










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.23 no.1 Bogotá Jan./July 2005
Evaluacion del potencial de rendimiento y de la resistencia a Phytophthora infestans (Mont. de Bary) en la colección de papas redondas amarillas de la especie Solanum phureja (Juz. et Buk.)
Evaluating potential yield and resistance to Phytophthora infestans (Mont. de Bary) in the Solanum phureja (Juz. et Buk.) yellow potato collection
Rocío Escallón1Mónika Ramírez1y Carlos E. ñústez2
1Ingeniera Agrónoma, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.2Profesor Asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. e-mail: cenuztezl@unal.edu.co
Fecha de recepción: 19 de noviembre de 2004 Aceptado para publicación: 27 de mayo de 2005
Resumen
En el Centro Experimental ICA San Jorge (Soacha, Cundinamarca) a 2.900 m.s.n.m. y en el Centro Agropecuario Marengo (Mosquera, Cundinamarca) a 2.547 m.s.n.m. se realizó la evaluación de 25 genotipos de papa con tubérculos redondos amarillos de la especie S. phureja, provenientes de una colecta realizada por el convenio Fenalce-Fedepapa. Se utilizó un diseño de bloques completos al azar y las variables evaluadas fueron el peso de los tubérculos distribuidos en tres categorías: primera, segunda y tercera. Los resultados permitieron identificar un grupo de genotipos con promedios de producción comercial (tubérculos de primera y segunda) superiores al cultivar tradicional "yema de huevo". La evaluación de la resistencia frente al patógeno P. infestans, empleando la metodología de inoculación sobre foliolos en cámara húmeda de las razas cero y compleja H1N, permitió concluir que no existen genes mayores 'R' y en consecuencia son fuente de resistencia horizontal.
Palabras clave adicionales: Papa diploide, papa nativa, recursos genéticos, tizón tardío.
Summary
Twenty-five Solanum phureja yellow potato (round tuber) genotypes were evaluated at ICA's San Jorge Experimental Station in Soacha (Cundinamarca department, 2,900 m a.s.l.) and at the Universidad Nacional's Marengo Experimental Station in Mosquera (Cundinamarca department, 2,550 m a.s.l.); the tubers were obtained from the Fenalce-Fedepapa project potato collection. A random complete block design was employed for measuring tuber size and weight based on first, second and third class. The results led to a group of genotypes being identified having average commercial production size (first and second class tubers) superior to the traditional "egg yolk" variety. Inoculating leaves in a damp chamber using Zero biotypes and the HIN complex for evaluating resistance to P. infestans led to the conclusion that there are no R genes in these genotypes, meaning that they are thus a source of horizontal resistance.
Additional key words: Diploid potato, native potato, genetic resources, late blight.
Introducción
Solanum phureja, CONOCIDA COMO "PAPA CRIOLLA", es una especie de papa cultivada diploide (2n = 2X = 24) (Hawkes, 1990) que es cultivada en Colombia, parte de Ecuador y algunas zonas bolivianas y centroamericanas. Las grandes zonas productoras en Colombia están ubicadas en los departamentos de Cundinamarca, Boyacá, Nariño y Antioquia, algunas zonas menores se encuentran en los departamentos de Santander y Norte de Santander (Del Valle, 1994).
Algunas de las características mas prominentes de la especie son: periodo vegetativo corto, carencia de periodo de reposo del tubérculo y alto valor nutritivo, el cual es superior a variedades de papa de la especie S. tuberosum (Barrera, 1994). Esta especie es muy susceptible a heladas, y se cultiva comercialmente en regiones comprendidas entre 2.300 y 3.000 m.s.n.m., con un óptimo de producción a los 2.600 m.s.n.m. (Fedepapa, 1988).
La producción y la calidad de la papa es el resultado de la interacción de la planta con factores agroecológicos (suelo y clima) durante el periodo vegetativo del cultivo (Lujan, 1994). El funcionamiento de la planta está influenciado por la temperatura, longitud del día, intensidad de la luz, densidad de siembra, estado sanitario y fisiológico de la semilla y la disponibilidad de agua y nutrientes, sin embargo, es difícil delimitar la influencia de cada factor separadamente, puesto que, además de su propia influencia, existe una interacción entre todos (López, 1977).
El convenio Fedepapa – Fenalce realizó una colección de papas redondas amarillas de la especie Solanum phureja durante los años 1992-1993, como parte de un trabajo que pretendía buscar los genotipos mas indicados para un proyecto de agroindustria, con el cual se buscaba realizar reconversión de áreas de producción de cebada en los departamentos de Cundinamarca y Boyacá. Luego de un trabajo de caracterización morfológica realizado junto con el ICA y la Universidad Nacional se dejaron 25 clones, de los cuales fue entregada una copia al programa de mejoramiento genético de papa de la Facultad de Agronomía de la Universidad Nacional para posteriores evaluaciones y mantenimiento (Argüello, 2004).
La evaluación de caracteres de importancia agronómica en materiales de colección, es una actividad necesaria, que redunda en el conocimiento de lo que se tiene en ellas, facilitando de esta forma la toma de decisiones para integrar nuevos genotipos en los programas de mejoramiento de las especies cultivadas.
En el cultivo de la papa, tanto diploide como tetraploide, la enfermedad conocida como 'gota' producida por Phytophthora infestans puede ocasionar pérdidas totales del cultivo; el control, de las epidemias producidas por este patógeno se ha realizado mediante el uso de fungicidas influyendo directamente sobre los costos de producción y deterioro ambiental, debido a que el cultivo de la papa se localiza en zonas ecológicas altamente favorables para el desarrollo de la enfermedad (Malamud et al., 1994). Una herramienta para el manejo de la enfermedad es el uso de materiales resistentes, los cuales están determinados por características genéticas. Esta resistencia puede ser de tipo hipersensitivo dado por genes mayores, los cuales se relacionan gen a gen con el patógeno, y resistencia parcial la cual se basa en muchos genes con efecto aditivo y herencia cuantitativa (Black 1970, citado por Chacón, 1995).
Carlisle et al. (2002) evaluaron la agresividad de 20 aislamientos de P. infestans de Irlanda del Norte utilizando foliolos de tres cultivares de papa con diferentes niveles de resistencia no específica, y encontraron variación significativa para los parámetros: periodo de latencia, frecuencia de infección, área bajo la curva de infección y capacidad de esporulación, demostrando la variación en la severidad sobre foliolos de la población de P. infestans evaluada, indicando la importancia de seleccionar apropiadamente los aislamientos agresivos para la evaluación de resistencia del hospedero al tizón tardío dentro de los programas de mejoramiento. Zimnoch-Guzowska et al. (2003), evalúo en 16 clones de hibridación somática [Solanum nigrum L. + ZEL-1136 (diploide)], sus parentales y tres cultivares de papa estándares, la resistencia a P. infestans, utilizando plantas completas y foliolos, con dos aislamientos altamente agresivos (US8 y MP322). En el análisis de foliolos se observó la ruptura de la resistencia del parental S. nigrum y la respuesta variable de los clones híbridos individuales. Nueve híbridos mostraron resistencia significativamente más alta que la de S. nigrum, y seis clones expresaron resistencia similar a este parental. Flier et al. (2003) evaluaron la respuesta en resistencia de varios cultivares de papa a cepas agresivas de P. infestans en experimentos de campo y laboratorio. El análisis de varianza reveló interacciones cultivar x aislamiento para la resistencia foliar y de tubérculo. No encontraron correlación entre los análisis de campo y laboratorio para la resistencia del tubérculo al tizón. En condiciones de campo encontraron asociación entre la infección de tizón tardío en follaje y la incidencia de tizón en tubérculo.
Sieczka et al. (2002) evaluaron la resistencia a P. infestans de 19 materiales de papa (entre los que se encontraban cultivares y líneas de mejoramiento) en campo y laboratorio, en este último se realizaron análisis en foliolos separados y cortes transversales de tubérculos inoculados con un aislamiento altamente patogénico (MP – 324). El área bajo la curva de progreso de la enfermedad fue calculada, y se encontraron diferencias significativas entre materiales. En los análisis de laboratorio se encontraron materiales de papa resistentes a P. infestans, sin embargo no coincidieron con los materiales que resultaron más resistentes en las evaluaciones de campo.
Con el propósito de documentar y ampliar el uso de la colección de papas redondas amarillas de S. phureja se realizó el presente trabajo, cuyos objetivos fueron, determinar el potencial de rendimiento de los materiales en dos localidades, y evaluar la resistencia a P. infestans en las accesiones a nivel de laboratorio.
Materiales y métodos
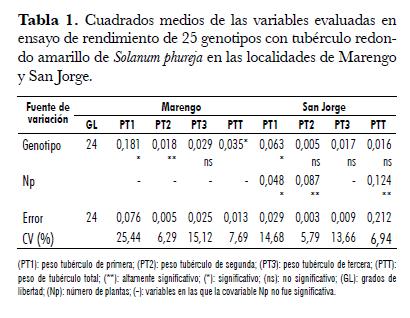
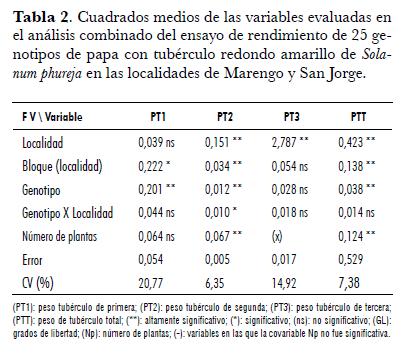
Se evaluaron 25 genotipos de la colección de papas redondas amarillas de la especie Solanum phureja. La evaluación se realizó en el Centro ICA San Jorge (Soacha, Cundinamarca) a 2.900 m.s.n.m. y en el Centro Agropecuario Marengo (Mosquera, Cundinamarca) a 2.547 m.s.n.m. Para le evaluación del potencial de rendimiento se utilizó el diseño de bloques completos al azar con dos repeticiones y 25 tratamientos (entradas de la colección). La unidad experimental estuvo conformada por tres surcos de 5 m cada uno, con una densidad de siembra de 33.300 sitios/ha. La variable evaluada fue peso de tubérculo para tres categorías por tamaño: primera (diámetro > 4 cm), segunda (diámetro 2-4 cm) y tercera (diámetro < 2 cm). El número de plantas cosechadas en el surco fue usado como covariable. Para analizar la interacción Genotipo x Localidad se realizó un análisis combinado.
La evaluación preliminar de resistencia a P. infestans, se realizó empleando la prueba de foliolos en cámaras húmedas. Los foliolos fueron inoculados en experimentos independientes con aislamientos de P. infestans correspondientes a la raza cero y a una raza compleja H1N (9 genes) colectada en el departamento de Antioquia. El primer experimento buscó evaluar la presencia o ausencia de genes mayores de resistencia a P. infestans. El segundo, buscaba identificar entradas con potencial resistencia horizontal, mediante comparación cuantitativa del tamaño de las lesiones cuando se utilizó la raza compleja H1N de P. infestans. Se utilizaron como testigos las variedades Diacol Capiro (muy susceptible), Diacol Monserrate (resistencia horizontal) y Clon 1 (cultivar comercial tipo "Yema de huevo" susceptible). Las pruebas se realizaron en el laboratorio de Fitopatología de la Facultad de Agronomía de la Universidad Nacional (sede Bogotá), empleando la siguiente metodología: Obtención de foliolos. Los foliolos fueron recolectados de la colección sembrada en la localidad San Jorge, del tercio superior de la planta, antes de floración, de cada hoja solo se tomó el foliolo terminal para inoculación, los foliolos fueron lavados con agua destilada.
Obtención del inóculo. El inóculo para las dos pruebas fue obtenido de dos multiplicaciones sucesivas empleando medio de cultivo Agar-Arveja, bajo condiciones de laboratorio. La suspensión para inoculación fue preparada con aislamientos sembrados 15 días antes de la prueba. El micelio se lavó con agua destilada para la extracción de esporangios y se filtró para eliminar residuos de micelio, la calibración del inóculo se realizó con la ayuda de un hematocitómetro a una concentración de 10.000 esporangios/ml.
Inoculación y evaluación. Los foliolos se colocaron en cajas de petri, con papel absorbente humedecido (2,5 mL de agua destilada). Por cada genotipo se hicieron 12 cámaras húmedas, seis fueron inoculadas con raza cero y las seis restantes con la raza compleja (H1N). La inoculación se realizó por adición de una gota de 20 microlitro (µL) de la suspensión de esporangios en el extremo proximal, cerca de la nervadura central. Las cajas fueron selladas con parafilm evitando disminuciones de humedad relativa dentro de la caja. Se mantuvieron a una temperatura de 18 °C y un fotoperiodo de 12 horas luz. La evaluación se realizó 6 días después de la inoculación. Se evaluó la posibilidad de presencia de genes mayores de resistencia a P. infestans teniendo en cuenta la formación de lesión, su tipo y la presencia o no de micelio. El indicio de resistencia horizontal fue evaluado por el diámetro de la infección medido paralelo a la nervadura central del foliolo. En este ensayo se utilizó un DCA, análisis de varianza y prueba de Tuckey.
Resultados y discusión
Peso de tubérculos de primera (PT1)
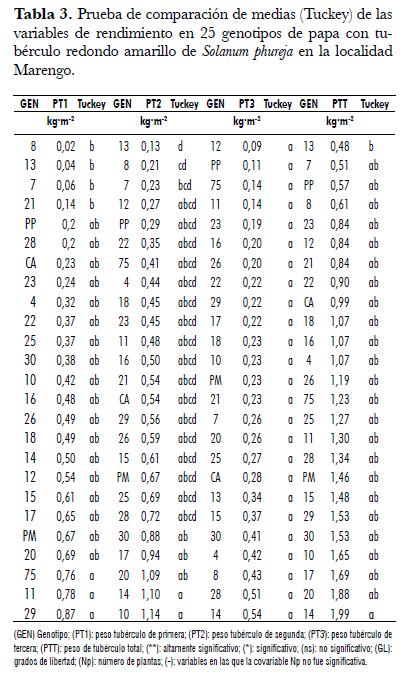
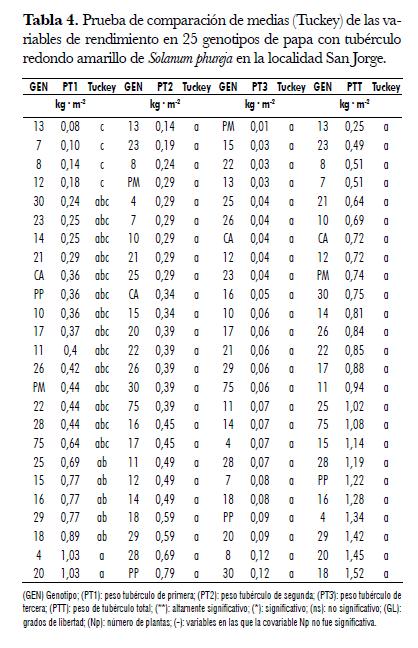
Esta variable presentó diferencias entre genotipos en las dos localidades. En la localidad San Jorge la covariable número de plantas fue significativa, por lo cual re realizó análisis de covarianza (tab. 1). En Marengo, el rango de la variable PT1 osciló entre 0,02 y 0,87 kg· m-2 correspondientes a los genotipos 8 y 29 respectivamente. Los genotipos 8, 13, 7 y 21 fueron inferiores a los genotipos 75, 11 y 29, e iguales a los restantes (tab. 3). En la localidad San Jorge, el rango de la variable osciló entre 0,08 y 1,03 kg· m-2, correspondientes a los genotipos 13 y 20 respectivamente. La prueba de t student realizada con los datos ajustados por la covarianza, mostró que los promedios de los genotipos 13, 7, 8 y 12 fueron inferiores a los promedios de siete genotipos (25, 15, 16, 29, 18, 4, 20) cuyas producciones fueron superiores a 0,65 kg· m-2 (tab. 4). Como se aprecia en las dos localidades los genotipos 7, 8 y 13 fueron los de menor respuesta en esta variable, este resultado permite decir que es factible realizar selección para la variable y adelantar mejoramiento genético de este carácter. En el análisis combinado se encontraron diferencias entre genotipos y no se encontraron diferencias para localidades, ni para la interacción Genotipo x Localidad, lo cual nos indica que la respuesta de los genotipos para esta variable fue similar en los ambientes de evaluación (tab. 2).
Peso de tubérculos de segunda (PT2)
En la localidad Marengo, la variable PT2 presentó diferencias entre genotipos. El rango de esta variable osciló entre 0,13 y 1,14 kg· m-2 correspondientes a los genotipos 13 y 10 respectivamente. La prueba de Tuckey permitió establecer que el genotipo 13, fue inferior a cinco genotipos (10, 20, 14, 30, 17), los cuales presentaron valores promedios superiores a 0,725 kg· m-2. Los promedios de los genotipos 10, 14 y 20 fueron superiores al promedio del genotipo 8; de igual forma, los genotipos 10 y 14 superaron al genotipo 7 (tab. 3). En la localidad San Jorge, para esta variable no se encontraron diferencias significativas entre genotipos y, el rango observado en esta localidad osciló entre 0,15 y 0,8 kg· m-2, correspondientes a los genotipos 13 y PP respectivamente (tab. 4).
En el análisis combinado de esta variable, se encontraron diferencias para genotipos, localidades y la interacción Genotipo x Localidad, lo que evidencia que la respuesta de los genotipos en esta variable fue diferente en las dos localidades, ello explicable seguramente por la existencia de adaptaciones de genotipos a ambientes determinados. Arias y Bustos (1996), obtuvieron para esta variable en la localidad Marengo, un promedio de 0,32 kg· m-2, valor que fue superado por el promedio de 20 genotipos de la colección en esta localidad y por 16 en la localidad San Jorge. Es importante destacar que al ser PT2 la variable que mayor participación tiene en la producción, los valores obtenidos en el ensayo muestran el potencial de la colección desde el punto de vista comercial.
Peso de tubérculos de tercera (PT3)
En esta variable en ninguna de las localidades se encontró diferencias entre genotipos (tab. 1). En la localidad Marengo el rango de esta variable osciló entre 0,1 y 0,55 kg· m-2 correspondientes a los genotipos 12 y 14 respectivamente (tab. 3), en tanto que, el rango en la localidad San Jorge osciló entre 0,015 y 0,13 kg· m-2 correspondientes a los genotipos PM y 30 (tab. 4). La interacción Genotipo x Localidad no fue significativa para esta variable, pero si se presentaron diferencias entre localidades, siendo superior la respuesta de la variable en la localidad Marengo.
Peso total de tubérculos (PTT)
La variable PTT en la localidad Marengo presentó diferencias entre genotipos (tab. 1). En esta localidad la variable osciló entre 0,53 y 2,16 kg· m-2 correspondientes a los genotipos 13 y 14, respectivamente, siendo ellos diferentes entre sí. Entre los genotipos restantes no se encontraron diferencias en la respuesta de la variable (tab. 3). En San Jorge no se encontraron diferencias entre genotipos para esta variable. El rango osciló entre 0,26 y 1,59 kg· m-2 correspondientes a los genotipos 13 y 18 respectivamente (figura 1 , tab. 3).
En el análisis combinado de la variable se encontraron diferencias entre localidades y genotipos, siendo superior el peso total de tubérculos en la localidad Marengo, lo cual sugiere que bajo condiciones de la Sabana de Bogotá se encuentran mejores potenciales de rendimiento. No se encontró interacción Genotipo x Localidad, lo que significa que los genotipos en producción total presentaron respuestas similares en los ambientes de evaluación, por lo tanto, la selección de genotipos superiores por esta variable se puede realizar sin mayor problema en cualquiera de los ambientes evaluados, aunque este resultado debe verificarse en un número mayor de ambientes (tab. 2).
Estudio de la resistencia a P. infestans
Presencia de genes mayores. La respuesta de los genotipos frente a la inoculación de la raza cero, permitió determinar ausencia de genes mayores, debido a que todos los genotipos redondos amarillos presentaron infección, diferente a lo observado en la variedad testigo Diacol Capiro, que no presentó infección, lo cual evidencia que por lo menos tiene un gen mayor de resistencia a P. infestans. Este resultado está de acuerdo con lo reportado por Forbes et al. (1994).
Evaluación preliminar de resistencia horizontal. Se evalúo el diámetro de la lesión en los foliolos inoculados con la raza compleja H1N, variable que permitió establecer que existen diferencias entre los genotipos en la respuesta a la enfermedad (figura 2 ), lo que evidencia la posible existencia de resistencia horizontal en la colección evaluada. El genotipo PM presentó un promedio de diámetro de la lesión inferior a la variedad Monserrate, variedad testigo en la cual se reconoce resistencia horizontal y, un grupo importante de genotipos presentó respuesta en la variable similar al genotipo PM. Este resultado seguramente se debe a la cercanía genética de los materiales y plantea la necesidad de evaluar la respuesta de la variable en condiciones de campo (Figura 2). Es igualmente importante destacar la respuesta observada en la variedad Diacol Capiro, la cual presentó el segundo mayor promedio para la variable, evidenciándose claramente su alta susceptibilidad a la enfermedad, al igual que los genotipos de la colección 11, 13 y 30. El resultado encontrado en la evaluación es muy positivo, porque permitió identificar genotipos que deben ser estudiadas con mayor profundidad, de tal manera que puedan ser incluidos dentro del programa de mejoramiento para este carácter.
Literatura citada
Argüello, G. 2004. Director "Proyecto para la producción, industrialización y comercialización de la papa criolla (Solanum phureja)." Convenio Fedepapa-Fenalce. Comunicación personal. [ Links ]
Arias, V. y P. Bustos. 1996. Evaluación de rendimiento y algunos aspectos fisiológicos en papa criolla Solanum phureja bajo diferentes densidades de siembra en la Sabana de Bogotá. Trabajo de grado. Facultad de Agronomía, Universidad Nacional, Bogotá. 120 p. [ Links ]
Barrera, O. 1994. Mercadeo interno de la papa criolla. Revista Papa 10, 19-27. [ Links ]
Carslile, D.J., L.R. Cooke, S. Watson y A. Brown. 2002. Foliar aggressiveness of Northern Ireland isolates of Phytophtora infestans on detached leaflets of three potato cultivars. Plant Pathology 51, 424-434. [ Links ]
Chacón, M. 1995. Detección de genes mayores en especies silvestres, variedades nativas mejoradas y diploides de papa y evaluación de la resistencia en campo de especies silvestres a Phytophthora infestans. Trabajo de Grado. Facultad de Agronomía, Pontificia Universidad Católica de Ecuador. 98 p. [ Links ]
Del Valle, A. 1994. Papa amarilla, yema de huevo o papa "criolla" colombiana. Revista Papa 10, 3. [ Links ]
FEDEPAPA. 1988. La papa criolla una alternativa agroindustrial. Boletín de información técnica No. 103. [ Links ]
Flier, W.G.; G.B.M. van Den Bosch y L.J. Turkensteen. 2003. Stability of partial resistance in potato cultivars exposed to aggressive strains of Phytophtora infestans. Plant Pathology 52, 326-337. [ Links ]
Forbes, G.A. y M.C. Jarvis. 1994. Host resistance for management of potato late blight. In: Advances in potato pest biology and management. APS, St. Paul, Minnesota. 446 p. [ Links ]
Hawkes, J.G. 1990. The potato evolution, biodiversity and genetic resource. Smithsonian Institution Presss, Washington D.C. 259 p. [ Links ]
López, G. 1977. Crecimiento del cultivo de la papa. En: Prácticas culturales en papa. ICA, Pasto. 40 p. Luján, L. 1994. La ecología de la papa. Revista Papa 12. [ Links ] [ Links ]
Malamud, O.S.; T. Ames de Icoechea y H. Torres. 1994. Potato disease management in Latin America. En: Advances in potato pest biology and management. APS, St. Paul, Minnesota. pp. 616-626. [ Links ]
Pineda, C.R. 1995. Perspectivas para el desarrollo agroindustrial del cultivo de la papa en Colombia. Revista Papa 13, 4-14. [ Links ]
Sieczka, M.; B. Flis; M. Pakosinska y B. Tatarowska. 2002. Resistance to Phytophthora infestans in selected potato cultivars bred in former Soviet Union Republics (Poland). En: http://www.cipotato.org/gilb/Conf2002/AbstractsBreeding.pdf consulta: mayo 2005. [ Links ]
Zimnoch-Guzowska, E.; R. Lebecka; A. Kryszczuk; U. Maciejewska; A. Szczerbakowa y B. Wielgat. 2003. Resistance to Phytophtora infestans in somatic hybrids of Solanum nigrum L. and diploid potato. Theoretical and Applied Genetics 107, 43-48. [ Links ]

