










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.23 n.1 Bogotá jan./jul. 2005
FITOMEJORAMIENTO, RECURSOS GENéTICOS Y BIOLOGíA MOLECULAR
Caracterización molecular y evaluación bioquímica de cultivares colombianos y germoplasma elite de maíz según contenido de aceite
Molecular characterization and biochemical evaluation of elite Colombian maize germplasm and cultivars regarding their oil content
María Isabel Peñaranda1 y Alejandro A. Navas2
1Investigadora, Programa Nacional de Recursos Genéticos y Biotecnología Vegetal, CORPOICA, C.I. Tibaitatá, Bogotá. e-mail: mipenarandar@unal.edu.coanavasa@corpoica.org.co2Coordinador de Investigación y Desarrollo Tecnológico, CORPOICA, C.I. La Libertad, Villavicencio. e-mail: anavasa@corpoica.org.coanavasa@corpoica.org.co
Recibido: 25 de marzo de 2004 Aprobado: 27 de mayo de 2005
Resumen
Existe evidencia que indica que algunos cultivares de maíz colombianos poseen contenidos de aceite similares a germoplasmas elite conocidos. Para conocer su uso potencial en las industrias del aceite y de los concentrados animales, y como base para programas de mejoramiento vegetal, es preciso analizar el germoplasma disponible mediante estudios genéticos y fenotípicos. A tal fin se evaluó la diversidad genética y el contenido de aceite de 22 genotipos de maíz que incluyeron cultivares criollos colombianos y germoplasma elite; la caracterización molecular se llevó a cabo con marcadores del tipo microsatélites (Simple Sequence Repeats, SSR) y análisis bioquímico. Los resultados de este estudio mostraron amplia variabilidad genética entre los genotipos, lo cual podría representar un potencial de uso en programas de mejoramiento, especialmente por las altas probabilidades de encontrar heterosis. Así mismo, la evaluación bioquímica permitió identificar aquellos materiales con los mayores contenidos de aceite, los cuales fueron evaluados adicionalmente para el perfil de ácidos grasos. Los resultados del estudio muestran la utilidad de los SSR para profundizar en el conocimiento y asignación de grupos heteróticos y revelar la relación de parentesco. Así mismo, aporta evidencia adicional a favor que la caracterización molecular permite la estimación de la variabilidad genética, la cual, combinada con caracteres fenotípicos de interés, puede convertirse en una herramienta útil que apoye el mejoramiento genético convencional.
Palabras clave adicionales: Microsatélites, variabilidad genética, mejoramiento, fingerprinting, heterosis, cultivares criollos.
Abstract: Evaluating germplasm by DNA fingerprinting or phenotyping represent necessary, commonly-used practices in improvement programmes. Molecular characterisation using microsatellite molecular markers, such as simple sequence repeats (SSR), and biochemical analysis allowed genetic diversity and oil content to be evaluated in 22 maize genotypes, including Colombian inbred germplasm and elite lines. The results presented broad genetic variability amongst genotypes, representing great potential for using the Colombian Maize germplasm bank in improvement programmes, especially in terms of generating a good hybrid. Biochemical evaluation allowed material having greater oil content to be identified and then evaluated regarding fatty acid profile. This study illustrated the use of SSR in gaining knowledge about heterotic groups, allocating them and revealing ancestral relationships among genotypes having known characteristics or pedigree. This further indicates that molecular characterisation allows genetic variability to be estimated and that it can be a useful tool for supporting conventional genetic improvement when combined with phenotypical characteristics of interest.
Additional key words: Microsatellite, genetic variability, breeding, fingerprinting, heterosis, native cultivars.
Introducción
EL MAÍZ es un cultivo económicamente importante para muchos países, lo cual ha llamado la atención de los fitomejoradores a nivel mundial. La producción global de maíz blanco y amarillo alcanza 605 millones de toneladas métricas por año (FAOSTAT, 2003). Su demanda a nivel mundial, está sujeta a los requerimientos de la industria de cereales, tanto para consumo humano como animal, bien como forraje o en concentrados y para cientos de propósitos industriales; presenta amplia distribución global, bajo precio en relación con otros cereales y posee propiedades biológicas e industriales (TEC, 2003). En Colombia, la producción en áreas de economía campesina se destina básicamente para consumo humano y la producción de cultivos mecanizados abastece de forma parcial la industria de concentrados; las importaciones abastecen los requerimientos manteniendo restringidos los cultivos nacionales y generando la salida de divisas; por lo cual se hace necesario aumentar la producción de manera competitiva tanto en calidad, como en cantidad.
El contenido de aceite es una variable agronómica de tipo cuantitativo de gran importancia por su uso potencial en la industria; el aceite proporciona mayor cantidad de calorías por unidad de peso que los carbohidratos. Investigaciones realizadas en cerdos y pollos demostraron la eficiencia en la alimentación animal del utilizar maíz con alto contenido de aceite, comparado con maíces normales (Thomison y Geyer, 2001). Otros estudios en alimentación de ganado bovino mostraron que su inclusión en dietas mejora la ganancia diaria de peso y reduce los costos adicionales de engorde, con respecto a dietas de maíz amarillo normal (Dekalb, 1997). Adicionalmente, el aceite de maíz es reconocido por su estabilidad y energía metabolizable, gracias a la distribución proporcional de sus ácidos grasos, especialmente los ácidos oleico y esteárico (Thomison et al., 2003).
El conocimiento del genoma es de gran utilidad para el mejoramiento de caracteres de tipo cuantitativos, tanto para estimar la variabilidad, como para efectos de mejora genética por cruzamiento o recombinación, puesto que estos parámetros dependen de la base genética de la población y de la detección de grupos heteróticos (Smith, 2000). La estimación de la similaridad genética (GS) parece ser efectiva para evaluar la diversidad genética entre genotipos de maíz, seleccionar materiales y hacer inferencias acerca de los mejores cruzamientos (Enoki et al., 2002).
Colombia cuenta con un gran potencial genético en maíz compuesto por la colección nacional (2.200 accesiones) y de materiales extranjeros (2.000 accesiones) cuya custodia reposa en el banco genético de CORPOICA; estos materiales son susceptibles de ser valorados y utilizados en programas de mejoramiento. Para hacer selección de cultivares en mejoramiento, es esencial tener una descripción exacta del germoplasma. Existen evidencias que indican que algunos materiales criollos colombianos poseen alto contenido de aceite. En el año 2000, Pioneer-Dupont publicó los resultados de un estudio en la base de datos Maize DB (2000) de Missouri, sobre el contenido de aceite en 9.000 accesiones del banco de germoplasma de Ames, Iowa (USA), empleando tecnología NIRS (Near Infrared Reflectance Spectroscopy) lo cual permitió la identificación de materiales colombianos con altos contenidos de aceite en grano.
Para la fase de caracterización molecular del germoplasma se han realizado estudios comparativos de marcadores en plantas que demostraron la eficiencia de los microsatélites (Simple Sequence Repeats, SSR) sobre otros marcadores (Powell et al., 1996), lo cual proporciona una poderosa metodología para discriminar aquellos genotipos con una significativa contribución de diversidad alélica (Djé et al., 2000). En este trabajo se realizó una aglomeración jerárquica mediante el método de Ward con el fin de disponer de una estimación del número de grupos e inferir sobre las relaciones de filogenia entre los materiales. Este método es apropiado porque incorpora el criterio de proximida, que consiste en buscar el par de grupos más próximos entre sí para formar un nuevo grupo en cada etapa, de forma que el incremento de la varianza intragrupal sea lo más pequeño posible. El método Ward (Procedimiento Jerárquico Aglomerativo) ha sido ampliamente probado en la caracterización de germoplasma exótico y mejorado de maíz por Franco et al. (1998, 1999) y por Franco y Crossa (2002). Sus resultados, que usaron la estrategia de la Ward-MLM, mostraron una clara distinción entre grupos y una predicción eficaz del número verdadero de subpoblaciones.
La valoración genética realizada en este estudio, que utilizó marcadores del tipo SSR, permitió estimar la variabilidad genética y la potencialidad de heterosis entre un grupo de variedades criollas y algunas líneas de maíz elite; adicionalmente, se estimó el contenido de aceite en ambos grupos. El reto hacia el futuro es gestionar recursos que permitan la continuación de este tipo de estudios.
Materiales y métodos
Material vegetal
En este estudio se evaluaron accesiones criollas colombianas de maíz con contenidos de aceite altos en grano, además de una variedad de grano comercial aceitero proveniente de un genotipo híbrido importado de Estados Unidos que se seleccionó como testigo y fue aumentado en campo, y del cual se obtuvo segregación fértil; éste se denominó "Importado USA". Así mismo, se usó germoplasma elite con contenido normal de aceite que fue seleccionado por sus características de interés tales como alto rendimiento, adaptación, calidad de grano y capacidad para generar buenos híbridos. Finalmente, se evaluaron dos genotipos cerosos (Waxy 100% amilopectina en grano) identificados por Sarmiento (2002, comunicación personal) y tres materiales (las líneas QPM-161 y QPM-165, y la línea 210) identificados por Pabón (2002, comunicación personal) (tabla 1).
Extracción del ADN
Muestras aleatorias de tejido vegetal (tres plantas por genotipo) fueron tomadas y liofilizadas siguiendo el protocolo de Hoisington et al. (1993). La extracción del ADN se realizó utilizando la metodología modificada de Saghai-Maroof et al. (199). El ADN fue cualificado en gel agarosa al 0,8%.
Amplificación del ADN
Se realizó un barrido molecular con un grupo de marcadores del tipo SSR reportados por Maize DB (2003). La selección de iniciadores se hizo teniendo en cuenta su distribución uniforme en el genoma. Se fabricó una mezcla balanceada o bulk de ADN, previa identificación del polimorfismo intravarietal (Ribaut et al., 1997). La amplificación de SSR permitió la obtención del patrón de bandas para establecer el fingerprinting. El procedimiento de PCR fue realizado en un termociclador programable (ERICOMP TwinBlockTM System Thermocycler); el volumen de reacción fue de 20 µl que contenían 10 ng de ADN genómico. Las condiciones de amplificación fueron las mismas a excepción de la temperatura, que osciló entre 50 y 60° C. El establecimiento de patrones de bandas fue revelado utilizando geles de agarosa al 3% (1,5% Metaphor - 1,5% Seakem) corrido a 100 voltios en TBE 5X (Tris/ácido bórico/EDTA), buffer (basado en el método de Hoisington et al., 1993); éstos fueron teñidos con bromuro de etidio para su visualización bajo luz ultravioleta.
Análisis estadístico de datos moleculares
El agrupamiento de los genotipos se basó en el Método de Mínimas Varianzas de Ward (SAS® V8e). El índice contenido de polimorfismo (PIC) para cada locus microsatélite fue determinado como lo describe Smith (2000), y es una medida de la diversidad alélica para un locus igual a 1-Sh2k , donde hk es la frecuencia de Kth alelo. Cuando se calcula de esta manera, PIC es sinónimo de diversidad genética (Enoki et al., 2002), donde el valor de uno '1', corresponde al máximo de variabilidad genética y cero '0' corresponde al monomorfismo total.
Análisis bioquímico
Determinación del contenido de aceite. Los genotipos fueron sembrados en una misma localidad (C.I. Turipaná), en la misma época y bajo condiciones de riego y fertilización similares a fin de aislar el efecto de la interacción genotipo × ambiente; ello permitió la generación de semilla fresca para realizar el análisis de aceite. Las semillas fueron tomadas a partir de la mezcla de los aumentos de semilla realizados en la primera siembra del 2002. La selección de la semilla fue un procedimiento al azar, puesto que algunos estudios sugieren que el contenido de aceite del grano procedente de la región media de la mazorca es más alto que el de granos provenientes de los extremos de la mazorca (Alexander y Lambert, 1968). Las muestras (5 g de semilla por accesión), con tres réplicas por accesión, fueron molidas, desecadas y pesadas para posterior extracción de grasa con éter de petróleo mediante la eliminación del solvente por evaporación. La grasa obtenida se pesó y recuperó para realizar posterior determinación del porcentaje de aceite (AOAC, 1986). Un genotipo adicional (CML 170) proveniente del Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT, 1996) fue incluido en este estudio.
Establecimiento del perfil de ácidos grasos. Se realizó cuantificación directa de los ácidos grasos palmítico, esteárico, oleico, linoleico y linolénico, mediante cromatografía de gases; ello requirió esterificación (derivatización pre-columna de la muestra). Para preparar los ésteres metílicos, se pesaron 50 mg del aceite por muestra, se adicionaron 0,5 mL de una solución 1N de KOH en metanol y se agitó vigorosamente. Se adicionó 0,7 mL de xileno y se dejó en reposo hasta la separación de las dos fases. De la fase oleosa se tomó una muestra (5 µL) para ser inyectada directamente en el cromatógrafo de gases. Se utilizó una columna SupelcoTM 2330 capilar 30 × 0, 25 mm × 0, 2 µm y como gas de arrastre N2 a 15 mL. min-1, con velocidades entre 0-20° C/min y temperaturas entre 150 y 240° C.
Resultados y discusión
Estimación de la diversidad genética
Fueron utilizados en este estudio marcadores moleculares del tipo microsatélite para caracterizar el germoplasma de maíz. Pero más allá de esto, se pretendió agruparlos en función de su similaridad genética con el propósito de estimar grupos heteróticos e inferir posibles loci asociados con contenidos y/o calidad de aceite. Una matriz de datos generada a partir del fingerprintig permitió identificar marcadores polimórficos entre cultivares criollos y germoplasma elite.
Polimorfismo dentro de genotipos
El polimorfismo intravarietal mostró que las variedades muestran un perfil molecular heterogéneo y heterocigoto. La variación fue detectada por identificación de diferentes bandas dentro de una misma variedad. La mezcla balanceada (bulk) de ADN incrementó la confiabilidad del estudio por el aumento de la probabilidad de identificar diferentes alelos.
Polimorfismo entre genotipos
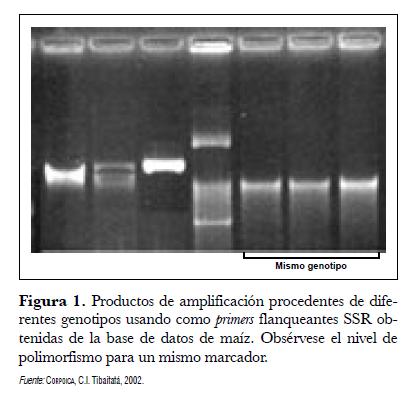
Se encontró alta divergencia en la frecuencia alélica entre los materiales (fig. 1). De los 90 primers SSR seleccionados, 4 resultaron monomórficos; se obtuvo un total de 391 bandas y el nivel de polimorfismos osciló entre 2 y 9 bandas por SSR, con un promedio 4,3 alelos por primer y un peso promedio de 145,7 pb. El análisis estimativo de los pesos de bandas fue determinado por interpolación entre las bandas de un marcador de peso conocido (25 Kb) y los productos de la amplificación. El procedimiento fue realizado utilizando el programa GeneTools® de Singene (Synophtics Ltda.).
El valor del índice de polimorfismo contenido (PIC) osciló en un rango de 0,0455 y 0,86, con un promedio de 0,7. El promedio de la medida del PIC en este estudio fue similar a la reportada por Enoki et al. (2002) (0,69) y más alta que la determinada por Smith et al. (1997) (0,62) y Senior et al. (1998) (0,59) en estudios similares. Los iniciadores más polimórficos se destacaron por tener en común un alto número en secuencia de repetición, con valores entre 16 y 29 'motifs'. Los resultados de la estimación de este índice se presentan en la tab. 2. El PIC reportado indica gran diversidad genética que es representada por genotipos elite y germoplasma local con características particulares y diferente origen.
En general, los materiales tendieron a agruparse en una escala de 0,01 a 0,1 de acuerdo con su centro de origen y/o características de interés (cultivares criollos aceiteros), germoplasma mejorado por el CIMMYT, genotipos Waxy, genotipos dulce y genotipos ICA-CORPOICA. El resultado más sobresaliente es una clara separación entre cultivares colombianos y los materiales elite, lo cual permitió inferir altas probabilidades de heterosis entre los dos grupos (fig. 2); el primer gran grupo fue conformado por genotipos elite, el cual se ramificó en cinco subgrupos de acuerdo con sus características y su genealogía. Los genotipos desarrollados por el Instituto Colombiano Agropecuario –ICA–, conformaron un solo cluster; igual sucedió con los materiales de CIMMYT y los genotipos Waxy.
Las razas criollas colombianas de maíz formaron un subgrupo con alta cercanía dentro del cual se encuentra el genotipo aceitero Importado USA el cual, a pesar de no ser un genotipo colombiano, parece compartir parte del background genético de los cultivares colombianos previamente seleccionados por contenido de aceite. Esto podría indicar que el recurso genético colombiano Cundinamarca 316 y el genotipo Importado USA, tienen carácteres y genes asociados con contenido de aceite en el grano procedentes de un mismo origen. Cabe destacar que la accesión Cundinamarca 316 exhibió la mayor cercanía con el genotipo Importado USA, aunque no necesariamente lo es específicamente para la característica 'contenido de aceite'.
En el análisis del patrón de bandas se encontró que 60% del total fueron comunes entre los cultivares criollos y el germoplasma elite, 3% fueron exclusivas para los genotipos Waxy y 6% exclusivas para los cultivares criollos. Finalmente, hubo 30% del total de las bandas exclusivas para los genotipos elite. Esta comparación fue realizada entre los cultivares criollos y el germoplasma elite a fin de identificar alelos específicos para cada grupo y comunes a ambos.La similaridad genética (GS) entre pares de accesiones (germoplasma elite y material criollo) fue alta. Esta correlación representa un estimativo de divergencia genética útil en el proceso de mejoramiento vegetal. La diferencia entre la similaridad genética por pares de genotipos contrastantes reveló que las líneas CML161 y CML165 mostraron la mayor divergencia con los cultivares criollos; ello permite inferir que el grado de mayor heterosis podría alcanzarse a partir de los híbridos obtenidos al cruzar estas líneas elite con el germoplasma criollo. El genotipo criollo Antioquia 442 presentó la mayor GS con ICAV 109 e ICAV 156, lo cual sugiere una baja posibilidad de heterosis; sin embargo, sólo la evaluación en campo podrá determinar la calidad de estos híbridos. Los cruzamientos elite × criollo con mayor posibilidad de heterosis fueron: CML 165 × Antioquia 442, Cundinamarca 316, Córdoba 342, Córdoba 338, Cariaco, y Dulce; y la línea elite CML 161 con el genotipo aumentado aceitero de USA y con Cundinamarca 316.
Análisis bioquímico
Los resultados oscilan en un rango entre 2,59 y 6,98 con un promedio de 4,398. La desviación estándar fue 1,00 y el coeficiente de variación fue 22,79%. Los genotipos donantes con mayor contenido de aceite fueron en su orden: el grano aumentado del genotipo comercial importado de USA, la accesión Antioquia 442 y la accesión Dulce, los cuales mostraron variación con relación a los valores obtenidos en el análisis preliminar.
Los genotipos con mayor contenido de aceite fueron en su orden: Importado USA (aumentado del grano comercial aceitero de USA), Dulce, la línea elite CML165 y el genotipo Antioquia 442 (tab. 3), los cuales mostraron variación con relación a los valores obtenidos en el análisis preliminar en el que el mejor grano aceitero fue Antioquia 442.
El genotipo Importado USA mostró un aumento de 6,12% a 6,98% en contenido de aceite. La segregación del Importado USA, aumentado en Colombia, exhibió niveles de aceite alrededor 2% y 3% más altos al ser comparado con los genotipos normales; estos resultados coinciden con los reportados por Thomison y Geyer (2001) al evaluar el grano comercial aceitero TCBlend®. El alto contenido de aceite en este genotipo puede deberse a que la mayoría de polinizaciones fueron efectivas para el macho aceitero y no para la línea androestéril. Antioquia 442 mostró diferencia de 1,06% entre la primera y segunda evaluación; esta variación puede atribuirse a que esta accesión fue aumentada en campo a 2.400 m.s.n.m. en la primera evaluación, en tanto fue cosechado a 28 m.s.n.m. en la segunda. El cultivar Dulce fue cosechado a 28 m.s.n.m para ambas evaluaciones y mostró una diferencia de 0,37%.
Los genotipos recurrentes CML 176 y CML 165 mostraron valores de 5% y 5,4% respectivamente, valores altos teniendo en cuenta que se trata de genotipos normales; ello indica que pueden dar origen a genotipos con alto contenido de aceite en menor tiempo. Por tanto, las posibilidades de heterosis entre estos genotipos y los tres mejores donantes aceiteros son altas. Respecto del contenido de aceite, se encontró variación entre los resultados obtenidos en los análisis realizados (años 2000 y 2001) empleando la misma metodología y el análisis final (año 2002) (tab. 4). Es posible que diversos factores tales como la magnitud de la influencia ambiental, la ocurrencia de efectos génicos (de los tipos aditivo y no aditivo) y la interacción genotipo-ambiente estén afectando la expresión de esta característica poligénica. Debe considerarse además, que los genotipos fueron sembrados en una sola localidad con una elevación de 28 m y las accesiones provienen de orígenes diferentes con elevaciones diferentes de hasta 2.200 m (tab. 1), lo cual sugiere que presiones climático-ambientales pudieron afectar la expresión de la característica bajo estudio.
Establecimiento del perfil de ácidos grasos
Este análisis fue realizado sólo para los genotipos con contenidos de aceite superiores a 5%. Los cultivares Waxy (100% amilopectina en grano) también fueron incluidos en este análisis. Los niveles de los ácidos palmítico y oleico mostraron los valores más bajos (tab. 5) en el genotipo importado de USA derivado del 'TCBlend®', mientras que mostraron el nivel más alto de ácido linoleico (51,8%); por su parte, el genotipo CML 176 mostró el mayor contenido de ácido linolénico (2,52%). Es preciso señalar que los ácidos grasos insaturados con más de una instauración (linoleico y linolénico) no pueden ser sintetizados por el organismo y deben ser suministrados en la dieta. Thomison et al. (2003) encontraron un nivel de ácido palmítico similar para cultivares normales y TCBlend® y niveles de ácido esteárico y oleico más altos. El contenido de ácido linolénico es concordante si se compara con los resultados de Thomison en 1999 (datos no mostrados) y los de este estudio; por supuesto, fue superior el contenido para los genotipos con contenidos normales de aceite. Los genotipos criollos Antioquia 442 y Dulce, se destacan por su alto contenido de ácido oleico (importante en la dieta alimenticia de monogástricos), a partir de lo cual se puede inferir que son apropiados por contenido y calidad de aceite. El genotipo Importado USA presentó los valores más bajos de ácido palmítico y el genotipo CML 156 mostró los valores más bajos de ácido esteárico, lo cual es positivo puesto que se trata de ácidos grasos saturados.
Discusión
Los datos SSR permiten estimar la diversidad genética entre y dentro las accesiones de muchas especies y pueden ser útiles para dilucidar estrategias de manejo de colecciones de germoplasma (Garland et al., 1999). El bulk de ADN permitió la identificación rápida de la diversidad alélica intra-cultivar para el caso de materiales heterogéneos-heterocigotos como las variedades criollas; ello incrementa la confiabilidad en la identificación de cultivares. Sin embargo, es necesario determinar el tamaño de la muestra para razas criollas, a fin de que se represente la variabilidad existente en la población. El número de marcadores utilizados permitió estimar la diversidad genética de la población. Enoki et al. (2002), reportaron que la precisión en la estimación de la similaridad genética aumenta en tanto se incrementa el número de loci que son evaluados. Por lo tanto, es presiso incurrir en altos costos y una labor intensiva de ADN-marcadores a fin de seleccionar los marcadores más informativos y estimar el mínimo número de marcadores que se requieren para obtener un buen nivel de precisión en la estimación de GS.
El contenido de aceite en maíz está sujeto a influencias de tipo genético y ambiental. Según Hellum et al. (1996), las prácticas agronómicas pueden influir en el contenido de aceite y la composición de los ácidos grasos; éstas diferencias pueden ser explicadas por efectos genéticos, ambientales y, de interacción genotipo-ambiente, que puede ser ejercido sobre caracteres de herencia poligénica.
El ambiente ejerce su efecto no sobre los genes sino sobre sus productos (expresión fenotípica del gen) lo cual puede verse reflejado en la heredabilidad de caracteres y, por tanto, en el desempeño agronómico de un cultivo. En condiciones inapropiadas o de estrés ambiental estos valores pueden ser bajos, situación que cambia en experiencias de campo bien planeadas; así, bajo excelentes condiciones es posible maximizar la expresión del carácter y hacer máxima la ganancia por selección de los mejores individuos (Kuruvadi et al., 1998).
Conclusiones
El uso de primers de tipo SSR permitió estimar la variabilidad y la organización del material genético entre cultivares. Todos los genotipos fueron asignados a clusters específicos. Se observó una clara asignación de los cultivares criollos y germoplasma elite a clusters diferentes; este nivel de disimilaridad dentro del pool de genes fue el esperado teniendo en cuenta el origen y características de los genotipos. Ello indica que el análisis mediante SSR proporciona suficiente precisión para estimar la variabilidad genética, lo cual muestra su utilidad en la caracterización y estimación de la base genética como base para estrategias de mejoramiento exitosas.
Se observó una amplia divergencia en el contenido de aceite en este estudio. Los cultivares criollos Antioquia 442 y Dulce fueron seleccionados como los mejores cultivares para la característica aceite, junto con el testigo Importado USA. Estos genotipos representan un precioso recurso para adelantar programas de mejoramiento destinadoa regiones del trópico bajo colombiano.
Los resultados obtenidos proporcionan un criterio útil para la exploración e introducción de genotipos, y para formar buenos híbridos con alto contenido de aceite, adaptados al trópico bajo colombiano. Los genotipos elite CML 161 y CML 165 mostraron el menor índice de similaridad genética con los cultivares criollos; así, estos materiales podrían emplearse como probadores dentro de un esquema de mejoramiento; los individuos resultantes de tales cruzas pueden exhibir buenas combinaciones alélicas (heterosis).
En este estudio se identificaron poblaciones base para desarrollar genotipos y/o líneas endocriadas con características especiales de industria y/o gran valor nutricional. La variabilidad de la muestra de trabajo representa un gran potencial de utilización en el desarrollo de programas de mejoramiento con base en diseños de cruzamientos y retrocruzamientos.
Agradecimientos
A Omar Cardona, Pilar Sarmiento y Edwin Pabón del Programa Nacional de Recursos Genéticos y Biotecnología Vegetal de CORPOICA (Corporación Colombiana de Investigación Agropecuaria); así mismo, agradecemos a las doctoras Beatriz Abadía y Olga Mayorga del Laboratorio de Nutrición Animal de CORPOICA en el C.I. Tibaitatá.
Literatura citada
Alexander, D.E. y R.J. Lambert. 1968. Relationship of kernel oil content to yield in maize. Crop Science 3, 354-355. [ Links ]
AOAC. 1986. Association Official Analytical Cchemists. Special properties of fats, oils, and shortening. Method 58-18. Washington D.C. [ Links ]
CIMMYT. 1996. Molecular marker applications to plant breeding. A workshop offered by the Applied Molecular Genetics Laboratory. El Batán, México. [ Links ]
Dekalb, N. 1997. TC Blend Seed for High Oil Opportunities. Business Bulletin DP97509. [ Links ]
Djé, Y; M. Heuertz; C. Lefebvre y X. Vekemans. 2000. Assessment of genetic diversity within and among germoplasma accessions in cultivated sorghum using microsatellite markers. Theoretical Applied Genetics 100, 918-925. [ Links ]
Enoki, H.; H. Sato y K. Koinuma. 2002. SSR analysis of genetic diversity among maize inbred lines adapted to cold regions of Japan. Theoretical Applied Genetics 104, 1270-1277. [ Links ]
FAOSTAT. 2003. Bases de datos de producción mundial y comercio internacional de maíz. FAO, Roma. [ Links ]
Franco, J. y J. Crossa. 2002. The modified location model for classifying genetic resources. Crop Science 42, 1719-1726. [ Links ]
Franco, J.; J. Crossa; J. Ribaut y J. Beltrán. 2001. A method combining molecular markers and phenotypic attributes for classifying plant genotypes. Theoretical Applied Genetics 103, 944-952. [ Links ]
Franco, J.; J. Crossa; J. Villaseñor; A. Castillo; S. Taba y S.A. Eberhart. 1999. A two-stage, three-way method for classifying genetic resources in multiple environments. Crop Science 39(1), 259-267. [ Links ]
Franco, J.; J. Crossa; J. Villaseñor; S. Taba y S.A. Eberhart. 1998. Classifying genetic resources by categorical and continuous variables. Crop Science 38(6), 1688-1696. [ Links ]
Garland, S.; M. Abedinia y R. Henry. 1999. The use of microsatellite polymorphisms identification of Australian breeding lines of rice (Oriza sativa L.). Netherlands 108, 53-63. [ Links ]
Hoisington, D.; M. Khairallah y D. González de León. 1993. Protocolos de Laboratorio. Laboratorio de genética molecular aplicada del CIMMYT. México D.F. [ Links ]
Hellum, J.M.; D. Helm; V.L. Fergason y M.S. Zuber. 1996. Fatty acid composition of oil from normal and high-amilosa strain of corn (Zea mays L.). Crop Science 9, 746-749. [ Links ]
Kuruvadi, S.; H. Navarro; A. Paccapelo y H. De León. 1998. Aptitud combinatoria y heterosis entre cruzas dialélicas de maíz (Zea mays L.) bajo riego y temporal. Simposio sobre desarrollo de maíz tolerante a sequía y bajo nitrógeno. Poster 3-17. Maize DB. 2003. Disponible en: http://agron.missouri.edu.html. [ Links ]
Navas, A. 2000. Molecular studies for linkage analysis and determination of qualitative trait loci (QTL) for acid soil tolerance in maize (Zea mays L.). Tesis doctoral. Iowa State University, Ames, Iowa. 64 p. [ Links ]
Powell, W.; M. Morgante y M. Hanafey. 1996. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germoplasma analysis. Molecular Breeding 2, 225-238. [ Links ]
Ribaut, J.; X. Hu; D. Hoisington y D. González. 1997. Use of STSs, a rapid and reliable preselection tools in a marker assisted selection-backcross scheme. Plant Molecular Biology 15, 154-162. [ Links ]
Saghai Maroof, M.A.; G.P. Yang; R.M. Biyashev; P.J. Maughan y Q. Zhang. 1996. Analysis of the barley and rice genomes by comparative RFLP linkage mapping. Theoretical Applied Genetics 92, 541-551. [ Links ]
SAS Institute Inc. 1990. SAS/ STAT® User'Guide. Vol 2. GLM-VARCOMP Version 6. Fourth Edition. [ Links ]
Senior, M.; M. Murohy; M. Goodman y C. Stuber. 1998. Utilizaty of SSRs for determining genetic similarities and relationships in maize using an agarosa gel system. Crop Science 38, 1088-1098. [ Links ]
Smith, J. 2000. Genetics diversity among elite sorghum inbreds lines assessed whit simple sequences sepeats (SSR). Crop Science 49, 226-232. [ Links ]
Smith, J.S.C.; F.L. Chin; H.Shu; O.F. Smith; F.J. Wall; M.L. Semior; F.E. Mitchell; F. Kresobich y Ziegle, J. 1997. An evaluation of the utility of SSR loci as molecular markers in maize (Zea mays L.): Comparation with date from RFLP and pedigree. Theoretical Applied Genetics 95, 163-173. [ Links ]
TEC. 2003. Instituto Tecnológico de Monterrey. En: http://www.qro.itesm.mx/agronomia2/extensivos/Maíz. [ Links ]
Thomison, P. y A. Geyer. 2001. High Oil TC Blend® Performance Test. Ohio State University Extension. Ractsheet. Horticulture and Crop Science. Columbus, Ohio. [ Links ]
Thomison, P.; A. Geyer; I. Lotz; H. Siegrist y T. Dobbels. 2003. TopCross high oil corn production: select grain quality attributes. Agronomy Journal 95, 147-154. [ Links ]
Thomison, P.; A. Seller; I. Lotz; H. Siegrist y T. Dobbels. 2002. TopCross high oil corn production: agronomic performance. Agronomy Journal 94, 2901-299. [ Links ]

