáticos cercanos al tumor de pacientes con cáncer pancreático y de mama la existencia de linfocitos T citotóxicos con capacidad de eliminar tumorales, gracias al reconocimiento del epítope PDTRP del dominio extracelular de MUC1 (35). Anticuerpos tipo IgM y IgG contra MUC1 han sido encontrados en sueros de mujeres multíparas sanas; generados posiblemente por un aumento en la producción de MUC1 durante el embarazo y la lactancia, y se cree que estos anticuerpos tienen un efecto protector capaz de reducir el riesgo de cáncer de mama (36), por lo que se podría considerar a MUC1 como un adecuado blanco para el desarrollo de inmunotera-pias en cáncer. Vale destacar que MUC1 está aberrantemente glicosilado en las células cancerígenas, por lo que su estructura difiere ampliamente del expresado en células normales, así que un anticuerpo dirigido contra el MUC1 aberrante de la célula cancerígena posiblemente no afecte el tejido normal.
Diferentes inmunoterapias han sido diseñadas basadas en la proteína MUC1, como los anticuerpos monoclonales humanizados huBrE-3 y R-1549 (pemtumomab), utilizados en tratamientos combinados de radioinmuno-terapia en cáncer de mama y otros cánceres con promisorios resultados (37, 38).
La inmunización activa a través del diseño de vacunas también ha utilizado a MUC1 como blanco inmunológico.
Immucin es una vacuna basada en un péptido sintético de MUC1 que expresa varios epítopes capaces de ser presentados por moléculas del complejo mayor de histocompatibilidad clase I y II y estimular los linfocitos CD8+ y CD4+, utilizando GM-CSF (factor estimulante de colonias granulocíticas) como adyuvante (39).
Stimuvax o L-BLP25 es una vacuna liposomal basada en un péptido sintético de 25 aminoácidos que contiene la secuencia repetitiva del dominio extracelular de MUC1, unida a una estructura lipídica (coleterol, dimiristoil fos-fatidilglicerol y dipalmitoil fosfatidilcolina) y al lípido A como adyuvante (40).
MUC1-KLH plus QS-21 es una vacuna que conjuga una secuencia de 30 aminoácidos del dominio extracelular de MUC1 con KLH (Keyhole Limpet Hemocyanin) como molécula portadora y QS-21 como adyuvante; la inmunización con esta vacuna en pacientes con cáncer de mama produjo altos títulos de anticuerpos tipo IgM y IgG, capaces de reconocer el epítope APDTRPA de MUC1 en células tumorales (41,42).
Otra vacuna peptídica fue diseñada utilizando los epítopes de muc1 rpapgs, ppahgvt y pdtrp; este último glicosilado aberran-temente en la treonina y utilizando como adyuvante pam3cyssk4 (43); panvac es una vacuna basada en los virus vaccinia y virus de la viruela aviar que contienen transgenes de muc1 y del antígeno carcinoembrionario, así como de moléculas inmunoestimulato-rias como b7-1 (cd80), icam-1 (cd54) y lfa-3 (cd58); la infección de células dendríticas con estos virus recombinantes desencadena una reacción inmunológica capaz de eliminar células tumorales que sobreexpresen estos antígenos (44).
Estas vacunas se encuentran actualmente en ensayo clínico fase ii o iii, con alentadoras perspectivas terapéuticas.
Antígenos testiculares de cáncer (ATC)
Los antígenos testiculares de cáncer (atc) son una categoría de antígenos que se expresan en gran variedad de tumores, pero no en los tejidos normales, a excepción de la placenta y las células germinales masculinas, de donde deriva su nombre; por esta razón son considerados importante blanco inmunológico para la eliminación de tumores. Estos antígenos pueden ser clasificados de acuerdo con la ubicación de los genes que los codifican en dos grupos: los que son codificados por genes presentes en el cromosoma X y los que no (45). Los atc más frecuentemente expresados en tumores son magea1, magea3, ny-eso-1,ssx-2 y ssx-4.
Los antígenos asociados a melanoma (mage) pertenecientes a la categoría atc son una familia de proteínas clasificadas de acuerdo con el nivel de conservación de la secuencia central de aminoácidos en dos tipos: mage tipo I, al que pertenecen los subgrupos mage-a, mage-b y mage-c (46), que se expresan en células tumorales de pulmón, cabeza, cuello, vejiga, mama, cáncer gástrico y colorrectal, entre otros (47, 48, 49); y mage tipo ii, al que pertenecen los subgrupos mage-d, mage-e, mage-f, mage-g, mage-h, mage-l2 y Necdin, que son codificadas por genes presentes en los cromosomas X, 3 y 15 y que se expresan en algunos tejidos normales y su función está relacionada con procesos de ubiquitinación de proteínas (46, 50).
La expresión de las proteínas mage tipo I, al igual que la de otros atc, está inhibida en el tejido normal por mecanismos epigenéticos de represión transcripcional por hipermetila-ción del gen promotor; diversos autores han demostrado que la acetilación a nivel de las histonas y la desmetilación del dna juegan un papel importante en la expresión de estas proteínas en las células tumorales (51, 52). A nivel funcional mage-Aactúa como inhibidor de p53, al interferir con su función de regulador transcripcional de genes relacionados con la reparación o eliminación apoptótica de células dañadas, favoreciendo la formación del tumor (53).
El gen mage a-3 codifica un antígeno presente en un alto porcentaje de tumores, principalmente en melanomas, y ha sido uno de los más importantes blancos inmunológicos en el desarrollo de terapias contra el cáncer; este antígeno posee epítopes capaces de ser presentados por moléculas hla i y hla ii (54).
La empresa GlaxoSmithKline ha patentado una vacuna que contiene la proteína recombinante mage a3 unida a la proteína D de Haemophilus influenzae como acompañante de fusión y as02b (monofosforil lípido a-qs21)como adyuvante; esta vacuna ha demostrado aumentar la sobrevida libre de enfermedad en pacientes postquirúrgicos con melanoma estadio II y se encuentra actualmente en estudio clínico fase III (55).
NY-ESO-1 es otro de los antígenos ATC considerado importante candidato para inmuno-terapia; su expresión ha sido evidenciada por RT-PCR de mRNA en el 30 % de los cánceres de pulmón, vejiga y melanomas (56), 42 % de tumores de mama (57) y 65 % de carcinomas medulares de tiroides (58), entre otros.
La frecuencia de anticuerpos anti-NY-ESO-1 en sueros de pacientes con tumores que expresan este gen es del 25 al 50 % y se ha observado una relación directa entre la evolución del tumor y el título de estos anticuerpos. La presencia de anticuerpos en sueros de pacientes con tumores NY-ESO-1 positivos demuestra la capacidad de este antígeno para inducir una respuesta inmunológica mediada por linfocitos CD4+ y CD8+ (59).
El antígeno NY-ESO-1 ha sido utilizado en diversas inmunoterapias contra el cáncer. Adams y colaboradores, por ejemplo, observaron una importante reacción humoral y celular en pacientes con melanoma maligno a quienes se inyectó intradérmicamente la proteína recombinante NY-ESO-1 en un área de piel previa y posteriormente tratada con Imiquimod, una amina imidazoquinolina sintética inmunomoduladora de aplicación tópica, que actúa como ligando de receptores tipo Toll (TLR7), generando una respuesta inmune local (60).
Aunque el uso de adyuvantes tópicos resulta una alternativa segura y novedosa, títulos mayores de anticuerpos fueron obtenidos cuando la proteína recombinante NY-ESO-1 se inyectó intramuscularmente con el adyuvante isocomatrix en pacientes con melanoma (61) o con adyuvantes como Montanide ISA-51 y CpG 7909 (62).
Telomerasa
La telomerasa es una enzima con actividad transcriptasa reversa, encargada de añadir repeticiones teloméricas de novo para mantener la longitud del telómero en diversos tejidos. Esta proteína se expresa en células con importante actividad replicativa, como células madre, tejido embrionario, hematopoyético y germinal, pero no en otras células somáticas, en las que su ausencia causa acortamiento de los telómeros durante las sucesivas divisiones celulares, envejecimiento y muerte celular.
La evidencia de la reactivación de la actividad telomerasa en muchos tejidos tumorales se ha considerado una causa de la inmortalidad de estas células. Estructuralmente, la telomerasa está integrada por un complejo ribonucleoproteína que consiste en una proteína transcriptasa reversa (hTERT) y un componente RNA (hTR) que sirve como molde a la secuencia telomérica TTAGGG (63).
Se ha demostrado que la expresión de la proteína hTERT en células somáticas que no la expresan genera un alargamiento en los telómeros, lo cual sugiere que la actividad telomerasa está regulada por la expresión de esta proteína.
Aunque no se han dilucidado completamente los mecanismos de regulación transcripcional de hTERT, se sabe que existen diferentes activadores y represores, entre los que se incluye el regulador c-Myc y mecanismos de regulación epigenéticos mediante metilación y acetilación génica (64).
Diferentes métodos han sido utilizados para inhibir la actividad telomerasa como posible tratamiento contra el cáncer; teniendo en cuenta que esta enzima se expresa en la mayoría de cánceres humanos, la telomerasa se ha considerado uno de los blancos terapéuticos contra el cáncer más universales (65).
El componente hTR, que contiene la región molde de la secuencia telomérica, es susceptible de hibridación con oligonucleótidos complementarios capaces de inhibir su actividad; es el caso del lípido modificado GRN163L (3-mer oligonucleótido N3'-P5' tiofos-foramidato - 5'-TAGGGTTAGACAA-3'), también conocido como Imetelstat, un potente inhibidor competitivo de la unión al telómero del enzima, que ha demostrado ser eficaz en el acortamiento de telómeros y muerte de células tumorales en pacientes con cánceres hematológicos y tumores sólidos. Imetelstat se encuentra actualmente en estudio clínico fase II (66, 67).
Por su parte, la proteína hTERT ha demostrado tener capacidad inmunogénica y epítopes reconocibles por los linfocitos T citotóxicos en células tumorales, mas no en células normales que también expresan esta proteína, como las CD34+; esto posiblemente debido a la excesiva cantidad de este antígeno presente en células tumorales (68).
Variados péptidos de hTERT han sido estudiados como posibles blancos para inmuno-terapia en cáncer. El péptido hTERT (ILAKFL-HWL), que es normalmente presentado en moléculas HLA A-2 de células tumorales, fue uno de los primeros y más controversiales respecto a los resultados de los ensayos, ya que para algunos autores la vacunación con células dendríticas estimuladas con este péptido es capaz de generar una respuesta inmune con actividad antitumoral (69),mientras otros consideran que este péptido no es presentado por las células tumorales y no puede ser utilizado en inmunoterapia (70). El péptido GV1001, que corresponde a la secuencia (EARPALLTSRLRFIPK), es capaz de generar una respuesta inmune mediada por linfocitos CD4+ y CD8+; este péptido ha sido utilizado para vacunación en combinación con GM-CSF como adyuvante y ha demostrado inducir una respuesta inmune en pacientes con melanomas, carcinoma de pulmón y adenocarcinoma pancreático, entre otros, y ha mejorado la sobrevida del paciente con cáncer cuando se utiliza adicional a la terapia tradicional (71, 72).
Antígeno carcinoembrionario
Cuando se habla de Antígeno Carcinoembrio-nario (CEA) se hace referencia a una familia de proteínas de superficie perteneciente a la superfamilia de las inmunoglobulinas;. La familia CEA está compuesta por dos subgrupos: el de las glicoproteínas (PSG) y el de las moléculas de adhesión (CEACAM) (73). Estructuralmente, los miembros del grupo CEACAM están compuestos por varios dominios extracelulares de inmunoglobulina anclados o no a la membrana celular por una molécula de glicosilfosfatidilinositol (GPI); algunos tienen dominios transmembrana y otros, como CECAM1, tienen además dominios citoplasmáticos con residuos de serina, treoni-na y tirosina, que al ser fosforilados participan en procesos de señalización celular (74).
CEACAM1 es el más ampliamente expresado y estudiado miembro de la familia CEA; se encuentra normalmente en la superficie de algunas células epiteliales, en células endo-teliales y en células del sistema inmunoló-gico como las natural killer, linfocitos B y T, macrófagos y células dendríticas, donde cumple funciones de adhesión, comunicación celular, receptor de patógenos y control de la angiogénesis, entre otras (74).
CEACAM5, otro conocido miembro de esta familia, está compuesto por un dominio extra-celular de siete inmunoglobulinas ancladas a la membrana celular por una molécula de GPI; esta proteína tiene un peso molecular de 180 kDa y es altamente glicosilada, se expresa normalmente en abundantes cantidades en el epitelio colónico durante el desarrollo embrionario y en míninas cantidades en el adulto.
CEACAM5, llamado también CD66e, se conoce desde los años sesenta como el marcador tumoral CEA, convirtiéndose en el primer marcador tumoral utilizado en el mundo; sus niveles en sangre se encuentran aumentados en más del 70 % de los cánceres humanos, incluidos cáncer gástrico, pulmonar, mama, pancreático, ovárico y próstata, entre otross(75).
CEACAM6, conocido como NCA90, está compuesto por dos dominios extracelulares de inmunoglobulina y un anclaje a membrana GPI; al igual que CECAM5, está sobreexpresado en muchos tipos de cáncer, y ambos (CEA-CAM5 y CEACAM6) se asocian con procesos de adhesión celular y metástasis.
Algunos autores consideran que CEACAM6 puede ser utilizado como marcador tumoral en el seguimiento de pacientes con cáncer colorrectal y que altos niveles de este marcador están asociados a peor pronóstico, metástasis y acortamiento de la sobrevida de los pacientes (76).
La relación directa de CEACAM5 y CEACAM6 con procesos de adhesión y metástasis en células tumorales ha generado el diseño de anticuerpos dirigidos contra epítopes
específicos ubicados en sitios de unión en la región N terminal de estas moléculas, capaces de bloquear su actividad in vitro e in vivo, disminuyendo procesos metastásicos, con aumento de la sobrevida en modelos animales con cáncer (77, 78).
Aunque CEACAM5 se encuentra expresado en mínimas cantidades en el epitelio colónico adulto normal, su sobreexpresión en células tumorales ha permitido que esta molécula sea blanco inmunológico para el diseño de diversas vacunas.
La inmunización con una proteína recombinante de CEACAM5 y GM-CSF como adyuvante ha generado una respuesta inmune específica celular y humoral en pacientes con cáncer de colon (79).
El diseño de vacunas basadas en el uso de virus recombinantes para la expresión de transgenes del CEA ha permitido la efectiva inmunización de pacientes con avanzados carcinomas CEA positivos (80).
Virus como el vaccinia, capaz de infectar y replicarse en las células presentadoras de antígeno, para que estas puedan procesar y presentar epítopes específicos del antígeno CEA a los linfocitos T en moléculas HLA I y II (81), o como el canarypox , un poxvirus de aves que a diferencia del virus vaccinia no se replica en las células humanas, solo las infecta y expresa sus transgenes, favoreciendo la presentación antigénica (82), han sido utilizados en diferentes modelos de inmunoterapia.
Aunque los estudios clínicos de estas vacunas se encuentran actualmente en fase II, su uso es adicional a las terapias convencionales, y los resultados han permitido solo un aumento en la sobrevida de los pacientes.
Otros antígenos
Existen otros antígenos que también han sido blanco inmunológico en el desarrollo de terapias contra el cáncer; algunos de ellos son: (i) El hexa-sacárido Globo H, un glu-coesfingolípido que está sobreexpresado en tumores de colon, ovario, páncreas, pulmón, próstata y mama; este antígeno, producido sintéticamente y utilizado como vacuna con el adyuvante alfa-galactosilceramida, produce anticuerpos contra Globo-H y contra SSEA3, su pentasacárido precursor, el cual es sobreexpresado en cáncer de mama (83,84). (ii) El gen WT1 codifica para una proteína que actúa como regulador transcripcional de las proteínas anfiregulina, bcl2-2 y E-cadherina, implicadas en procesos de crecimiento, adherencia y apoptosis celular; la expresión anormal del gen WT1 se ha observado en leucemias y en tumores sólidos como el nefroblastoma de Wilms; péptidos sintéticos de esta proteína con el adyuvante ISA51 han sido empleados como vacuna, y han mostrado promisorios resultados (85, 86). (iii) El gen P53 codifica una proteína que actúa como regulador transcripcional de genes involucrados en los procesos de reparación celular, senescencia y apoptosis, así como en la promoción de la respuesta inmune antitumoral mediante la regulación transcripcional de los ligandos ULBP1 y ULBP2 para los linfocitos natural killer en la célula tumoral (87); P53 está alterado y sobreexpresado en un alto porcentaje de cánceres, lo que junto a su capacidad inmu-nogénica ha permitido que sea utilizado en el diseño de vacunas (88).
En general son cientos los AAT que han sido descubiertos y que podrían por sus características individuales ser utilizados en el desarrollo de inmunoterapias. Estos antígenos han sido objeto de diversas clasificaciones; una de ellas está basada en su modelo y nivel de expresión y en la presencia de mutaciones, e incluye cinco grupos: (i) Antígenos testiculares de cáncer, como los antígenos MAGE1 y NY-ESO1; (ii) Antígenos de diferenciación, presentes exclusivamente en un tipo de tejido, como la tirosinasa de los melanosomas; (iii) Antígenos específicos de tumor, originados por mutaciones específicas, como algunas mutaciones en P53; (iv) Antígenos sobreexpresados, como HER2, y (v) Antígenos oncofétales, que se expresan en la fase de desarrollo embrionario pero no en el tejido adulto, como el CEA (89).
Expectativas terapéuticas basadas en AAT
En los últimos años ha sido creciente el interés en la búsqueda de nuevos AAT y en el desarrollo de terapias basadas en ellos; prueba de esto es que de acuerdo con el informe de la FDA sobre medicinas y vacunas en desarrollo de 2013, de un total de 907 posibles nuevos medicamentos, 338 son terapias para el cáncer, y de estos, 170 están basados en anticuerpos monoclonales y más de 80 son vacunas que se encuentran en estudio clínico fase II o III (90).
Existen diferentes estrategias para el diseño de vacunas con fines terapéuticos en cáncer (figura 1); algunas utilizan células dendríticas (CDs) de pacientes con cáncer, generadas ex vivo a partir de células CD34+ estimuladas con células o extractos celulares de tumor, epítopes específicos, péptidos sintéticos o proteínas recombinantes de AAT; estas CDs al ser reinyectadas generan una respuesta inmune celular y humoral capaz de combatir el tumor.
Estos modelos de vacunas autólogas, aunque laboriosas y costosas por lo personificadas, han mostrado ser eficaces en la inmunización,ya que una estimulación controlada de las CDs permite establecer parámetros inmu-nológicos ideales, lo cual evita reacciones de tolerancia inmunológica (91).
Otras vacunas incluyen péptidos sintéticos, proteínas recombinantes o transgenes de AAT en partículas virales acompañados de variados adyuvantes; entre los que se destaca el GM-CSF, molécula capaz de inducir la proliferación, maduración y migración de CDs y la diferenciación de linfocitos B y T, lo cual genera una importante inmunidad antitumor demostrada en animales y humanos (92).
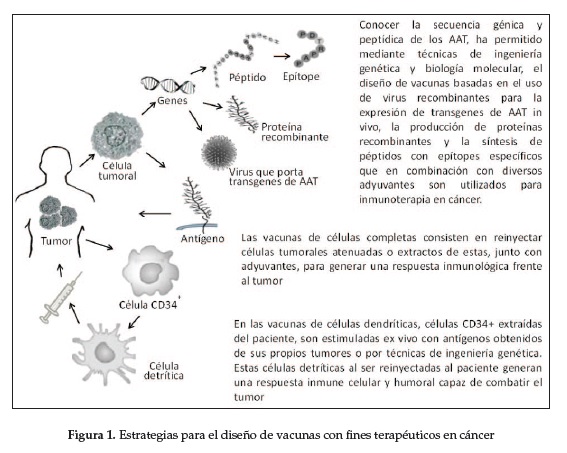
La inmunoterapia pasiva basada en el uso de anticuerpos monoclonales de origen externo dirigidos contra epítopes específicos de AAT resulta ideal cuando una inmunización activa podría generar el riesgo de un efecto autoinmune; tal es el caso de algunos epíto-pes específicos de AAT, que aunque están sobreexpresados en la célula tumoral, también están presentes en menores cantidades en tejido normal.
Estos anticuerpos también han sido utilizados como mecanismos localizadores de tumor y transportadores de isotopos radioactivos o medicamentos, que son liberados en el microambiente tumoral como una estrategia terapéutica combinada.
Algunos ejemplos de terapia pasiva y de vacunas fueron descritos en detalle con los antígenos incluidos en esta revisión.
Cuando se plantea la posibilidad de la vacunación como terapia para la eliminación de tumores se hace referencia a la activación del sistema inmune para que pueda reconocer y destruir al tumor, utilizando todos los mecanismos celulares y humorales necesarios para este propósito, los cuales incluyen: inducir una inmunidad celular con linfocitos T cito-tóxicos específicos contra el tumor; disminuir la actividad de los linfocitos T supresores antígeno específicos; producir una memoria humoral antígeno específica y estimular la producción de citoquinas mediadoras de lisis tumoral (93).
La destrucción inmune de las células tumorales es un proceso que debe presentarse en condiciones fisiológicas normales, así que la formación del tumor podría considerarse como una derrota del sistema inmunológico frente a la célula tumoral.
Actualmente se sabe que los AAT juegan un papel fundamental en el reconocimiento y destrucción de las células transformadas, por ser el punto diferencial entre estas y sus antecesoras normales. Schreiber y colaboradores plantean la existencia de un proceso de "inmunoedición", en el que la interacción del tumor en sus primeras etapas de formación con el sistema inmunológico podría determinar sus características inmunogénicas. Este proceso de "inmunoedición" tendría básicamente tres fases: (i) eliminación: etapa inicial en la que el sistema inmunológico innato y adaptativo procura eliminar las células transformadas; (ii) equilibrio: en esta etapa las células transformadas que no lograron ser eliminadas permanecen latentes, pero su crecimiento es controlado por el sistema inmunológico adaptativo; esta interacción por un periodo relativamente largo de laten-cia puede generar tolerancia inmunológica frente a algunos AAT o causar nuevas modificaciones en la célula tumoral que permitan su adaptación; por ello es en esta etapa en la que se produce la edición inmune del tumor propiamente dicha; (iii) escape: en esta etapa las células tumorales pueden proliferar y generar un tumor clínicamente evidente, gracias a que han logrado evadir el sistema inmunológico durante la etapa de equilibrio, o a la existencia de una deficiencia inmune temporal o permanente (93).
En la última década se han dilucidado los mecanismos por los cuales procesos de inflamación crónica asociada al microambiente tumoral promueven la angiogénesis, la liberación de factores que estimulan la proliferación celular y la tolerancia inmunológica, lo cual favorece el crecimiento tumoral y la metástasis (94). También se ha observado que células del sistema inmunológico asociadas al microambiente tumoral expresan mayor cantidad de receptores, como el CTLA4 (Cytotoxic Tlymphocyte-associated antigen 4) y PD1 (programmed cell death protein 1), implicados en la regulación negativa de la activación de los linfocitos y en la tolerancia inmunológica.
El bloqueo de los receptores CTLA4 mediante anticuerpos dirigidos contra ellos o sus li-gandos favorece la inmunidad antitumoral; es por ello que la FDA ha aprobado el uso de algunos de estos anticuerpos, como el Ipili-mumab, para la terapia complementaria en cáncer (95, 96).
En este contexto podría decirse que el tumor tiene la capacidad "aprendida" de evadir el sistema inmune durante los procesos de proliferación celular, crecimiento tumoral y metástasis; que la estructura antigénica del tumor fue modelada por un proceso de "inmunoedición" que le permite ser tolerada o que es ineficientemente reconocida por el sistema inmunológico para su destrucción, más aun dentro del microambiente tumoral; y que la célula tumoral se vale de mecanismos inmunológicos para favorecer su desarrollo. Es por ello que el éxito de una inmunoterapia para la eliminación del tumor requiere de una minuciosa y adecuada selección de los AAT a los que está dirigida y al direccionamiento de una respuesta inmune rápida y eficaz en la que se disminuya el riesgo de generar tolerancia inmunológica.
Conclusiones
Actualmente el mayor entendimiento de la inmunogenicidad del tumor y la dinámica de sus relaciones con el sistema inmunológico ha permitido el desarrollo de terapias contra el cáncer basadas en el uso de anticuerpos o en el diseño de vacunas, en las que el descubrimiento y detallado conocimiento de los AAT juega un papel fundamental. Sin embargo, en la mayoría de estas terapias (las cuales generalmente han sido complementarias a los esquemas tradicionales de tratamiento) solo ha sido posible un aumento en la sobrevida de los pacientes.
Es fundamental continuar en el propósito de mejorar y diversificar la terapia inmuno-lógica, combinando no solo la búsqueda de nuevos AAT, sino también de mecanismos de inmunomodelación y control de la respuesta, que permitan bloquear las vías de tolerancia inmunológica y controlar los microambientes inflamatorios asociados al tumor que puedan beneficiar el crecimiento de este.
Aunque la mayoría de los estudios realizados para determinar la eficacia de la inmu-noterapia en cáncer han sido realizados en pacientes con estadios clínicos avanzados de la enfermedad y en combinación con tratamientos quimio o radioterapéuticos, que por su naturaleza citotóxica afectan también las células del sistema inmunológico, resultaría muy interesante conocer el efecto de estas terapias en etapas tempranas, posteriores a un diagnóstico oportuno, o incluso como medida profiláctica en pacientes de alto riesgo de padecer cáncer; en tal caso deberían estar dirigidas a AAT con características muy específicas de universalidad (expresados en un alto porcentaje de tumores), especificidad (exclusiva o aberrantemente expresados en células tumorales) e inmunogenicidad, la cual va a estar muy influenciada por la estrategia de presentación antigénica, evitando la tolerancia inmunológica.
Respecto a las ventajas del uso de los AAT como blancos terapéuticos, indudablemente el efecto selectivo de la inmunoterapia sobre las células tumorales y la baja toxicidad de los tratamientos reportada en los diferentes estudios clínicos resulta el mayor beneficio de esta en relación con los tratamientos convencionales de radio y quimioterapia, con una elevada citotoxicidad que afecta de manera importante las células sanas, principalmente las de mayor actividad mitótica como las del sistema inmunológico.
Vale la pena destacar que por la especificidad del tratamiento es fundamental hacer una identificación completa de los antígenos presentes en las células tumorales para cada paciente; más aun cuando se conoce que mediante los procesos de inmuno modulación del tumor y tolerancia inmunológica, un determinado clon celular metastásico o no puede tener modificaciones en la expresión de AAT en relación con el tumor original, presentando, por consiguiente, resistencia a la inmunoterapia inicial, indiferente de si esta es pasiva o activa, lo que podría considerarse una desventaja.
Respecto a los costos y facilidad de producción de las inmunoterapias, estos pueden variar mucho y resultar desde muy costosos y laboriosos como las vacunas autólogas personalizadas, basadas en células dendríticas estimuladas in vitro y reinoculadas al paciente, hasta las basadas en péptidos sintéticos de producción en masa y a bajo costo.
El creciente interés en la investigación y el uso de la inmunoterapia en la prevención y tratamiento oportuno del cáncer podría resultar una estrategia clave para combatir este flagelo del siglo XXI.
Conflicto de interés: ninguno.
Financiación: Universidad Metropolitana de Barranquilla.
Referencias
1. Emal A, Bray F, Ferlay J. Global Cancer Statistics. CA Cancer J Clin 2011; (2): 69-90. [ Links ]
2. Scharovsky OG, Matar P, Fluck MZ, Rico MJ, Rabinovich GA. From immune surveillance to tumor-immune escape : the story of an enemy with multiple strategies. Inmunology 2006; 25(1): 101-14. [ Links ]
3. Sahin U, Tureci O, Schmitfr H, Cochlovius B, Johannes T, Schmits R et al. Human neo-plams elicit multiple specific inmune response in the autologous host. Inmunology 1995; 92: 11810-3. [ Links ]
4. Kasture VS, Musmade DS, Aher SJ, Patil PP. Tyrosine Kinases : promising targets for cancer chemotherapy. Med Chem Drug Discov. 2012; 2(2): 37-51. [ Links ]
5. Robinson DR, Wu YM, Lin SF. The protein tyrosine kinase family of the human genome. Oncogene 2000; 19(49): 5548-57. [ Links ]
6. Ross JS, Fletcher JA, Bloom KJ, Linette GP,Stec J, Symmans WF et al. Targeted therapy in breast cancer: the HER-2/neu gene and protein. Mol. Cell. Proteomics 2004; 3(4): 37998. [ Links ]
7. Normanno N, Bianco C, De Luca A, Maiello MR, Salomon DS. Target-based agents against ErbB receptors and their ligands: a novel approach to cancer treatment. Endocr. Relat. Cancer 2003; 10(1): 1-21. [ Links ]
8. Kumar V, Tewari M, Singh U. Significance of Her-2 / neu protein over expression in Indian breast cancer patients. Indian J. Surg 2007; (69): 122-8. [ Links ]
9. Ross J, Fletcher J. The HER-2/neu Oncogene in Breast Cancer: Prognostic Factor, Predictive Factor, and Target for Therapy. Oncologist 1998; 3(4): 237-52. [ Links ]
10. Park D Il, Kang MS, Oh SJ, Kim HJ, Cho YK, Sohn C Il et al. HER-2/neu overexpression is an independent prognostic factor in colo-rectal cancer. Int. J. Colorectal Dis 2007; 22(5): 491-7. [ Links ]
11. Zhou F, Li N, Jiang W, Hua Z, Xia L, Wei Q et al. Prognosis significance of HER-2/ neu overexpression/amplification in Chinese patients with curatively resected gastric cancer after the ToGA clinical trial. World J. Surg. Oncol 2012; 10(1): 274-282. [ Links ]
12. Disis ML, Calenoff E, McLaughlin G, Murphy a E, Chen W, Groner B et al. Existent T-cell and antibody immunity to HER-2/neu protein in patients with breast cancer. Cancer Res 1994; 54(1): 16-20. [ Links ]
13. Glennie M J, Johnson P W M. Clinical trials of antibody therapy. Immunology today 2000;21(8): 403-410. [ Links ]
14. Hudis C A. Trastuzumab-mechanism of action and use in clinical practice. N. Engl. J. Med 2007; 357(1): 39-51. [ Links ]
15. Sakai K, Yokote H, Murakami-Murofushi K, Tamura T, Saijo N, Nishio K. Pertuzumab, a novel HER dimerization inhibitor, inhibits the growth of human lung cancer cells mediated by the HER3 signaling pathway. Cancer Sci 2007; 98(9): 1498-503. [ Links ]
16. Kamat V, Donaldson JM, Kari C, Quadros MRD, Lelkes PI, Chaiken I et al. Enhanced EGFR inhibition and distinct epitope recognition by EGFR antagonistic mAbs C225 and 425. Cancer Biol. Ther 2008; 7(5): 726-33. [ Links ]
17. Zhang P, Gao WY, Turner S, Ducatman BS. Gleevec (STI-571) inhibits lung cancer cell growth (A549) and potentiates the cisplatin effect in vitro. Molecular Cancer 2003; 9: 1-9. [ Links ]
18. Soverini S, Colarossi S, Gnani A, Castag-netti F, Rosti G, Bosi C et al. Resistance to dasatinib in Philadelphia-positive leukemia patients and the presence or the selection of mutations at residues 315 and 317 in the BCR-ABL kinase domain. Haematologica 2007; 92(03): 401-404. [ Links ]
19. Konecny GE, Pegram MD, Venkatesan N, Finn R, Yang G, Rahmeh M et al. Activity of the dual kinase inhibitor lapatinib (GW572016) against HER-2-overexpressing and trastuzumab-treated breast cancer cells.Cancer Res 2006; 66(3): 1630-1639. [ Links ]
20. Hollingsworth MA, Swanson BJ. Mucins in cancer: protection and control of the cell surface. Nat. Rev. Cancer 2004; 4(1): 45-60. [ Links ]
21. Chauhan SC, Kumar D, Jaggi M. Mucins in ovarian cancer diagnosis and therapy. J. Ovarian Res 2009; 2: 21-29. [ Links ]
22. Yang E, Hu XF, Xing PX. Advances of MUC1 as a target for breast cancer immunotherapy. Histol Histopathol 2007; 2: 905-922. [ Links ]
23. Song W, Delyria ES, Chen J, Huang W, Lee JS, Mittendorf E a et al. MUC1 glycopeptide epitopes predicted by computational glyco-mics. Int. J. Oncol 2012; 41(6): 1977-1984. [ Links ]
24. Yamamoto M, Bharti A, Li Y, Kufe D.Interaction of the DF3/MUC1 breast carcinoma-associated antigen and beta-catenin in cell adhesion. J. Biol. Chem 1997; 272(19):12492-12494. [ Links ]
25. Wei X, Xu H, Kufe D. Human mucin 1 on-coprotein represses transcription of the p53 tumor suppressor gene. Cancer Res. 2007;67(4): 1853-1858. [ Links ]
26. Singh PK, Hollingsworth MA. Cell surface-associated mucins in signal transduction. Trends Cell Biol 2006; 16(9): 467-476. [ Links ]
27. Kozloski Goldi A, Carraway Coralie A. Ca-rothers; Carraway Kermit L. Mechanistic and signaling analysis of Muc4-ErbB2 signaling module: New insights into the mechanism of ligand independent ErbB2 activity. J. Cel. Physiology 2010; 224 (3): 649-657. [ Links ]
28. Wang F, Ye Y, Xu X, Zhou X, Wang J, Chen X. CA-125-indicated asymptomatic relapse confers survival benefit to ovarian cancer patients who underwent secondary cyto-reduction surgery. J. Ovarian Res 2013; 6(1):14-25. [ Links ]
29. Gubbels JAA, Felder M, Horibata S, Belisle J A, Kapur A, Holden H et al. MUC16 provides immune protection by inhibiting synapse formation between NK and ovarian tumor cells. Mol. Cancer 2010; 9: 11-25. [ Links ]
30. Parry S, Silverman HS, McDermott K, Willis A, Hollingsworth MA, Harris A. Identification of MUC1 Proteolytic Cleavage Sites in Vivo. BBRC 2001; 283(3): 715-720. [ Links ]
31. Bennett EP, Mandel U, Clausen H, Gerken T, Fritz T, Tabak L. Control of mucin-type O-glycosylation: a classification of the po-lypeptide GalNAc-transferase gene family. Glycobiology 2012; 22(6): 736-756. [ Links ]
32. Thathiah A, Blobel CP, Carson DD. Tumor necrosis factor-alpha converting enzyme/ADAM 17 mediates MUC1 shedding. J. Biol. Chem 2003; 278(5): 3386-3394. [ Links ]
33. Zaretsky JZ, Barnea I, Aylon Y, Gori-vodsky M, Wreschner DH, Keydar I. MUC1 gene overexpressed in breast cancer: structure and transcriptional activity of the MUC1 promoter and role of estrogen receptor alpha (ERalpha) in regulation of the MUC1 gene expression. Mol. Cancer 2006; 5:57-71. [ Links ]
34. Harris L, Fritsche H, Mennel R, Norton L, Ravdin P, Taube S et al. American Society of Clinical Oncology 2007 update of recommendations for the use of tumor markers in breast cancer. J. Clin. Oncol 2007; 25(33):5287-5312. [ Links ]
35. Jerome KR, Barnd DL, Bendt KM, Boyer CM, Taylor-papadimitriou J, Mckenzie IFC et al. Cytotoxic T-Lymphocytes Derived from Patients with Breast Adenocarcinoma Recognize an Epitope Present on the Protein Core of a Mucin Molecule Preferentially Expressed by Malignant Cells. Cancer Res 1991;51:2908-2916. [ Links ]
36. Von Mensdorff-Pouilly S, Moreno M, Ver-heijen RHM. Natural and Induced Humoral Responses to MUC1. Cancers 2011; 3(4):3073-3103. [ Links ]
37. Murawa P, Kobylarek R, Gracz A, Malicki J. Intraperitoneal administration radio- labelled monoclonal antibody Pertumomab (YTTRIUM-90-HMFG1) in gastric Cancer. Rep. Pract. Oncol. Radiother 2003; 8 (2): 112-120. [ Links ]
38. Kramer EL, Liebes L, Wasserheit C, Noz E, Zabalegui A. Initial clinical evaluation of radiolabeled MX-DTPA humanized BrE-3 antibody in patients with advanced breast cancer . Clin Cancer Res 1998; 4 (7):1679-1688. [ Links ]
39. Kovjazin R, Volovitz I, Kundel Y, Rosenbaum E, Medalia G, Horn G et al. ImMucin: a novel therapeutic vaccine with promiscuous MHC binding for the treatment of MUC1-expressing tumors. Vaccine 2011; 29(29-30): 4676-4686. [ Links ]
40. Sangha R, Butts C. L-BLP25: a peptide vaccine strategy in non small cell lung cancer. Clin. Cancer Res 2007; 13(15 Pt 2): 4652-4654. [ Links ]
41. Roulois D, Gregorie M, Fonteneau Jean-François. MUC1-specific cytotoxic T lymphocytes in cancer therapy: induction and challenge. BioMed Res. Inter 2012; 87-93. [ Links ]
42. Adluri S, Gilewski T, Zhang S, Ramnath V, Ragupathi G, Livingston P. Specificity analysis of sera from breast cancer patients vaccinated with MUC1-KLH plus QS-21. Br. J. Cancer 1999; 79(11-12): 1806-1812. [ Links ]
43. Lakshminarayanan V, Thompson P, Wolfert MA, Buskas T, Bradley JM, Pathangey LB et al. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. U. S. A. 2012;109(1): 261-266. [ Links ]
44. Arlen PM, Pazdur M, Skarupa L, Rauc-khorst M, Gulley JL. A Randomized Phase II Study of Docetaxel Alone or in Combination with PANVAC™V (Vaccinia) and PAN-VAC™ F (Fowlpox) in Patients with Me-tastatic Breast Cancer (NCI 05-C-0229). Clin. Breast Cancer 2006; 7(2): 176-179. [ Links ]
45. Fratta E, Coral S, Covre A, Parisi G, Colizzi F, Danielli R et al. The biology of cancer tes-tis antigens: putative function, regulation and therapeutic potential. Mol. Oncol 2011;5(2): 164-82. [ Links ]
46. Chomez P, Backer O De, Bertrand M, Plaen E De, Boon T, Lucas S. An Overview of the MAGE Gene Family with the Identification of All Human Members of the Family. Cancer Res 2001; 61 (14): 5544-51. [ Links ]
47. Jungbluth AA, Busam KJ, Kolb D, Iversen K, Coplan K, Chen YT et al. Expression of MAGE-antigens in normal tissues and cancer. Int. J. Cancer 2000; 85(4): 460-465. [ Links ]
48. Mori M, Inoue H, Mimori K, Shibuta K, Baba K, Nakashima H et al. Expression of MAGE genes in human colorectal carcinoma. Ann. Surg 1996; 224(2): 183-188. [ Links ]
49. Otte M, Zafrakas M, Riethdorf L, Pichlmeier U, Lo T, Ja F. MAGE-A Gene Expression Pattern in Primary Breast Cancer. Cancer Res 2001; 61(18): 6682-6687. [ Links ]
50. Doyle JM, Gao J, Wang J, Yang M, Potts PR.MAGE-RING protein complexes comprise a family of E3 ubiquitin ligases. Mol. Cell 2010; 39(6):963-974. [ Links ]
51. Wischnewski F, Pantel K, Schwarzenbach H. Promoter demethylation and histone acetylation mediate gene expression of MA-GE-A1, -A2, -A3, and -A12 in human cancer cells. Mol. Cancer Res 2006; 4(5): 339-349. [ Links ]
52. De Smet C, Lurquin C, Lethe B, Martelange V, Boon T. DNA methylation is the primary silencing mechanism for a set of germ line-and tumor-specific genes with a CpG-rich promoter. Mol. Cell. Biol 1999; 19(11): 7327-35. [ Links ]
53. Marcar L, Maclaine NJ, Hupp TR, Meek DW. Mage-A cancer/testis antigens inhibit p53 function by blocking its interaction with chromatin. Cancer Res 2010; 70(24): 10362-10370. [ Links ]
54. Schultz ES, Lethe B, Cambiaso CL, Lymphocytes CDCT, Lethe B, Heirman C et al. A MAGE-A3 Peptide Presented by HLA-DP4 Is Recognized on Tumor Cells by CD4 + Cyto-lytic T Lymphocytes Advances. Cancer Res 2000; 60: 6272-6275. [ Links ]
55. Atanackovic D, Altorki NK, Cao Y, Ritter E,Ferrara CA, Ritter G et al. Booster vaccination of cancer patients with MAGE-A3 protein reveals long-term immunological memory or tolerance depending on priming. Proc. Natl. Acad. Sci. U.S.A. 2008; 105(5):1650-1655. [ Links ]
56. Jungbluth AA, Chen YT, Stockert E, Busam KJ, Kolb D, Iversen K et al. Immunohisto-chemical analysis of NY-ESO-1 antigen expression in normal and malignant human tissues. Int. J. Cancer 2001; 92(6):856-860. [ Links ]
57. Sugita Y, Wada H, Fujita S, Nakata T, Sato S, Noguchi Y et al. NY-ESO-1 Expression and Immunogenicity in Malignant and Benign Breast Tumors NY-ESO-1. Cancer Res 2004;64:2199-2204. [ Links ]
58. Maio M. Analysis of Cancer/Testis Antigens in Sporadic Medullary Thyroid Carcinoma: Expression and Humoral Response to NY-ESO-1. J. Clin. Endocrinol. Metab 2003;88(2): 748-754. [ Links ]
59. Jäger E, Stockert E, Zidianakis Z, Chen YT, Karbach J, Jäger D et al. Humoral immune responses of cancer patients against "Can-cer-Testis" antigen NY-ESO-1: correlation with clinical events. Int. J. Cancer 1999; 84(5):506-510. [ Links ]
60. Adams S, Neill DWO, Nonaka D, Hardin E,Chiriboga L, Siu K et al. Imiquimod as Vaccine Adjuvant. J Immunol 2008; 181: 776-784. [ Links ]
61. Davis ID, Chen W, Jackson H, Parente P, Shackleton M, Hopkins W et al. Recombinant NY-ESO-1 protein with ISCOMATRIX adjuvant induces broad integrated antibody and CD4(+) and CD8(+) T cell responses in humans. Proc. Natl. Acad. Sci. U.S.A. 2004; 101(29): 10697-10702. [ Links ]
62. Valmori D, Souleimanian NE, Tosello V, Bhardwaj N, Adams S, O'Neill D et al. Vaccination with NY-ESO-1 protein and CpG in Montanide induces integrated antibody/ Th1 responses and CD8 T cells through cross-priming. Proc. Natl. Acad. Sci. U.S.A.2007;104(21): 8947-8952. [ Links ]
63. Zhang Q, Kim N-K, Feigon J. Architecture of human telomerase RNA. Proc. Natl. Acad.Sci. U.S.A. 2011; 108(51): 20325-20332. [ Links ]
64. Renaud S, Loukinov D, Abdullaev Z, Guilleret I, Bosman FT, Lobanenkov V et al. Dual role of DNA methylation inside and outside of CTCF-binding regions in the transcriptio-nal regulation of the telomerase hTERT gene.Nucleic Acids Res. 2007; 35(4): 1245-1256. [ Links ]
65. Shay JW, Bacchetti S. A survey of telomerase activity in human cancer. Eur. j. cancer 1997;33 (5): 787-791. [ Links ]
66. Joseph I, Tressler R, Bassett E, Harley C, Buseman CM, Pattamatta P et al. The telo-merase inhibitor imetelstat depletes cancer stem cells in breast and pancreatic cancer cell lines. Cancer Res 2010; 70(22): 9494-9504. [ Links ]
67. Roth, A, Harle Calvin B, Baerlocher GM. Imetelstat (GRN163L)-telomerase-based cancer therapy. Small Mol. One 2010;184: 221-234. [ Links ]
68. Vonderheide RH, Schultze JL, Anderson KS, Maecker B, Butler MO, Xia Z et al. Equivalent Induction of Telomerase-specific Cytotoxic T Lymphocytes from Tumor-bearing Patients and Healthy Individuals. Cancer Res 2001; 61(23): 8366-8370. [ Links ]
69. Vonderheide RH, Domchek SM, Schultze JL, George DJ, Hoar KM, Chen D et al. Vaccination of Cancer Patients Against Telome-rase Induces Functional Antitumor CD8 + T Lymphocytes. Clin. Cancer Res 2004;10(3):828-839. [ Links ]
70. Parkhurst MR, Riley JP, Igarashi T, Li Y, Ro-bbins PF, Rosenberg SA. Immunization of Patients with the hTERT : 540-548 Peptide Induces Peptide-Reactive T Lymphocytes That Do Not Recognize Tumors Endoge-nously Expressing Telomerase Clin. Cancer Res 2004;10(14): 4688-4698. [ Links ]
71. Kyte JA, Gaudernack G, Dueland S, Trach-sel S, Julsrud L, Aamdal S. Telomerase pep-tide vaccination combined with temozolo-mide: a clinical trial in stage IV melanoma patients. Clin. Cancer Res 2011;17(13): 4568-4580. [ Links ]
72. Inderberg-Suso E-M, Trachsel S, Lislerud K, Rasmussen A-M, Gaudernack G. Widespread CD4+ T-cell reactivity to novel hTERT epitopes following vaccination of cancer patients with a single hTERT peptide GV1001. Oncoimmunology 2012; 1(5): 670-686. [ Links ]
73. Kammerer R, Zimmermann W. Coevolu-tion of activating and inhibitory receptors within mammalian carcinoembryonic antigen families. BMC Biol 2010; 8: 12-20. [ Links ]
74. Kuespert K, Pils S, Hauck CR. CEACAMs:their role in physiology and pathophysiolo-gy. Curr. Opin. Cell Biol 2006; 18(5): 565-571. [ Links ]
75. Diamandis EP, Bast RC, Gold P, Chu TM,Magnani JL. Reflection on the discovery of carcinoembryonic antigen, prostate-specific antigen, and cancer antigens CA125 and CA19-9. Clin. Chem 2013; 59(1): 22-31. [ Links ]
76. Kim KS, Kim J-T, Lee S-J, Kang MA, Choe IS, Kang YH et al. Overexpression and clinical significance of carcinoembryonic antigen-related cell adhesion molecule 6 in colorectal cancer. Clin. Chim. Acta 2013; 415: 12-19. [ Links ]
77. Blumenthal RD, Leon E, Hansen HJ, Goldenberg DM. Expression patterns of CEA-CAM5 and CEACAM6 in primary and metastatic cancers. BMC Cancer 2007; 7: 2-10. [ Links ]
78. Orava EW, Abdul-Wahid A, Huang EH-B,Mallick AI, Gariepy J. Blocking the attachment of cancer cells in vivo with DNA ap-tamers displaying anti-adhesive properties against the carcinoembryonic antigen. Mol. Oncol 2013; 7(4): 799-811. [ Links ]
79. Ullenhag GJ, Frödin J-E, Jeddi-Tehrani M, Strigärd K, Eriksson E, Samanci A et al. Durable carcinoembryonic antigen (CEA)-speci-fic humoral and cellular immune responses in colorectal carcinoma patients vaccinated with recombinant CEA and granulocyte/ macrophage colony-stimulating factor. Clin.Cancer Res 2004; 10(10): 3273-3281. [ Links ]
80. Marshall BJL, Hawkins MJ, Tsang KY, Richmond E, Pedicano JE, Zhu M et al. Phase I Study in Cancer Patients of a Replication Defective. J. Clin. Oncol 1999;17(1): 332-337. [ Links ]
81. Conry RM, Allen KO, Lee S, Moore SE,Shaw DR, Lobuglio AF. Human Autoanti-bodies to Carcinoembryonic Antigen (CEA) Induced by a Vaccinia-CEA Vaccine. Clin. Cancer Res 2000; 6(1): 34-41. [ Links ]
82. Zhu M, Marshall J, Cole D, Schlom J, Tsang KY. Specific Cytolytic T-Cell Responses to Human CEA from Patients Immunized with Recombinant Avipox-CEA Vaccine. Clin.Cancer Res 2000; 6(1): 24-33. [ Links ]
83. Moulton HM, Yoshihara PH, Mason DH,Iversen PL, Triozzi PL. Active Specific Immunotherapy with a ß -Human Chorionic Gonadotropin Peptide Vaccine in Patients with Metastatic Colorectal Cancer : Antibody Response Is Associated with Improved Survival. Clin Cancer Res 2002; 8: 2044-2051. [ Links ]
84. Jeon Insik, Iyer Karthik, Danishefsky Samuel J. A Practical Total Synthesis of Globo-H for Use in Anticancer Vaccines. J. Organic Chem 2009; 74(21): 8452-8455. [ Links ]
85. Oka Y, Tsuboi A, Taguchi T, Osaki T, Kyo T, Nakajima H et al. Induction of WT1 Wilms tumor gene -specific cytotoxic T lympho- 95. cytes by WT1 peptide vaccine and the resultant cancer regression. PNAS 2004; 101(38):13885-13890. [ Links ]
86. Maeda T, Hosen N, Fukushima K, Tsuboi A, Morimoto S, Matsui T et al. Maintenance of complete remission after allogeneic stem cell transplantation in leukemia patients treated with Wilms tumor 1 peptide vaccine. Blood Cancer J 2013; 3(8): 130-133. [ Links ]
87. Textor S, Fiegler N, Arnold A, Porgador A, Hofmann TG, Cerwenka A. Human NK cells are alerted to induction of p53 in cancer cells by upregulation of the NKG2D ligands ULBP1 and ULBP2. Cancer Res 2011; 71(18): 5998-6009. [ Links ]
88. Speetjens FM, Kuppen PJK, Welters MJP, Es-sahsah F, Voet van den Brink AMEG, Lan-trua MGK et al. Induction of p53-specific immunity by a p53 synthetic long peptide vaccine in patients treated for metastatic co-lorectal cancer. Clin. Cancer Res 2009; 15(3):1086-1095. [ Links ]
89. Yang F, Yang X-F. New concepts in tumor antigens: their significance in future immunotherapies for tumors. Cell. Mol. Immunol 2005; 2(5): 331-341. [ Links ]
90. Frontiers P, With S, Than M. 2013 Report:Medicines in Development - Biologics. Pharmaceutical Research and Manufacturers of America: Washington, D. C, 2013. Disponible en: http://phrma.org/sites/default/files/2488/biologics2013.pdf. [ Links ]
91. Palucka AK, Ueno H, Fay J, Banchereau J. Dendritic cells: a critical player in cancer therapy? J. Immunol 2009; 31(9): 793-805. [ Links ]
92. Clive KS, Tyler JA, Clifton GT, Holmes JP,Mittendorf E a, Ponniah S et al. Use of GM-CSF as an adjuvant with cancer vaccines: beneficial or detrimental? Vaccines 2010; 9(5):519-25. [ Links ]
93. Schreiber RD, Old LJ, Smyth MJ. Cancer Immunoediting : Integrating Suppression and Promotion. Science 2011; 331: 1565-70. [ Links ]
94. Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell 2010;140(6): 883-99. [ Links ]
95. Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev.Cancer 2012; 12(4): 252-564. [ Links ]
96. Lipson, EJ, Charles G. Drake. Ipilimumab: an anti-CTLA-4 antibody for metastatic melanoma. Clin. Cancer Res 2011; 17(22): 6958-6962. [ Links ]
