










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista colombiana de Gastroenterología
Print version ISSN 0120-9957On-line version ISSN 2500-7440
Rev Col Gastroenterol vol.24 no.4 Bogotá Dec. 2009
Importancia de la proteína CagA en infección por Helicobacter pylori
The importance of CagA protein in Helicobacter Pylori infection
Azucena Arévalo, Bact. (1), Alba Alicia Trespalacios, MSc (2), William Otero, MD (3)
(1) Joven Investigador, Candidato a MSc, Facultad de Ciencias, Dpto Microbiología, Universidad Javeriana. Bogotá, Colombia.
(2) Profesora de Microbiología, Directora Especialización Microbiología Médica, Candidato PhD, Universidad Javeriana, Facultad de Ciencias, Dpto. de Microbiología. Bogotá, Colombia.
(3) Internista, Gastroenterólogo, Epidemiólogo, Profesor de Medicina, Unidad de Gastroenterología, Universidad Nacional de Colombia. Bogotá, Colombia.
Fecha recibido: 25-05-09 Fecha aceptado: 14-10-09
Resumen
Helicobacter pylori es una bacteria que tiene la capacidad de colonizar la mucosa gástrica de humanos y producir gastritis crónica y otras complicaciones. Está presente en 20-50% de la población en países industrializados y en el 80% o más en los países subdesarrollados. Es un microorganismo genéticamente variable, cuya mayor plasticidad genética se encuentra en un segmento de DNA de 40kb conocido como islote de patogenicidad (PAI), el cual codifica para la proteína CagA y para los componentes del sistema de secreción tipo IV, este último encargado de permitir la exportación de esta proteína al interior de la célula blanco. El gen cagA codifica la proteína CagA, cuya presencia es indicador de la isla de patogenicidad y de características patógenas de las cepas del microorganismo; con base en cag A, las cepas se clasifican como cagA+ y cagA- , siendo las primeras más virulentas que las segundas. La importancia de las cepas cagA+ es la evidencia de su relación con el desarrollo de cáncer gástrico. En la presente revisión se analiza el papel de los genes del islote de patogenicidad y su asociación con el desarrollo de patologías gastroduodenales.
Palabras clave
Helicobacter pylori, cagA, isla de patogenicidad.
Summary
Helicobacter pylori is a microorganism able to colonize gastric mucosa in humans where it can produce chronic gastritis and other type of complications. H. pylori is present approximately 20-50% in the industrialized countries but in developing countries its prevalence is the highest because approximately 80% of people are infected with the bacteria. In general this bacteria is variable in its genome but the greatest genetic plasticity is located at 40kb DNA segment, knowing as a pathogenicity island (PAI), inside of this DNA segment there are cagA gen which coding for CagA protein and genes that coding for type IV secretion system that is necessary for export CagA protein into target cell. cagA gen is important because it is a marker of PAI presence and because the presence of cagA has permitted classified H. pylori strains in cagA+ and cagA-, which is of great importance due cagA+ strains are more virulent than cagA- strains, although the principal importance of cagA + strains is its special association with gastric cancer. The aim of this review is study the functions of pathogenicity island genes and its association with gastro duodenal pathologies developing.
Key words
Helicobacter pylori, cagA, pathogenicity island.
Introducción
Helicobacter pylori (H. pylori) es una bacteria Gram negativa que coloniza el antro y/o el cuerpo del estómago humano, causando gastritis y complicaciones como úlcera gástrica, duodenal, linfomas MALT y adenocarcinoma gástrico (1-3). El desenlace de la enfermedad depende de factores ambientales, del huésped y de la bacteria (1-3). Dentro de las características atribuidas al microorganismo se han descrito varios factores de virulencia los cuales le permiten colonizar la mucosa gástrica, sobrevivir en el ambiente ácido del estómago y evadir la respuesta inmune (1-2).
A diferencia de otros patógenos entéricos como Salmonella o Yersinia que han desarrollado mecanismos para invadir las células M del epitelio intestinal, H. pylori permanece fundamentalmente en el moco gástrico, por fuera de la célula epitelial gástrica; sin embargo, gracias a varios de sus factores de virulencia es capaz de desencadenar señales en las células del huésped que interfieren con los procesos celulares básicos que finalmente culminan con la aparición de enfermedades (1, 4). Uno de los factores de virulencia más estudiados y codificados dentro del islote de patogenicidad es el gen cag A y su proteína Cag A (2-4). Las cepas de H. pylori capaces de codificar para esta proteína se han denominado cepas Cag A positivas; el gen se ha encontrado presente en aproximadamente el 90% de pacientes infectados con el microorganismo y su presencia se ha asociado estadísticamente con ulcera duodenal, atrofia de la mucosa gástrica y cáncer gástrico (5).
El objetivo de esta revisión es ampliar el panorama sobre la importancia del islote de patogenicidad asociado a cag en H. pylori, y analizar cómo algunos de los genes presentes en esta isla contribuyen al desarrollo de patologías gastroduodenales.
Características generales de las islas de patogenicidad
La evolución bacteriana no es un proceso continuo, por lo tanto, puede producirse por transferencia horizontal, es decir, adquiriendo segmentos de DNA de origen desconocido que se integran al cromosoma bacteriano mediante recombinación homóloga (4). Esta nueva porción de DNA integrado se conoce con el nombre de isla, cuyo DNA puede codificar para varias proteínas involucradas en sistemas de almacenamiento de hierro, enzimas metabólicas, sistemas de secreción, proteínas de superficie celular, factores de adherencia, toxinas etc., (4, 6). Las denominadas islas de patogenicidad PAIs se caracterizan por lo siguiente: tienen un contenido de guanina-citocina (G-C) diferente al resto del genoma, tienen un codón constante que se adapta al cromosoma bacteriano, están rodeadas de repeticiones directas (DR), se asocian a genes de RNA de transferencia (tRNA), tienen genes que codifican para factores móviles como integrasas, trasposasas y elementos de secuencias de inserción (IS) (6).
La importancia de las regiones DR, de genes de tRNA y de elementos IS es que actúan como sitios de deleción de las PAIs motivo por el cual estas son inestables (6). Sin embargo, estas regiones también pueden actuar como sitios de unión de DNA. En H. pylori, una de las secuencias que actúa como lugar de unión es el gen glr que codifica para la glutamato racemasa (6). Gracias a la presencia de estas regiones después de la adquisición inicial de DNA foráneo, las PAIs pueden optimizarse de acuerdo a las necesidades de la célula receptora (4).
Las islas de patogenicidad están reguladas por genes que codifican para factores reguladores localizados dentro de la misma isla que también están involucrados en la regulación de genes fuera de PAI; sin embargo, PAI también puede estar regulada por genes localizados fuera de ella y que a la vez regulan genes housekeeping (6).
Isla de patogenicidad de H. pyloriEl concepto de PAIs fue inicialmente desarrollado para describir la adquisición de segmentos de DNA de cepas de E. coli patógenas ausentes en las no patógenas. Cencini y col (7), en 1996, encontraron la presencia de PAI en H. pylori, cuyo hallazgo ha sido confirmado y extendido por estudios subsecuentes. PAI de H. pylori inicialmente se denominó cag (gen asociado a citotoxina) ya que se pensaba que estaba asociado con la expresión de VacA (vacuolotoxina); sin embargo, posteriormente se observó que los dos factores VacA y PAI son independientes (4, 8).
A nivel genético, acorde con análisis del genoma de H. pylori, PAI es considerada la principal zona variable en todo el genoma de la bacteria (9). Está constituida por un marco abierto de lectura (ORF) de 40 kb rodeada por 31pb de repeticiones directas, las cuales contienen sitios de recombinación que corresponden con los últimos nucleótidos del gen glr (glutamato racemasa) (4), es decir, que cagPAI se encuentra entre el gen glr y un ORF cuya función aún no se ha descrito, conocido como ORF5 y descrito por Tomb et al, en 1997 como HP0519 (10). Tanto a la izquierda como hacia el extremo derecho tiene secuencias de inserción, llamadas elementos IS605, las cuales varían en número dependiendo de la cepa. En razón a esto, cepas con muchos elementos de inserción se las ha designado cepas tipo II y se caracterizan por ser menos virulentas que las cepas tipo I que contienen a PAI (11). Adicionalmente, dos de estos elementos IS605 se encargan de dividir PAI en dos regiones, designadas como cag I y cag II (4, 7) (figura 1A).
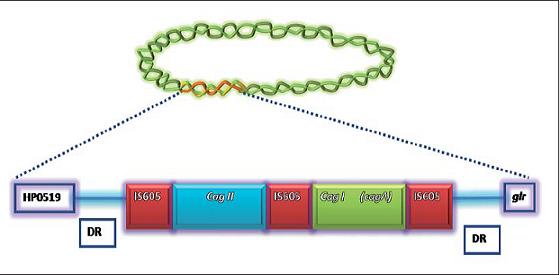
Figura 1a. Representa la isla de patogenicidad de H. pylori. Región que se encuentra entre el gen glr y un ORF HP0519 del cual no se conoce su función. PAI está rodeada por secuencias de repeticiones directas (DR), está dividida en cagI y cagII por elementos IS605 cuya cantidad pueden variar entre cepas. Dentro de la región cagI y cagII hay genes involucrados en codificar para el T4SS con la diferencia de que dentro de la región cagI se encuentra el gen cagA.
En H. pylori, cagPAI se puede encontrar como una sola unidad no interrumpida, dividida en las dos regiones ya mencionadas por la secuencia de inserción o como un fragmento largo de DNA parcialmente delecionado. Audibert C, y col (5), al evaluar la estructura de cag PAI encontraron cepas que no tenían interrupciones de cag PAI, cepas con cag PAI dividido en dos y cepas sin cag PAI, además, reportan el hallazgo de cepas que presentaban solo la región media (IS605) y cag II, así como también cepas con la región media y cag I, o cepas con solo la región cagII, cag I o IS605 únicamente (5).
Algunos de los genes en la isla codifican para proteínas similares a las involucradas tanto en la transferencia de DNA (Vir y Tra) y en la exportación de toxina (Ptl); sin embargo, estos genes no tienen operones conservados de estas proteínas (Vir, Tar, Ptl) como en otros microorganismos, lo que sugiere que PAI no se deriva directamente de estos sistemas. Dentro de las proteínas involucradas en transferencia se encuentra el sistema de secreción tipo IV (12). Acorde con la secuencia del genoma de la cepa 26695 y NCTC 11638 de H. pylori se ha reportado que 30 proteínas codificadas por los genes de PAI tienen secuencia de señal, las cuales tienen un tamaño aproximado de 25 aminoácidos, lo que sugiere que PAI es de origen Gram negativo (4). También codifica para ocho proteínas de membrana interna con al menos dos dominios membrana. El porcentaje de identidad en los nucleótidos de PAI es de aproximadamente 97% lo que significa que entre cepas este 3% restante quizás represente diferencias notables en funciones biológicas relevantes (12).
cag II y cag. Censini y col (7) designaron como cag II a un segmento de DNA de cepas cagA+ presente en PAI y cag I al segmento que contiene el gen cagA. Sin embargo, en un estudio realizado por NS Akopyants, et al (10) se encontró que porciones del segmento cag II hibridaban con segmentos de cepas cagA- indicando que a lado y lado de este segmento hay secuencias comunes para todas las cepas de H. pylori (10).
Estudios realizados por NS Akopyants y col (10) evidenciaron que cag II es un segmento de 19kb constituido por 15 marcos abiertos de lectura único en cepas cagA+, cuyo contenido de A+T es corresponde al 64%, mientras que cag I tiene un contenido que corresponde al 65% comparado con el 61% de A+T presente en todo el cromosoma del microorganismo. En cag II se han encontrado cuatro ORFs cuyos genes codifican para proteínas homologas a A. tumefaciens y B. pertusis necesarias para la liberación de toxina y transferencia de DNA (tabla 1). Adicionalmente, a la derecha de cag II hay dos ORFs, 21 y 22 que codifican para transposasas de elementos IS homólogas en otras bacterias y que corresponden a secuencias repetitivas ("RS2"), conocidos como IS605 (10).
Tabla 1. ORFs en H. pylori cuyos genes codifican para proteínas homólogas a A. tumefaciens y B. pertusis.

Sistema de secreción tipo IV codificado por PAI
Los microorganismos han desarrollado sistemas de comunicación entre ellos y su hábitat, como taxis, quórum sensing y de secreción (4). En las bacterias patógenas gram negativas se han descrito cuatro sistemas de secreción diferentes, lo cual no excluye que pueda haber más, mediante los cuales se excretan proteínas, enzimas, DNA o factores de virulencia (4). Sin embargo, dentro de estos cuatro sistemas de secreción solo se ha descrito a dos como capaces de transportar factores de virulencia, el sistema de secreción tipo III presente en algunas enterobacterias y el sistema de secreción tipo IV (T4SS) (4), pero aunque los dos sistemas son capaces de transportar factores de virulencia, su origen evolutivo es diferente ya que el primero se relaciona con el sistema flagelar, mientras que el segundo es probablemente originado del aparato de conjugación bacteriana motivo por el cual se permite la transferencia de DNA; en H. pylori permite no solo la transferencia horizontal de genes sino también la importación de peptidoglicano y de la proteína CagA al interior de la célula epitelial (4).
Los componentes del sistema de secreción tipo IV de H. pylori así como su función y localización son homólogos a los componentes de A. tumefaciens (identificados como VirB) (13). En H. pylori se han identificado cerca de 18 genes encargados de codificar para el sistema de secreción tipo IV, los cuales están dentro de PAI. Gracias a estudios de microscopia electrónica se ha logrado identificar que el T4SS es un organelo filamentoso localizado en un polo de la superficie bacteriana e inducido por contacto (13, 14). El modelo de organización del sistema de secreción tipo IV propone que sus proteínas se agrupan en: proteínas citoplasmáticas o de la membrana interna, proteínas que forman el complejo central o core localizadas en el periplasma y proteínas que hacen parte del pili o de la estructura superficial que se proyecta mas allá de la membrana externa (14).
Una vez el microorganismo entra en contacto con la célula epitelial gracias a adhesinas accesorias comienza el ensamblaje del T4SS, inicialmente con las proteínas que hacen parte de la membrana interna como CagE/HPO544 (VirB4) y HPO525 (VirB11) (15), las cuales poseen actividad ATPasa para permitir la translocación del sustrato, una proteína adicional, HPO524 homóloga a VirD4 es la encargada de entregar el sustrato a la maquinaria de secreción de T4SS (15).
Posteriormente, comienza el ensamblaje de las proteínas que conforman el complejo central o core del T4SS, dentro de las cuales se encuentran CagT/12 (HPO532) que se halla en la base del organelo y permite el ensamblaje del resto del organelo; y CagV/10, CagX/8 (HPO528) y CagY/7 (HPO527) homólogas a VirB7, VirB8, VirB9, y VirB10 respectivamente, que hacen parte del complejo central propiamente dicho (14, 16). Aunque estas proteínas han sido descritas como parte del complejo central, un estudio de interacción proteína-proteína realizado por Valerie J Busler, et al (14), en 2006, reveló que tres de estas proteínas, CagY/7, CagX/8, CagT/12 están asociadas con el pili que se forma entre H. pylori y la célula epitelial; además, proponen que otras proteínas Cag como CagM/16, CagI/19, CagG/21 y CagF/22 podrían representar componentes del complejo core de H. pylori específicos (14). Debido a que las proteínas que forman el core deben atravesar el peptidoglicano se han identificado proteínas adicionales como HPO523 homóloga a VirB1 que podría actuar como una transglycosilasa, encargada de lisar la capa de mureína de la pared bacteriana y así facilitar el ensamblaje de T4SS a través de la pared bacteriana, la cual también es necesaria para la maduración de CagY/7, CagT/12 y de HPO539, una posible chaperona de estas proteínas (16).
Finalmente, se ensamblan las proteínas que conforman el pili, dentro de las que su mayor componente es CagC homólogo a VirB2, la cual a la vez puede estar recubierta total o parcialmente por CagY y Cag L, esta última actúa como adhesina y permite la conexión entre T4SS y la célula blanco (15) (figura1b).
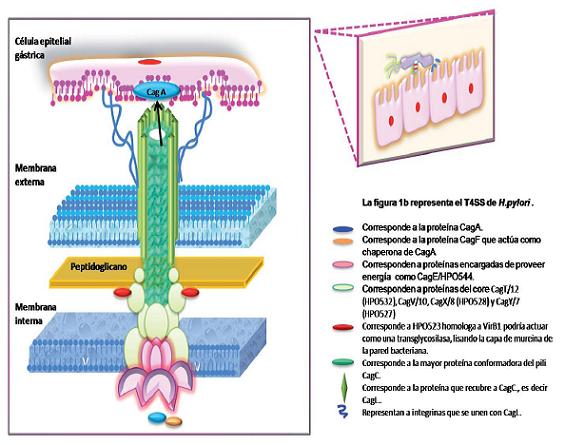
Figura 1b. Sistema de secreción tipo IV de H. pylori: En la figura representa la organización de las proteínas que conforman el T4SS del microorganismo, dentro de las que se encuentran proteínas con función ATPasa, las cuales se organizan en la membrana interna del la bacteria. Proteínas que hacen parte del complejo central y que se disponen a nivel del periplasma bacteriano, y finalmente atravesando parte del peptidoglino y la membrana externa se encuentran las proteínas que hacen parte del pili y que permitirán la comunicación con la célula epitelial a través de integrinas, para la posterior inyección de la proteína CagA.
Una vez se ha ensamblado el T4SS, para que se pueda transportar CagA a la célula epitelial se requiere que el sistema de secreción interactúe con un receptor específico en la célula huésped, por lo que se ha propuesto que las integrinas de la célula huésped podrían interactuar con la proteína CagL. Las integrinas son moléculas transmembrana de adhesión célula-célula y célula-matrix extracelular que se unen a motivos Arg-Gly-Asp (RGD); CagL contiene estos motivos por los cuales posiblemente suceda la interacción del pili a la integrina α5β1 para que posteriormente se inyecte la proteína CagA (15).
Transcurrido el ensamblaje del T4SS y la adhesión del sistema de secreción IV a la célula epitelial, se transloca la proteína CagA para lo cual se requiere de la proteína CagF que actúa como chaperona uniéndose a la porción C-terminal de la secuencia señal de la proteína CagA hasta que esta es translocada a la célula epitelial (15).
Cag A
Es el gen asociado a citotoxina, que codifica para la proteína CagA (2). Dentro de la región cag PAI hay 31 marcos abiertos de lectura (ORF), uno de estos ORF codifica para el antígeno inmunodominante CagA, que está localizado hacia la región 3´ final del islote de patogenicidad; CagA fue la primera proteína identificada de PAI y el mayor factor de virulencia de H. pylori, por lo que su presencia (cepas cagA +) es un marcador directo de PAI. La proteína fue descubierta a comienzos de 1990 de manera independiente por Cover, et al (15), Jean Crabtree, et al (15), y Covacci, et al (15), y desde entonces varios estudios han puesto en evidencia la importancia de esta proteína en la patogénesis del microorganismo (17). El tamaño molecular de cagA es variable (120-145 kDa) y depende del número de secuencias repetitivas localizadas en la región 3' del gen, es decir que a nivel proteico CagA sin repeticiones tienen un peso molecular de 128 kDa, mientras que cada repetición incrementa el peso molecular de la proteína en 4kDa, aceptándose un máximo de 4 repeticiones (4). En cepas occidentales se han encontrado dos tipos de repeticiones: una de 57 pb seguida de otra de 102 pb. En cepas asiáticas se ha encontrado la misma repetición inicial a las occidentales de 57pb, pero la segunda repetición es de 162pb (18). La proteína, gracias al sistema de secreción tipo IV es translocada ("inyectada") a la célula huésped y posteriormente fosforilada en sitios específicos conocidos como EPIYA y de esta manera interactúa con diversas vías de señalización, desencadenando cambios en el citoesqueleto, en la morfología y en la movilidad de la célula huésped (15, 19). Una vez CagA es translocada a la célula huésped en tirosin-fosforilada por las kinasas del huésped, la fosforilación ocurre en los sitios Glu-Pro-lle-Tyr-Ala también denominados motivos EPIYA los cuales se pueden repetir hasta cinco veces en la mitad del carbono terminal de CagA. Las repeticiones de estos motivos se encuentran rodeados por secuencias repetitivas de DNA involucradas en recombinación, basados en estas secuencias se han denominado cuatro sitios EPIYA (A, B, C, D) (15, 20). La presencia de estas repeticiones que rodean los motivos EPIYA son de gran importancia ya que podrían explicar la variabilidad en el número de estos motivos en CagA, así como diferencias en la patogenicidad de las cepas de H. pylori, ya que por ejemplo las cepas occidentales se caracterizan porque su proteína CagA está conformada por sitios EPIYA-A y B seguidos por una a tres repeticiones de 34 aminoácidos que contiene el sitio de EPIYA-C; mientras que las cepas de H. pylori asiáticas se caracterizan generalmente por expresar CagA con sitios de EPIYA-D que reemplazan a EPIYA-C (3, 15, 20). Los diferentes tipos de EPIYA determinan diferentes proteínas CagA y antropológicamente se podrían identificar el origen de los antepasados de una determinada población (18). El número de repeticiones, en especial las secundarias en los motivos de la proteína son de especial importancia en cáncer gástrico, cuya incidencia es significativamente más alta en pacientes infectados con cepas de H. pylori que presenten múltiples repeticiones; sin embargo, un inconveniente en la capacidad de estas cepas con múltiples repeticiones de contribuir al desarrollo de cáncer gástrico es su escasa supervivencia en pH ácido por lo que se ha sugerido que estas se manifiestan después de que los pacientes desarrollan atrofia e hipocloridria (3, 15). Sin embargo, esta posibilidad es aún teórica y se desconoce la forma en que se llevarían a cabo interacciones del microorganismo con la mucosa que conllevarían a condiciones de atrofia para permitir que se desarrollen variantes del microorganismo que empeoren esta condición (21). Una posibilidad podría ser la coinfección al inicio o durante el transcurso de la enfermedad con cepas de H. pylori con CagA diferentes.
La fosforilación es mediada por kinasas conocidas de la familia Src como SFKs cuya función en las células normales es controlar procesos en citoesqueleto, proliferación celular y diferenciación pero también se han descrito en procesos de carcinogénesis, otra kinasas que fosforilan a CagA pertenecen a la familia Abl como c-Abl y Arg (15). En la infección por H. pylori se ha descrito que actúan en forma y tiempo específico ya que Src es activada durante los estadios iniciales de la infección (0,5-2h), mientras que Abl es continuamente activada por el microorganismo (15). Una vez CagA es tirosin fosforilada por estas kinasas se une específicamente y activa tirosin fosfatasas SHP-2, una oncoproteína cuya mutación está asociada con procesos malignos en humanos. CagA desregula a SHP-2 perturbando a la Erk MAP quinasa así como desfosforilando FAK kinasas involucradas en adhesión focal induciendo cambios en la morfología celular, la cual adopta un fenotipo conocido como célula en forma de colibrí (15). CagA también daña la interacción célula-célula de manera independiente a la fosforilación. CagA destruye las uniones estrechas y causa pérdida de la polaridad en las células epiteliales, también desestabiliza el complejo E-caderina / β-catenina (3, 22). Existe la posibilidad de que esta interacción de CagA con desarreglos celulares contribuya con pasos claves a un motivo de estudio en el desarrollo de cáncer gástrico (22). Este mecanismo implica que H. pylori puede tener un efecto oncogénico directo sobre las células del epitelio gástrico por su oncoproteína CagA, y no solamente de manera indirecta produciendo inflamación persistente e hiperproliferación con el riesgo de que radicales libres lesionen el DNA de estas células con rápido crecimiento (22).
Naomi Ohnishi, et al (22) demostraron in vivo que CagA es una oncoproteína bacteriana cuya expresión es suficiente para desarrollar neoplasias. En su importante investigación utilizaron ratones transgénicos, que tuvieran capacidad para expresar la proteína CagA, que se pudiera o no fosforilar. La formación de tumores en los ratones estuvo asociada a la tirosin-fosforilación de CagA lo cual es esencial para que esta interactúe con la oncoproteína SHP-2 desregulándola, y permita la proliferación anormal de células epiteliales gástricas y altera la leucogénesis lo cual apoya la asociación de la infección por H. pylori y el desarrollo de linfomas de células B (22).
Sin embargo, la translocación de CagA y su posterior fosforilación, así como los efectos posteriores que causa en las vías de señalización y morfología celular pueden estar influenciados por factores adicionales a las kinasas. Chin Ho Lai, et al (23) demostraron que el colesterol es un factor importante para la acción de CagA ya que al haber una disminución de colesterol celular se reduce la translocación de CagA por parte de TSS4, la fosforilación, y se produce un bloqueo de la respuesta celular inducida por la proteína, incluyendo el fenotipo en forma de colibrí y la inducción de IL-8 (23).
cagA PAI en pacientes infectados con H. pyloriH. pylori coloniza la mucosa gástrica en aproximadamente el 70-90% de los individuos en países subdesarrollados, los cuales adquieren el microorganismo en edades tempranas pero la mayoría (80%) permanecen asintomáticos lo cual se puede deber en parte a la cepa que los infecta; sin embargo, individuos susceptibles es posible que desarrollen un grado variable de daños en la mucosa gástrica que van desde gastritis, úlceras, carcinomas gástricos y linfomas MALT (24).
La presencia de la isla de patogenicidad se ha estudiado usando como marcador el gen cagA y en varios de estos estudios a nivel mundial se ha reportado asociación entre la presencia de este factor de virulencia y la severidad de la enfermedad presentada por el paciente (11, 17,24). Hasan Umit, et al (24) encontraron asociación entre la presencia de cagA y el grado de inflamación en cuerpo y antro, así como la atrofia glandular en antro se ha asociado más con cepas cagA+ que cag A-. Aime T y otros autores (25) demostraron que las cepas cagA + son requeridas para la inducción de displasia y adenocarcinoma en la mucosa gástrica de gerbos mongolianos, así como la presencia de otros factores de virulencia como OipA el cual también ha sido encontrado en lesiones precancerosas en humanos (25). Nilsson, et al (26) reportaron que cagPAI sin deleciones se ha asociado con un aumento en el riesgo de desarrollar enfermedad severa, mientras que cepas con deleciones internas en cagPAI son menos virulentas. Mohaboob A, et al (11) tuvieron resultados concordantes con los hallazgos de Nilsson al encontrar asociación entre el daño histológico causado por cepas de H. pylori con cagPAI completa y con deleciones parciales en correlación con el estado de la enfermedad; observaron que cepas con deleciones parciales de cagPAI se encontraban en pacientes con úlcera duodenal y dispepsia funcional, mientras que en pacientes con úlcera gástrica y cáncer gástrico predominaron cepas con cagPAI completo, así como también la presencia de cagA sin deleciones predominó en esta población (11). En niños coreanos, se encontró que la estructura de cagPAI evaluada mediante la presencia de cagA, cagE y cagT, no presenta diferencias al encontrarse completa o parcialmente delecionada en relación a la severidad de la inflamación y la actividad de neutrófilos (27).
Además de la relación antes mencionada, se ha postulado la capacidad de cepas cagA positivas de inducir la producción de interleukina 8 (IL-8) gracias a su capacidad de activar MAP kinasas, las cuales regulan no solo diferenciación, proliferación, apoptosis, respuestas de estrés, sino también la respuesta inflamatoria (5, 28). Sin embargo, recientes estudios han replanteado esta posibilidad, al encontrar correlación entre la estructura de cag PAI y la inducción de IL-8, ya que al producir deleción en varios genes del islote de patogenicidad a excepción de cagA, se disminuye la inducción de la producción de esta interleukina, postulando así la presencia de genes diferentes dentro de cagPAI involucrados en la inducción de IL-8; como por ejemplo, proteínas externas de membrana como OipA (5, 30, 31). OipA es una proteína inflamatoria de membrana externa, su expresión está regulada por un mecanismo de reparación basado en el número de repeticiones citocinas-timinas (CT) en la región 5´ del gen oipA y dependiendo del estado del gen "prendido"-"apagado" puede verse afectada las características de virulencia del microorganismo; su expresión se ha asociado con úlcera duodenal, cáncer gástrico, con altas densidades de H. pylori, presencia de cepas cagA+, infiltración severa de neutrófilos, e inducción de IL-8 (21, 29-31). En un estudio, Yamaoka, et al (30) examinaron la capacidad de cuatro genes que codifican para proteínas de membrana externa (HPO638 (oipA), HPO796, HP1501 y babA2) para producir IL-8, y encontraron que cepas mutantes en HPO796, HP1501 y babA2 e incluso en cagE no tienen efecto significativo sobre la producción de IL-8, a diferencia de cepas mutantes en oipA y que en un 81% eran cagA+ en las que encontraron una reducción del 50% en la producción de IL-8, adicionalmente, que cepas cagA- pero con el gen oipA activo producen tres veces más IL-8 que cepas cagA- y oipA inactivo (30). Sin embargo, los mecanismos de inducción de IL-8 en la infección por H. pylori aún no son totalmente entendidos.
Conclusiones
Helicobacter pylori tiene la habilidad de inducir gastritis crónica, linfoma MALT y adenocarcinoma gástrico, por lo que ha sido clasificado como carcinógeno tipo I. Aunque los factores que determinan el desarrollo de la infección aún no son comprendidos, se ha sugerido que varios de sus factores de virulencia pueden estar involucrados en el desenlace de la enfermedad. Se ha planteado a cagA como un marcador de cag PAI, y si PAI está presente significa que el microorganismo tiene la capacidad de codificar para toda una maquinaria involucrada en transferencia horizontal y la exportación de proteínas, adicionalmente la presencia de cagPAI nos da un panorama de la amplia diversidad genética del microorganismo, al poseer puntos de recombinación y deleción de DNA foráneo confiriéndole ventajas a la bacteria, y permitirle adquirir factores de virulencia y por qué no, diseminación de la resistencia antibiótica.
Aunque en varios estudios se ha planteado la posible relación entre cepas cagA+ y el desenlace de la enfermedad, aún es materia investigación ya que el desenlace no solo depende exclusivamente del microorganismo, sino de varios factores adicionales, como es el ambiente, las características inmunes, genéticas, sociales y culturales del huésped; es importante tener presente que si dependiese solo del microorganismo, además de determinar cagA en una determinada población sería necesario determinar cuáles son las características presentes en los motivos de la proteína, así como también las de cagPAI específicamente para la población en estudio, y el análisis de la presencia de otros factores de virulencia del microorganismo independientes de cagPAI que pueden estar influenciando en la patogénesis de la bacteria. Dicho de otra manera, H. pylori con todos sus factores de virulencia es necesario pero no suficiente para producir la mayoría de los adenocarcinomas gástricos en las personas genéticamente predispuestas.
Referencias
1. Amieva Manuel R, El Omar E. Host Bacterial Interactions in Helicobacter pylori Infection. Gastroenterology 2008; 134: 306-329. [ Links ]
2. Michael Höcker, Peter Hohenberger. Helicobacter pylori virulence factorsone part of a big picture. Lancet 2003; 362: 1231-33. [ Links ]
3. Osamu Handa, Yuji Naito, Toshikazu Yoshikawa. Cag A protein of Helicobacter pylori: A hijacker of epithelial cell signaling. Biochemical pharmacology 2007; 73: 1697-1702. [ Links ]
4. Mobley Harry LT, Mendz George L, Hazell Stuart L. Helicobacter pylori. Physiology and Genetics. Chapter 31. The cagPathogenecy Island 2001. p. 608. [ Links ]
5. C Audibert, C Burucoa, B Janvier, JL Fauchére. Implication of the Structure of the Helicobacter pylori cag Pathogenicity Island in Induction of Interleukin-8 Secretion. Infection and Immunity 2000; 1625-1629. [ Links ]
6. Jôrg Harcker, James B. Kaper. Pathogenicity Islands and the Evolution of Microbes. Annual Review of Microbiology 2000; 54: 641-679. [ Links ]
7. Stefano Censini, Christina Lange, Zhaoying Xiang, Jean E Crabtree, Paolo Ghiara, Mark Borodovsky. cag, a pathogenicity island of Helicobacter pylori, encodes typeI-specific and disease-associated virulence factors. Proc Natl Acad Sci USA 1996; 93: 14648-14653. [ Links ]
8. Tummuru MK, Cover TL, Blaser MJ. Cloning and expression of a high-molecular-mass major antigen of helicobacter pylori: evidence of linkage to cytotoxin production. Infection and Immunity 1994; 61: 1799-1809. [ Links ]
9. Armelle Ménard, Antoine Danchin, Sandrine Dupouy, Francis Mégraud, Philippe Lehours. A Variable Gene in a Conserved Region of the Helicobacter pylori Genome: Isotopic Gene Replacement or Rapid Evolution? Dna Research 2008; 15: 163-8. [ Links ]
10. NS Akopyants, SW Clifton, DKersulyte, JECrabtree, BEYouree, CA Reece, et al. Analysis of the cag pathogenicity island of Helicobacter pylori. Molecular Microbiology1998; 28: 37-53. [ Links ]
11. Mohaboob A, Aleem A Khan, Santosh K Tiwari, Niyaz Ahmed, L Venkateswar Rao, CM Habibullah. Association between cag-pathogenicity island in Helicobacter pylori isolates from peptic ulcer, gastric carcinoma, and non-ulcer dyspepsia subjects with histological changes. Gastroenterol 2005; 11: 6815-6822. [ Links ]
12. Tomb, J.F, Owen White, Anthony R. Kerlavage, Rebecca A. Clayton, Granger G. Sutton, et al. The complete genome sequence of the gastric pathogen Helicobacter pylori. Nature. 1997; 388: 539-547. [ Links ]
13. Stefan Kutter, Renate Buhrdorf, Jûrgen Haas,Wulf Schneider-Brachert, Rainer Haas, Wolfgang Fischer. Protein Subassemblies of the Helicobacter pylori Cag Type IV Secretion System Revealed by Localization and Interaction Studies. Journal of Bacteriology 2008; 190: 2161-2171. [ Links ]
14. Valerie J Busler, Victor J Torres, Mark S McClain, Oscar Tirado, David B Friedman, Timothy L. Cover. Protein-Protein Interactions among Helicobacter pylori Cag Proteins. Journal of Bacteriology 2006;188: 4787-4800. [ Links ]
15. Steffen Backert, Matthias Selbach. Role of type IV secretion in Helicobacter pylori pathogenesis. Cellular Microbiology 2008; 10: 1573-1581. [ Links ]
16. Zhong Qiao, Shao Shi-He, Cui Lei-Lei, Mu Run-Hong, Ju Xiao-Li and Dong Su-Rong. Type IV secretion system in Helicobacter pylori: a new insight into pathogenicity. Clin Med J 2007; 120: 2138-2142. [ Links ]
17. Leen-Jan Van Doorn, Céu Figueiredo, Ricardo Sanna, Anton Plaisier, Peter Schneeberger, Wink de Boer, et al. Clinical Relevance of the cagA, vacA, and iceA Status of Helicobacter pylori. Gastroenterology 1998; 115: 58-56. [ Links ]
18. Yoshico Yamaoka, Mototsugu Kato, Masahiro Asaka. Geographic Differences in Gastric Cancer Incidence Can be explained by differences between Helicobacter pylori Strains. Internal Medicine 2008; 47: 1077-1083. [ Links ]
19. Yuri Churin, Laila Al-Ghoul, Oliver Kepp, Thomas F. Meyer, Walter Birchmeier, and Michael Naumann. Helicobacter pylori CagA protein targets the c.Met receptor and enhances the motogenic response. The Journal of Cell Biology 2003; 161: 249-255. [ Links ]
20. Masanori Hatakeyama. Oncogenic Mechanisms of the Helicobacter pylori CagA protein. Nature Reviews Cancer 2004; 4: 688-694. [ Links ]
21. Y Yamaoka, O Ojo, S Fujimoto, S Odenbreit, R Haas, O Gutierrez, H M T El-Zimaity. Helicobacter pylori outer membrane proteins and gastroduodenal disease. Gut 2006; 55: 775-781. [ Links ]
22. Naomi Ohnishi, Hitomi Yuasa, Shinya Tanaka, Hirofumi Sawa, Motohiro Miura, Atsushi Matsu, et al. Transgenic expression of Helicobacter pylori CagA induces gastrointestinal and hematopoietic neoplasms in mouse. PNAS 2008; 105: 1003-1008. [ Links ]
23. Chin Ho Lai, Yun-Chieh Chang, Shin-Yi Du, Hung-Jung Wang, Chun-Hsien Kuo, Shih-Hua Fang, Hua et al. Cholesterol Depletion Reduces Helicobacter pylori CagA translocation and CagA-Induced Responses in AGS Cells. Infection and Immunity 2008; 76: 3293-3303. [ Links ]
24. Hasan Umit, Ahmet Tezel, Sebnem Bukavaz, Gulbin Unsal, Muserref Otkun, Ali Riza Soylu, et al. The Relationship between Virulence Factors of Helicobacter pylori and Severity of Gastritis in Infected Patients.Dig Dis Sci 2009; 54: 103-110. [ Links ]
25. Aime T Franco, Elizabeth Johnston, Uma Krishna, Yoshio Yamaoka, Dawn A. Israel, Toni A. Nagy, et al. Regulation of Gastric Carcinogenesis by Helicobacter pylori Virulence Factors. Cancer Res 2008; 68: 379-387. [ Links ]
26. Nilsson Anna Sillén, Lena Eriksson, Mona-Lisa Strand, Helena Enroth, Staffan Normark, et al. Correlation between cag Pathogenicity Island Composition and Helicobacter pylori-Associated Gastroduodenal Disease. Infection and Immunity 2003; 71: 6573-6581. [ Links ]
27. Jae Sung Ko, Jeong Kee Seo. Cag Pathogenicity Island of Helicobacter pylori in Korean Children. Helicobacter 2002; 7: 232-236. [ Links ]
28. Sarah Keates, Andrew C Keates, Michel Warny, Richard M Peek, Jr., Paul G. Murray, Ciarán P. Differential Activation og mitogen-Activated Protein Kinanses in AGS Gastric Epithelial Cells by cag+ and cag- Helicobacter pylori. The Journal of Immunology 1999; 163: 5552-5559. [ Links ]
29. Takahiko Kudo, Zhannat Z. Nurgalieva, Margaret E. Conner, Sue Crawford, Stefan Odenbreit. Correlation between Helicobacter pylori OipA Protein Expression and oipA Gene Switch Status. Journal of Clinical Microbiology 2004; 42: 2279-2281. [ Links ]
30. Yoshio Yamaoka, Dong H. Kwon, and David Y. Graham. A Mr 34,000 proinflammatory outer membrane protein (oipA) of Helicobacter pylori. PNAS 2000; 97: 7533-7538. [ Links ]
31. Sicheng Wen, Dominique Velin, Christian P. Felley, Likun Du, Pierre Michetti, and Qiang Pan-Hammarstrôm. Expression of Helicobacter pylori Virulence Factors and Associated Expression Profiles of Inflammatory Genes in the Human Gastric Mucosa. Infection and Immunity 2007;75: 5118-5126. [ Links ]

