










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Cardiología
Print version ISSN 0120-5633
Rev. Colom. Cardiol. vol.16 no.1 Bogota Jan./Feb. 2009
(1) Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Farmacia. Grupo Unimol. Bogotá, DC., Colombia.
(2) Universidad de Cartagena, Facultad de Medicina.Cartagena, Colombia.
Correspondencia: Dr. Claudio Gómez. Profesor Asociado. Universidad Nacional de Colombia. Carrera 30 No. 45-03, Ciudad Universitaria, Edificio 450. Teléfono: (57-1) 316-5000. Fax: (57-1) 316-5060. Bogotá, D.C., Colombia. Correo electrónico: cjgomeza@unal.edu.co
Recibido: 18/12/2008. Aceptado: 10/03/2009.
La superfamilia de receptores de hormonas nucleares, es un amplio grupo de proteínas cuya función es actuar como factores de transcripción para modular, de manera positiva o negativa, la expresión de genes involucrados en procesos de diferenciación, crecimiento, reproducción y metabolismo. Dada su participación en procesos fisiológicos claves, las disfunciones asociadas con estos receptores tienen enormes implicaciones en enfermedades de elevada importancia en salud pública como la enfermedad cardiovascular, la diabetes mellitus tipo 2 y el cáncer, entre otras.
En este trabajo se revisan algunos aspectos de esta superfamilia de proteínas, incluyendo su estructura, relación con el metabolismo de lípidos e implicaciones cardiovasculares. El trabajo se enfoca en los receptores activados por el proliferador del peroxisoma (PPAR), aunque se da una breve mirada a los receptores X hepáticos (LXR).
Palabras clave: receptores nucleares, metabolismo de lípidos, PPAR, LXR.
Nuclear hormone receptors superfamily are a wide group of proteins which function is to act as transcription factors in order to modulate in a positive or negative way the expression of genes involved in differentiation processes, growth, reproduction and metabolism. Given its participation in key pathologic processes, the disfunctions associated to these receptors have huge implications in diseases of great importance in public health such as cardiovascular disease, diabetes mellitus type 2, and cancer between others.
Some aspects of this protein superfamily are reviewed in this study, including its structure, relationship with lipid metabolism and cardiovascular implications. This study focuses on the peroxisome proliferator-activated receptor (PPAR), and briefly on the liver X receptors (LXR).
Key Words: nuclear receptors, lipid metabolism, PPAR, LXR.
Introducción
La reciente finalización del proyecto Genoma Humano, que significó conocer con detalle la secuencia de nucleótidos de todo nuestro genoma, constituye una verdadera revolución en el campo de las ciencias biomédicas. En farmacología es un hito sin precedentes si se piensa en la cantidad de potenciales blancos farmacológicos, actuales y futuros, que se encuentran codificados en nuestro genoma (1).
Los receptores nucleares (NR) constituyen una familia de proteínas que funcionan como reguladores transcripcionales (positivos o negativos) de numerosos genes involucrados en importantes funciones fisiológicas, incluyendo control del desarrollo embrionario y diferenciación celular, y en mantener la homeostasis metabólica (2, 3). En consecuencia, cualquier disfunción de las vías reguladas por esta familia de receptores se asocia con enfermedades de tipo reproductivo, proliferativo y/o metabólico de enorme repercusión social, como la enfermedad cardiovascular, la diabetes, la hipertensión y el cáncer, por mencionar sólo algunas (3-8).
Ciertos miembros de NR reconocen uno o más ligandos naturales o sintéticos que se usan de manera terapéutica en el manejo de enfermedades de origen metabólico como lo es la diabetes mellitus tipo 2 y su compromiso cardiovascular (9-14).
Estructura
Todos los miembros de la superfamilia de receptores nucleares comparten una organización estructural común (Figura 1) que, para efectos de simplicidad, puede dividirse en tres regiones o dominios funcionales:
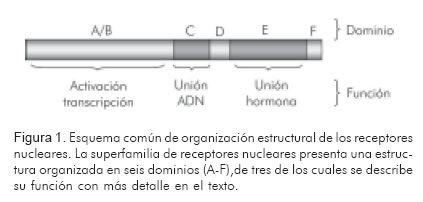
(i) una región N-terminal (dominio A/B), que varía en secuencia y longitud entre los diferentes miembros y contiene determinantes estructurales para la interacción con la maquinaria de transcripción basal y la regulación de su actividad.
(ii) una región muy conservada (dominio C, que también se conoce como DBD por las siglas del término en Inglés «Dominio de Unión a ADN»), implicada en la unión a ADN a nivel de los elementos genéticos específicos localizados en la región reguladora de cada gen diana.
(iii) región moderadamente conservada a la cual se fija el ligando (o fármaco) de cada receptor (domino de unión a ligando o LBD por sus siglas en Inglés) (15, 16).
Nomenclatura
En el genoma humano se han identificado 48 genes que codifican todos los miembros de la superfamilia de receptores nucleares conocidos a la fecha. Recientemente, un comité de expertos designados por la Unión Internacional de Farmacología Básica y Clínica (IUPHAR), propuso un sistema de nomenclatura basada en análisis filogenético de las secuencias génicas, específicamente en la evolución de los dominios conservados de unión a ligando y a ADN (16, 17). Este sistema divide la superfamilia en 6 subfamilias y 26 grupos de receptores (Tabla 1). Los receptores con estructura inusual, que contienen sólo uno de los dos dominios, se agrupan juntos como una subfamilia separada (llamada familia 0) independiente de su origen evolutivo. Cada gen se nombra con un prefijo «NR» seguido de un numeral arábigo que indica la subfamilia, una letra mayúscula que indica el grupo y otro numeral arábigo que se refiere a los genes individuales. Las isoformas alternativas del mismo gen se designan con una letra minúscula al final (16, 17). En la tabla 2 se muestra un ejemplo de la nomenclatura anterior, en este caso particular para miembros de la primera subfamilia, que incluye receptores de hormonas tiroideas, ácido retinoico y otros. Por el momento se acepta la terminología de la anterior nomenclatura, que usaba nombres triviales para los diferentes receptores, clasificados con base en la similitud de sus ligandos en tres grupos:
- Receptores de esteroides (ER), que incluye receptor de andrógenos (AR), receptor de progesterona (PR) y receptor de glucocorticoides (GR) entre otros.
- Receptores que forman heterodímeros con el receptor del ácido retinoico (RXR), incluyendo el receptor tiroideo (TR) y el receptor de vitamina D (VDR).
- Receptores huérfanos, llamados así porque no se había determinado aún sus ligandos fisiológicos encógenos o por mucho tiempo no fueron determinados.
Receptores activados por el proliferador del peroxisoma (PPAR)
Los receptores PPAR son factores de transcripción activados por ligando que pertenecen a la superfamilia de receptores nucleares y regulan la expresión de numerosos genes diana a través de su unión a elementos de secuencia específicos presentes en la región reguladora de tales genes. Se conocen tres isoformas de este receptor: PPARa (NR1C1), PPAR/d (NR1C2) y PPARg (NR1C3) (18).
PPARa está implicado en la oxidación de ácidos grasos y se expresa en mayor forma en tejidos metabólicamente activos como hígado, riñón y músculo esquelético, pero también se ha encontrado en corazón, monocitos, endotelio vascular y células de músculo liso (19-22). PPARg está involucrado en la diferenciación de células adiposas y en la sensibilidad a la insulina, y presenta dos isoformas: PPARg1 que se expresa a alto nivel en tejido adiposo, y PPARg2 de expresión en varios tejidos (18, 22). PPARg también se expresa en células musculares lisas del tejido vascular y en el corazón, si bien es muy escaso en este último. PPARb/a es el más extensamente distribuido en el organismo, y su nivel de expresión depende del grado de diferenciación (18).
Aunque se han identificado muchas moléculas activadoras de PPAR, sus ligandos fisiológicos endógenos todavía se desconocen; los ácidos grasos son uno de los ligandos naturales de PPARa, y activan genes del catabolismo de ácidos grasos; otros ligandos naturales son los eicosanoides (derivados del ácido araquidónico por las vías de la lipooxigenasa y la ciclooxigenasa). Los hipolipemiantes fibratos gemfibrozil y fenofibrato, son ligandos agonistas de PPARa que reducen la producción de triglicérido hepático por medio del aumento de la b-oxidación de ácidos grasos mediada por PPARa.
Los fármacos/moléculas activadores de PPARa (ácidos grasos, fibratos) y PPARg (tiazolidinedionas) tienen efectos antiproliferativos, antagonizan las acciones de la angiotensina II tanto in vivo como in vitro, y ejercen acciones antioxidantes al inhibir la generación de especies reactivas de oxígeno y la activación de mediadores inflamatorios en vasos sanguíneos y corazón. Estos agentes reducen la presión arterial en varios modelos de hipertensión, corrigen la disfunción endotelial y ejercen acciones anti-inflamatorias y antifibróticas en vasos y corazón (22-24).
Así, una serie de evidencias sugieren que los receptores PPAR cumplen una importante función vaso y cardioprotectora debido a su acción anti-inflamatoria y antioxidante a nivel de la pared vascular, mediada por un mecanismo de trans-represión de genes pro-inflamatorios (22-28). Todo este principio fisiológico permite mejorar el tratamiento y la evolución de las enfermedades cardiovasculares.
En tejido adiposo, PPARg es el principal regulador de la adipogénesis (diferenciación a adipocitos) tanto in vitro como in vivo (29). Evidencias in vivo en la que se usaron ratones knockout para PPAR, muestran que los heterocigotos PPARg+/- presentan disminución de tejido adiposo, y los ratones homocigotos faltos de ambos alelos (PPARg-/-) carecen completamente de tejido graso (30, 31). Adicionalmente, los ratones PPARg+/- presentan resistencia a ganar peso en una dieta alta en grasa (32). Así mismo, PPARg es importante para mantener el estado diferenciado funcional de las células adiposas (33).
PPARg regula de manera positiva la expresión de genes implicados en el metabolismo de lípidos en el adipocito, como aP2/FABP4, acil CoA sintetasa (34), proteína transportadora de ácido graso (35, 36) y lipoproteína lipasa (37), entre otros. Por otro lado, PPARg reprime la expresión de genes implicados en la lipólisis y la liberación de ácidos grasos tales como el receptor b3-adrenérgico (b3-AR) y de adipocitocinas como leptina y TNF-a (38-41). PPARg también regula efectivamente la expresión de genes implicados en la respuesta a la insulina, incluyendo el receptor de insulina y los sustratos del receptor de insulina (IRS-1, IRS-2) (42, 43), los cuales son necesarios para activar la captura celular de glucosa estimulada por insulina y mediada por el transportador de glucosa Glut4. Los fármacos de la familia de las tiazolidinedionas, estimulan la expresión y actividad de los transportadores de glucosa GLUT-4 en células de músculo liso (44, 45).
En cuanto al mecanismo de acción, los receptores PPAR se unen al ADN en la región reguladora de los genes sensibles, donde interactúan con secuencias de ADN específicas conocidas como elementos de respuesta al receptor del proliferador peroxisomal (PPRE). Por interacción directa con diferentes factores de transcripción y proteínas coactivadoras o correpresoras, los PPAR pueden inducir una respuesta positiva o negativa en la expresión de un gen determinado.
Receptores X hepáticos (LXR)
Se han identificado dos receptores tipo LXR, que se conocen como LXRa (NR1H3) y LXRb (NR1H2). LXRa es altamente expresado en hígado y en otros tejidos importantes para el metabolismo de lípidos (46). LXRb, por su parte, tiene una distribución menos específica y se halla en diferentes tejidos (47). Las secuencias de ambos receptores presentan 77% de identidad y son capaces de interactuar con el receptor X retinoide (RXR) para formar heterodímeros RXR/LXR los cuales son activados por unión del ligando a LXR pero no del ligando que se une a RXR (48). Los heterodímeros LXR/RXR, se enlazan a elementos de respuesta a LXR que contiene dos secuencias hexaméricas separadas por cuatro nucleótidos (47).
Los receptores LXR funcionarían como sensores de colesterol, respondiendo a ligandos naturales y sintéticos como los oxiesteroles, entre ellos 22(R)-, 24(S)-, 27-, y 24(S), 25-hidroxicolesterol (49). Por medio de ensayos con genes reporteros, se ha demostrado que los oxiesteroles se unen tanto a LXRa como a LXRb en rangos de concentración similares a los que se reportan para niveles fisiológicos, activando el heterodímero RXR/LXR (50). La hipótesis prevaleciente es que los LXR participan activamente en regular los mecanismos de conversión de colesterol en sales biliares (50, 51). Estudios con ratones knockout Lxra-/- demuestran que LXRa controla la expresión de la enzima 7a-hidroxilasa que incrementa la producción de ácidos biliares; dichos ratones muestran una disminución de LDL en plasma y demuestran también que el receptor LXRb no compensa la pérdida del receptor LXRa (52). Por otro lado, los ratones Lxrb-/- no tienen efecto sobre los niveles de ácidos biliares. Los ratones doble knockout Lxra-/- y Lxrb-/- muestran una fuerte variación en los niveles de expresión de proteínas implicadas en el manejo del colesterol a nivel hepático y una ingestión de colesterol disminuida (52), lo cual sugiere que los LXR regulan el metabolismo del colesterol a diferentes niveles.
Perspectivas
A pesar de los recientes avances, se requiere más investigación para comprender el papel exacto de algunos miembros de la superfamilia de receptores nucleares en la fisiopatología cardiovascular. El conocimiento de los genes implicados, sumado a las tecnologías actuales para generar animales transgénicos y knockout, nos acercará a responder tal interrogante, al tiempo que generará nuevos retos y desafíos en el campo de la farmacología molecular y clínica, como el enfoque hacia la búsqueda y el desarrollo de nuevos fármacos que mejoren las terapias actuales.
Agradecimientos
Esta revisión es parte de los requisitos que Adrián Sandoval debe cumplir para optar al grado de Magíster en Ciencias Farmacéuticas. Por ello expresa un sincero agradecimiento a todo el cuerpo docente del Departamento de Farmacia de la Universidad Nacional de Colombia por participar en su formación académica. Los autores Adrián Sandoval y Claudio Gómez-Alegría agradecen a la Universidad Nacional de Colombia - Dirección e Investigación Sede Bogotá (DIB) - por el apoyo a través de la financiación de los proyectos de investigación DIB Nº 8003056 y DIB Nº 8003115 que nos permite incursionar de forma experimental en este campo de investigación. Los autores Doris Gómez y Fernando Manzur Jattin, agradecen a la Universidad de Cartagena por facilitar el tiempo para este trabajo.
Bibliografía
1. Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, et al. Initial sequencing and analysis of the human genome. Nature 2001; 409 (6822): 860-921. [ Links ]
2. Gronemeyer H, Gustafsson JA, Laudet V. Principles for modulation of the nuclear receptor superfamily. Nat Rev Drug Discov 2004; 3 (11): 950-64. [ Links ]
3. Sonoda J, Pei L, Evans RM. Nuclear receptors: decoding metabolic disease. FEBS Lett 2008; 582 (1): 2-9. [ Links ]
4. Uauy R, Martinez JI, Rojas CV. Molecular nutrition, role of the PPAR system in lipidic metabolism and its importance in obesity and diabetes mellitus. Rev Med Chil 2000; 128 (4): 437-46. [ Links ]
5. Semple RK, Chatterjee VK, O’Rahilly S. PPAR gamma and human metabolic disease. J Clin Invest 2006; 116 (3): 581-9. [ Links ]
6. Conzen SD. Nuclear receptors and breast cancer. Mol Endocrinol 2008; 22 (10): 2215-28. [ Links ]
7. Bonta PI, Pols TW, de Vries CJ. NR4A nuclear receptors in atherosclerosis and vein-graft disease. Trends Cardiovasc Med 2007; 17 (3): 105-11. [ Links ]
8. Tenbaum S, Baniahmad A. Nuclear receptors: structure, function and involvement in disease. Int J Biochem Cell Biol 1997; 29 (12): 1325-41. [ Links ]
9. Staudinger JL. Liver-enriched nuclear receptors: therapeutic opportunities. Mol Pharm 2008; 5 (1): 1-2. [ Links ]
10. Sykiotis GP, Papavassiliou AG. Molecular mechanisms of transcriptional regulation by nuclear receptors. Perspectives for therapeutic implications. Hormones (Athens ) 2002; 1 (2): 69-75. [ Links ]
11. Smith AG, Muscat GE. Orphan nuclear receptors: therapeutic opportunities in skeletal muscle. Am J Physiol Cell Physiol 2006; 291 (2): C203-C217. [ Links ]
12. Zollner G, Marschall HU, Wagner M, Trauner M. Role of nuclear receptors in the adaptive response to bile acids and cholestasis: pathogenetic and therapeutic considerations. Mol Pharm 2006; 3 (3): 231-51. [ Links ]
13. Gong H, Xie W. Orphan nuclear receptors, PXR and LXR: new ligands and therapeutic potential. Expert Opin Ther Targets 2004; 8 (1): 49-54. [ Links ]
14. Sladek FM. Nuclear receptors as drug targets: new developments in coregulators, orphan receptors and major therapeutic areas. Expert Opin Ther Targets 2003; 7 (5): 679-84. [ Links ]
15. Robinson-Rechavi M, Escriva GH, Laudet V. The nuclear receptor superfamily. J Cell Sci 2003; 116 (Pt 4): 585-6. [ Links ]
16. Germain P, Staels B, Dacquet C, Spedding M, Laudet V. Overview of nomenclature of nuclear receptors. Pharmacol Rev 2006; 58 (4): 685-704. [ Links ]
17. A unified nomenclature system for the nuclear receptor superfamily. Cell 1999; 97 (2): 161-3. [ Links ]
18. Michalik L, Auwerx J, Berger JP, Chatterjee VK, Glass CK, Gonzalez FJ, et al. International Union of Pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacol Rev 2006; 58 (4): 726-41. [ Links ]
19. Staels B, Koenig W, Habib A, Merval R, Lebret M, Torra IP, et al. Activation of human aortic smooth-muscle cells is inhibited by PPARalpha but not by PPARgamma activators. Nature 1998; 393 (6687): 790-3. [ Links ]
20. Auboeuf D, Rieusset J, Fajas L, Vallier P, Frering V, Riou JP, et al. Tissue distribution and quantification of the expression of mRNAs of peroxisome proliferator-activated receptors and liver X receptor-alpha in humans: no alteration in adipose tissue of obese and NIDDM patients. Diabetes 1997; 46 (8): 1319-27. [ Links ]
21. Inoue I, Shino K, Noji S, Awata T, Katayama S. Expression of peroxisome proliferator-activated receptor alpha (PPAR alpha) in primary cultures of human vascular endothelial cells. Biochem Biophys Res Commun 1998; 246 (2): 370-4. [ Links ]
22. Schiffrin EL, Amiri F, Benkirane K, Iglarz M, Diep QN. Peroxisome proliferator-activated receptors: vascular and cardiac effects in hypertension. Hypertension 2003; 42 (4): 664-8. [ Links ]
23. Gerry JM, Pascual G. Narrowing in on cardiovascular disease: the atheroprotective role of peroxisome proliferator-activated receptor gamma. Trends Cardiovasc Med 2008; 18 (2): 39-44. [ Links ]
24. Genolet R, Wahli W, Michalik L. PPARs as drug targets to modulate inflammatory responses? Curr Drug Targets Inflamm Allergy 2004; 3 (4): 361-75. [ Links ]
25. Barish GD, Evans RM. PPARs and LXRs: atherosclerosis goes nuclear. Trends Endocrinol Metab 2004; 15 (4): 158-65. [ Links ]
26. Takase H, Nakazawa A, Yamashita S, Toriyama T, Sato K, Ueda R, et al. Pioglitazone produces rapid and persistent reduction of vascular inflammation in patients with hypertension and type 2 diabetes mellitus who are receiving angiotensin II receptor blockers. Metabolism 2007; 56 (4): 559-64. [ Links ]
27. Toba H, Miki S, Shimizu T, Yoshimura A, Inoue R, Sawai N, et al. The direct antioxidative and anti-inflammatory effects of peroxisome proliferator-activated receptors ligands are associated with the inhibition of angiotensin converting enzyme expression in streptozotocin-induced diabetic rat aorta. Eur J Pharmacol 2006; 549 (1-3): 124-32. [ Links ]
28. Tham DM, Martin-McNulty B, Wang YX, Wilson DW, Vergona R, Sullivan ME, et al. Angiotensin II is associated with activation of NF-kappaB-mediated genes and downregulation of PPARs. Physiol Genomics 2002; 11 (1): 21-30. [ Links ]
29. Rosen ED, Sarraf P, Troy AE, Bradwin G, Moore K, Milstone DS, et al. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol Cell 1999; 4 (4): 611-7. [ Links ]
30. Miles PD, Barak Y, He W, Evans RM, Olefsky JM. Improved insulin-sensitivity in mice heterozygous for PPAR-gamma deficiency. J Clin Invest 2000; 105 (3): 287-92. [ Links ]
31. Kubota N, Terauchi Y, Miki H, Tamemoto H, Yamauchi T, Komeda K, et al. PPAR gamma mediates high-fat diet-induced adipocyte hypertrophy and insulin resistance. Mol Cell 1999; 4 (4): 597-609. [ Links ]
32. Oliver WR, Jr., Shenk JL, Snaith MR, Russell CS, Plunket KD, Bodkin NL, et al. A selective peroxisome proliferator-activated receptor delta agonist promotes reverse cholesterol transport. Proc Natl Acad Sci U S A 2001; 98 (9): 5306-11. [ Links ]
33. Tamori Y, Masugi J, Nishino N, Kasuga M. Role of peroxisome proliferator-activated receptor-gamma in maintenance of the characteristics of mature 3T3-L1 adipocytes. Diabetes 2002; 51 (7): 2045-55. [ Links ]
34. Schoonjans K, Watanabe M, Suzuki H, Mahfoudi A, Krey G, Wahli W, et al. Induction of the acyl-coenzyme A synthetase gene by fibrates and fatty acids is mediated by a peroxisome proliferator response element in the C promoter. J Biol Chem 1995; 270 (33): 19269-76. [ Links ]
35. Frohnert BI, Hui TY, Bernlohr DA. Identification of a functional peroxisome proliferator-responsive element in the murine fatty acid transport protein gene. J Biol Chem 1999; 274 (7): 3970-7. [ Links ]
36. Martin G, Schoonjans K, Lefebvre AM, Staels B, Auwerx J. Coordinate regulation of the expression of the fatty acid transport protein and acyl-CoA synthetase genes by PPARalpha and PPARgamma activators. J Biol Chem 1997; 272 (45): 28210-7. [ Links ]
37. Schoonjans K, Peinado-Onsurbe J, Lefebvre AM, Heyman RA, Briggs M, Deeb S, et al. PPARalpha and PPARgamma activators direct a distinct tissue-specific transcriptional response via a PPRE in the lipoprotein lipase gene. EMBO J 1996; 15 (19): 5336-48. [ Links ]
38. Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM. Transcriptional regulation of adipogenesis. Genes Dev 2000; 14 (11): 1293-307. [ Links ]
39. Fajas L, Debril MB, Auwerx J. PPAR gamma: an essential role in metabolic control. Nutr Metab Cardiovasc Dis 2001; 11 (1): 64-9. [ Links ]
40. Kallen CB, Lazar MA. Antidiabetic thiazolidinediones inhibit leptin (ob) gene expression in 3T3-L1 adipocytes. Proc Natl Acad Sci U S A 1996; 93 (12): 5793-6. [ Links ]
41. Hofmann C, Lorenz K, Braithwaite SS, Colca JR, Palazuk BJ, Hotamisligil GS, et al. Altered gene expression for tumor necrosis factor-alpha and its receptors during drug and dietary modulation of insulin resistance. Endocrinology 1994; 134 (1): 264-70. [ Links ]
42. Iwata M, Haruta T, Usui I, Takata Y, Takano A, Uno T, et al. Pioglitazone ameliorates tumor necrosis factor-alpha-induced insulin resistance by a mechanism independent of adipogenic activity of peroxisome proliferator-activated receptor-gamma. Diabetes 2001; 50 (5): 1083-92. [ Links ]
43. Smith U, Gogg S, Johansson A, Olausson T, Rotter V, Svalstedt B. Thiazolidinediones (PPARgamma agonists) but not PPARalpha agonists increase IRS-2 gene expression in 3T3-L1 and human adipocytes. FASEB J 2001; 15 (1): 215-20. [ Links ]
44. Chai C, Chan WK. Developmental expression of a novel Ftz-F1 homologue, ff1b (NR5A4), in the zebrafish Danio rerio. Mech Dev 2000; 91 (1-2): 421-6. [ Links ]
45. Galarneau L, Drouin R, Belanger L. Assignment of the fetoprotein transcription factor gene (FTF) to human chromosome band 1q32.11 by in situ hybridization. Cytogenet Cell Genet 1998; 82 (3-4): 269-70. [ Links ]
46. Apfel R, Benbrook D, Lernhardt E, Ortiz MA, Salbert G, Pfahl M. A novel orphan receptor specific for a subset of thyroid hormone-responsive elements and its interaction with the retinoid/thyroid hormone receptor subfamily. Mol Cell Biol 1994; 14 (10): 7025-35. [ Links ]
47. Teboul M, Enmark E, Li Q, Wikstrom AC, Pelto-Huikko M, Gustafsson JA. OR-1, a member of the nuclear receptor superfamily that interacts with the 9-cis-retinoic acid receptor. Proc Natl Acad Sci U S A 1995; 92 (6): 2096-100. [ Links ]
48. Willy PJ, Mangelsdorf DJ. Unique requirements for retinoid-dependent transcriptional activation by the orphan receptor LXR. Genes Dev 1997 1; 11 (3): 289-98. [ Links ]
49. Beaven SW, Tontonoz P. Nuclear receptors in lipid metabolism: targeting the heart of dyslipidemia. Annu Rev Med 2006; 57: 313-29. [ Links ]
50. Janowski BA, Willy PJ, Devi TR, Falck JR, Mangelsdorf DJ. An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature 1996; 383 (6602): 728-31. [ Links ]
51. Lehmann JM, Kliewer SA, Moore LB, Smith-Oliver TA, Oliver BB, Su JL, et al. Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway. J Biol Chem 1997; 272 (6): 3137-40. [ Links ]
52. Peet DJ, Turley SD, Ma W, Janowski BA, Lobaccaro JM, Hammer RE, et al. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998; 93 (5): 693-704. [ Links ]

