










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkAgronomía Colombiana
versión impresa ISSN 0120-9965
Agron. colomb. v.23 n.1 Bogotá ene./jul. 2005
Estandarización de la técnica molecular de AFLP en palma de aceite tipo Dura (Elaeis guineensis Jacq.) y estudio preliminar de caracterización
Standardising Amplified Fragment-Length Polymorphisms (AFLP) for Dura oil palm (Elaeis guineensis Jacq.) and preliminary molecular characterization studies
Carlos Hernando Galeano1
1Investigador Asistente, Centro Internacional de Agricultura Tropical (CIAT), Palmira. e-mail: chgaleanom@unal.edu.coFecha de recepción: 15 de diciembre de 2004 Aceptado para publicación: 27 de mayo de 2005
Resumen
La palma de aceite (Elaeis guinnensis Jacq.) desempeña un importante papel en la economía de algunos países tropicales; Colombia ocupa el primer puesto en producción en América Latina y el quinto lugar a nivel mundial. En esta especie el mejoramiento genético busca generar materiales con alta producción de aceite y adaptados a las características edafoclimáticas de las diferentes zonas palmeras. El objetivo del presente trabajo fue estandarizar la técnica de AFLP (Amplified Fragment-Length Polymorphisms) en materiales Dura del Programa de Fitomejoramiento de CENIPALMA. Al protocolo de AFLP de la casa comercial se le realizaron algunas modificaciones, como el aumento de la concentración del ADN (270 ng· µL-1) y la dilución 1:10 del producto de la preamplificación. De igual forma, se determinó que para teñir los geles de poliacrilamida con nitrato de plata es necesario usar agua destilada deionizada (Milli Q®). También se realizó un estudio preliminar de caracterización molecular en 12 palmas de los tipos Dura, Ténera y E. oleifera; según los agrupamientos hechos según métodos fenéticos y multivariados se encontró que la mayoría de palmas Dura se agrupan según el lugar de colecta, mientras la palma de origen americano se ubica de forma distante al compararla con las palmas de origen africano.
Palabras clave adicionales: Marcadores moleculares, diversidad genética, similaridad genética, ADN.
Summary
Oil palm (Elaeis guinnensis Jacq.) plays an important role in some tropical countries' economies; Colombia occupies first place in production in Latin-America and fifth throughout the world. The aim of palm breeding research is producing palm having high oil production, adapted to different climatic condition. Cenipalma is currently using molecular tools which can reduce environmental effect and selection time. This research was aimed at standardising the Amplified Fragment-Length Polymorphisms (AFLP) technique for Dura material from Cenipalma's breeding programme. Modifications were made to the commercial AFLP kit protocol such as increasing initial DNA concentration to 270 ng· µL-1. Two amplification product dilutions (1:10 and 1.50) were evaluated, 1:10 dilution being the best for the AFLP technique. It was also found that deionised distilled water was needed for staining polyacrylamide gels with silver nitrate (Milli Q®). A preliminary analysis of genetic diversity was carried out on 12 palms (Dura, Ténera and E. oleifera). Clustering and principal coordinate analysis results revealed that most Duras clustered according to where they are collected and that the American palm clustered alone when compared to African palms.
Additional key words: Molecular marker, genetic diversity, genetic similarity, DNA.
Introducción
LA PALMA DE ACEITE (Elaeis guineensis Jacq.) es una planta perenne tropical originaria de áfrica Central y Occidental que se cultiva ampliamente en el trópico húmedo (Corley y Tinker, 2003), donde alcanzó un área total en producción cercana a 7,5 millones de hectáreas en el año 2002, de las cuales Colombia participó con 145.027 ha distribuidas en cuatro regiones productoras. Colombia es el quinto productor a nivel mundial y el primero en el continente americano, con una producción cercana a 577.117 t que incluye los aceites de palma y de palmiste (Fedepalma, 2003). Los programas de mejoramiento en palma de aceite están orientados hacia el aumento de la producción de racimos por palma y, por lo tanto, al incremento de la producción de aceite. En Colombia, además de este objetivo, deben identificarse materiales que presenten buena adaptación a las zonas agroecológicas palmeras y que sean resistentes a enfermedades y plagas limitantes del cultivo (Ayala y Gómez, 2000). Como estrategia, los programas de mejoramiento se basan en la selección de materiales tipo Dura (progenitor femenino) y plantas Pisíferas (progenitor masculino) para la posterior evaluación de progenies por alta producción de aceite.
El Centro de Investigación en Palma de Aceite (CENIPALMA) busca consolidar un Programa de Fitomejoramiento de palmas tipo Dura presentes en diferentes plantaciones del país. De esta manera, se contribuirá a evitar la 'erosión genética' y se generarán plantas madre con una combinación deseable de genes que confieran óptima adaptación a las condiciones de las zonas productoras colombianas (Rey et al., 2003).
Para cumplir con estos objetivos es esencial conocer la variabilidad genética existente dentro de la especie o especies intercruzables, a fin de identificar individuos con características deseables y la fuente de genes disponible para el mejoramiento del cultivo, además de obtener la mejor adaptación con los más altos rendimientos. Con el fin de planificar cruzamientos, se están utilizando marcadores moleculares tipo RAPD (Randomly Amplified Polymorphic DNA; Polimorfismo en el ADN Amplificado Aleatoriamente) y AFLP (Amplified Fragment-Length Polymorphisms, Polimorfismos de Longitud de Fragmentos Amplificados) como herramientas adicionales al registro individual de producción y de medidas vegetativas (Villegas et al., 2000; Rocha, 2002).
Los marcadores moleculares son secuencias de ADN que se han constituido en herramientas valiosas para la detección y el uso de la diversidad genética, con su consecuente aplicación en programas de selección genética de plantas, animales y microorganismos. Los polimorfismos de ADN pueden ser detectados mediante técnicas moleculares, algunas de las cuales han sido empleadas en palma de aceite en el análisis de variabilidad genética y el establecimiento de relaciones filogenéticas (Billotte et al., 2001), desarrollo de mapas de ligamiento (Mayes et al., 1997), señalización y seguimiento de genes de interés (árias y Rocha, 2004; Rance et al., 2001).
La técnica de AFLP combina la especificidad, la resolución y el poder de muestreo de la digestión con enzimas de restricción, con la velocidad y la facilidad para detectar polimorfismos vía PCR (Vos et al., 1995). Con esta técnica se han obtenido un gran número de marcadores moleculares distribuidos en genomas de procariotas y eucariotas. En palma se han utilizado en estudios de diversidad genética del género Elaeis (Barcelos et al., 2000; Barcelos, 1998), estudios de ligamiento genético a características como grosor de cuesco (Billotte et al., 2001), detección de variación somaclonal (Matthes et al., 2001) y estudios de progenie (Rajinder et al., 1998). El objetivo de este trabajo fue estandarizar la técnica de AFLP en materiales Dura bajo las condiciones del laboratorio de biología molecular de CENIPALMA y realizar un estudio preliminar de caracterización molecular de palmas Dura, Ténera y E. oleifera. Estos materiales presentan características de interés para programas de mejoramiento genético y la técnica orientará la selección de palmas Dura.
Materiales y métodos
Material vegetal
El material vegetal utilizado para la estandarización de AFLP correspondió a palmas Dura de la Zona Central (departamento de Santander, municipio de Puerto Wilches, Plantación Agropecuaria Monterrey) que presentaron altos registros de producción; las muestras consistieron en foliolos jóvenes y sanos de la hoja 17. Posteriormente, se colectaron otros materiales de palma de aceite de las Zonas Central y Norte (departamento del Atlántico, municipio de Fundación, Plantación Padornelo) de los cuales se seleccionaron 12 para realizar una evaluación preliminar de la diversidad genética de las muestras, las cuales comprendían materiales de los tipos Dura, Ténera y E. oleifera (tab. 1).
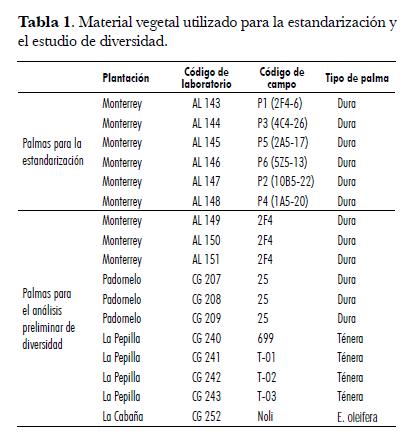
Extracción del ADN genómico
La metodología se realizó de acuerdo con el protocolo del laboratorio de marcadores moleculares de CENIPALMA (Rocha et al., 2003). El tejido foliar fue pulverizado con ayuda de nitrógeno líquido y en un tubo Epependorff de 1,5 mL se tomó una cantidad aproximada de 500 mg de tejido molido. Luego se agregaron 1,2 mL de solución de extracción caliente (Tris HCl 100 mM, EDTA 50 mM, NaCl 500 mM, ß-mercaptoetanol 10 mM y PVP 1% [p/v]) y posteriormente se agregaron 85 µL de SDS (dodesil sulfato de sodio) al 20%. Los tubos se incubaron en baño maría a 65°C durante 30 min y luego se agregaron 390 µL de acetato de potasio 5 M. La mezcla se centrífugó a 12.500 xg durante 15 min y el sobrenadante se transfirió a un tubo, al cual se adicionó 1 mL de isopropanol frío; la mezcla se incubó a –20° C durante toda la noche. Después se centrifugó a 25.000 xg durante 30 min y se desechó el líquido teniendo cuidado de no perturbar el botón de ADN, el cual se lavó con 2 mL de etanol al 70% para luego centrifugarlo a 25.000 xg durante 3 min. Finalmente, se dejó secar el botón a temperatura ambiente y se le adicionaron 350 µL de solución TE-ARNasa (10 mM de Tris HCl a pH 8,0; 1 mM de EDTA a pH 8,0; 5 mg· mL-1 de ARNasa) y se incubó a 37° C durante 15 min. El ADN obtenido se almacenó a 4° C para su posterior utilización. Para la cuantificación del ADN se utilizó un método comparativo teniendo como patrón de referencia un ADN genómico de tomate de concentración conocida. Se realizó una electroforesis en gel de agarosa al 0,7 % (p/v) en donde se colocó el ADN de tomate a diferentes concentraciones, el cual se comparó con el ADN de las muestras. La concentración de las muestras se determinó según la intensidad de fluorescencia emitida por el bromuro de etidio.
Técnica de AFLP
Para la técnica molecular AFLP (Amplified Fragment Length Polymorphisms, Polimorfismos de Longitud de Fragmentos Amplificados) se utilizó el kit AFLP Analysis System I? siguiendo el protocolo del fabricante con algunas modificaciones. La digestión se realizó con las enzimas EcoR I/Mse I (1,25 U · µL-1) al incubarlas con el ADN (270 ng) durante 2 h a 37°C. A los fragmentos de restricción se las adicionó 24 µL de una mezcla de adaptadores que, junto con la enzima T4 DNA ligasa, se unieron a los extremos cohesivos dejados por las enzimas de restricción. Para la preamplificación se tomaron 5 µL de una dilución 1:10 del resultado de la ligación, y se agregaron 40 µL de la mezcla de cebadores con un nucleótido seleccionante (+1), 5 µL de buffer 10X con Mg, y 1 µL de Taq DNA polimerasa. El termocilador se programó a 94° C durante 30 segundos (s), 56° C durante 60 s y 72° C durante 60 s, por 20 ciclos. El resultado de esta PCR se corroboró con una electroforesis en gel de agarosa al 1% y posteriormente se realizó una dilución 1:10.
Para la amplificación se tomaron 5 µL de la dilución, 5 µL del mix 1 (primers +3 y dNTP's) y 10 µL del mix 2 (taq polimerasa, buffer 10X) según protocolo del fabricante. El termociclador se programó a 94° C durante 30 s para denaturar la molécula, 65° C durante 30 s para alinear los cebadores y 72° C durante 60 s para la polimerización. Posteriormente, la temperatura de alineamiento bajó 0,7° C en cada ciclo durante 12 ciclos (touch down) y continuó con 94° C durante 30 s, 56° C durante 60 s y 72° C por 60 s, a lo largo de 23 ciclos.
El producto de la amplificación se observó en una electroforesis en gel de poliacrilamida al 5% y 8 M de úrea a 100 W durante dos horas, o hasta que la banda de xilene-cyanol llegara a la parte inferior del gel.
Para la tinción se colocó el gel en una solución ácido acético al 10% durante 20 minutos (min), luego se lavó con agua destilada deionizada dos veces durante 1 min cada vez. Posteriormente se incubó en una solución de tinción (2 g de AgNO3, 3 mL de formaldehidol 37% en 2 L de agua) durante 30 min. El exceso de plata se retiró lavando con agua destilada deionizada durante 10 seg. Después, se sumergió en la solución de revelado (60 g de Na2CO3, 3 mL de formaldehído al 37% y 400 µL de tiosulfato de sodio [10 mg· mL-1] en 2 L de agua) durante 5 a 10 min hasta visualizar las bandas. Finalmente, el gel se colocó a una solución de ácido acético al 10% para detener la reacción y se lavó con agua destilada deionizada.
Análisis de datos
Las bandas polimórficas en el gel fueron contadas visualmente con la ayuda de un transiluminador de luz blanca; una banda se considera polimórfica si está presente al menos en un individuo y ausente en los otros. Se generó una matriz binaria en la que las bandas fueron contadas como '1' presente y '0' ausente. La matriz binaria de datos fue convertida a una matriz de similaridad de acuerdo con la definición de Nei y Li (1979):
Sij = 2a / (2a + b + c)
Donde Sij es la similitud entre los individuos i y j; a es el número de bandas presentes en ambos i y j; b es el número de bandas presentes en i y ausentes en j; y c es el número de bandas presentes en j y ausentes en i. El dendograma fue construido a partir de la matriz de similaridad agrupando los datos con el método UPGMA (Unweighted Pair-Group Method Arithmetic Average). El coeficiente de correlación cofenética fue calculado entre la matriz de similaridad y la matriz cofenética. El análisis de coordenadas principales se basó en la matriz de similaridad utilizando los algoritmos DCENTER y EIGEN. Todos los datos se analizaron con el software NTSYS-pc® versión 2.11L (Rohlf, 2000).
Resultados y discusión
Extracción y cuantificación
El protocolo de extracción de ADN difirió del utilizado previamente en palma (Rocha, 2002), el cual resultaba dispendioso y costoso, y hacía difícil la extracción de un buen número de muestras. Uno de los cambios fue el uso de SDS (dodecil sulfato de sodio), un detergente aniónico cuya función principal es la de solubilizar proteínas y membranas; su utilización para una óptima extracción de ADN a partir de pequeñas cantidades de tejido foliar se reporta en varios protocolos (Quenzar et al., 1998; Li et al., 2002). A fin de prevenir la presencia de polifenoles que pueden unirse mediante enlaces covalentes a proteínas y ácidos nucleicos y generar un ADN de baja calidad, se recurrió –además del PVP y el SDS–, al ß-mercaptoetanol por su función inhibitoria de la oxidación de compuestos fenólicos (Li et al., 2002). Este procedimiento arrojó muy buenos resultados pues fue posible obtener ADN de alta calidad (figura 1 ).
Además, el método de cuantificación citado permite determinar la calidad del ADN extraído e identificar compuestos como ARN o proteínas que pueden interferir en las reacciones posteriores, acarreando problemas como una insuficiente digestión por parte de las enzimas en el caso de AFLP. La cuantificación del ADN mostró cantidades bastante diferentes, dependiendo de la calidad (fisiológica y sanitaria) del foliolo utilizado. En la figura 1 se observa que el ADN no tiene impurezas (ARN o proteínas) y que las bandas no presentan degradación por parte de las ADNasas. Estas características son las ideales para iniciar el protocolo de AFLP, ya que se necesita ADN de alta pureza para garantizar la digestión completa por parte de las enzimas de restricción en todas las muestras y así no tener problemas en la interpretación de los polimorfismos (Ferreira y Grattapaglia, 1998). Determinaciones electrofotométricas sobre la cantidad necesaria de ADN para iniciar trabajos con AFLP son reportadas por Ubi et al. (2003) al correr alícuotas de ADN en gel de agarosa al 8% (p/v) y compararlas con cadenas no digeridas del fago Lambda.
Técnica de AFLP
La primera modificación implementada fue la cantidad de ADN con que se inició la digestión enzimática al utilizar 270 ng, 20 ng más de lo sugerido por la casa comercial. Modificaciones similares se han reportado en papa (McGregor et al., 2002) especie de la que utilizaron entre 300 y 400 ng de ADN; de igual forma en café (Anthony et al., 2002), para lo cual se usaron alícuotas de 500 ng de ADN genómico; ello señala que el aumento de la concentración del ADN inicial genera resultados óptimos. Por otra parte, el incremento de la cantidad de ADN al inicio del protocolo de AFLP ha sido estudiado por Zhu et al. (1998) y Vos et al. (1995) quienes evaluaron cantidades entre 0,2 µg y 1,0 µg y determinaron que los perfiles de las bandas no son significativamente sensibles a la concentración inicial del ADN.
Posteriormente, el producto de preamplificación se diluyó 1:50 según las recomendaciones del manual, y a 1:10 según Julier et al. (2003) para posteriormente realizar la reacción de amplificación con los cebadores +3. Por último, los productos de PCR +3 se observaron en una electroforesis en gel de poliacrilamida al 5% sobre el cual se evaluaron las dos diluciones de la preamplificación (1:10 y 1:50) (figura 2A ); para la tinción se utilizó agua de consumo humano (Santa Clara®). En la figura 2A se evidencia que la concentración 1:50 recomendada por el fabricante de AFLP's no fue óptima para la palma, mientras que en la concentración 1:10 se pueden ver un buen número de las bandas. Este resultado es coherente con las modificaciones que se realizan al protocolo de AFLP en los trabajos de Rajinder et al. (1998) y Julier et al. (2003). El ajuste en la dilución del producto de la preamplificación puede realizarse de 1 a 50 veces, dependiendo de la intensidad de las bandas (Zhu et al., 1998), lo que permite obtener mejor resolución en los geles.
El otro problema que se observa en la figura 2A es la deficiente calidad de la tinción cuando se utiliza agua de consumo humano que tiene contaminantes, especialmente halógenos y/o iones metálicos, que generan una baja intensidad de las bandas, razón por la cual se recomienda la utilización de agua destilada y deionizada para tal fin.
Por lo anterior, se evaluaron tres tipos de agua: de consumo humano, destilada y Milli Q?. Las dos primeras generaron precipitación del carbonato de potasio, al igual que turbidez en la solución con nitrato de plata. El resultado final del uso de este tipo de agua son geles manchados con bandas muy tenues. Los análisis de conductividad eléctrica corroboraron los resultados, al encontrar mayor cantidad de sales en el agua de consumo humano (190 µmhos), y reducida cantidad de las mismas en el agua Milli Q? (1,7 µmhos), opción que mostró mejor desempeño para la tinción de los geles. En la figura 2B se puede observar un buen número de bandas polimórficas, cerca de 30 a 40 entre los 100 y 800 pb, un resultado similar al reportado por Rajinder et al. (1998) en palma de aceite.
Estudio preliminar de caracterización molecular
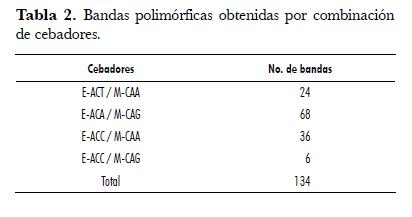
En las 12 muestras evaluadas (tab. 1) se detectaron un total de 134 bandas polimórficas con el uso de cuatro combinaciones de cebadores (tab. 2). La combinación de cebadores E-ACA/M-CAG presenta el mayor número de polimorfismos con 68 bandas polimórficas, a diferencia de la combinación E-ACC/M-CAG que muestra tan sólo 6 bandas polimórficas; la alta cantidad de bandas polimórficas es una primera indicación de la variación genética presente en los individuos evaluados. Estos resultados son consistentes con los polimorfismos detectados en palma por Maizura & Rajanaidu (2003) quienes usaron ocho combinaciones de cebadores, generando un total de 228 bandas polimórficas. De igual forma, Purba et al. (2000) reportaron la generación de 96 bandas polimórficas utilizando cinco combinaciones de cebadores.
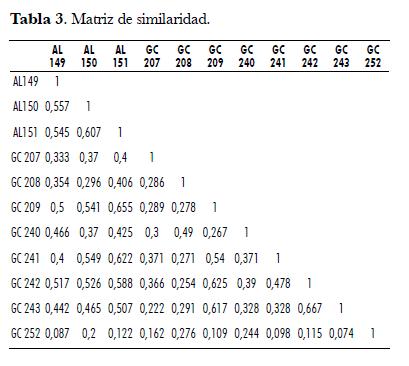
La variación genética entre los 12 materiales fue estimada usando el coeficiente de similaridad sugerido por Nei & Li (1979), el cual se recomienda para el análisis de datos de marcadores dominantes, pues da más importancia a las bandas compartidas o coincidentes (Zeid, 2003). En la matriz de similaridad (tab. 3) se evidencian relaciones muy cercanas entre las palmas Ténera que mostraron índices de similaridad de 0,667 (GC 243 y GC 242); por otra parte, se encuentran bajos valores de similaridad (0,074 a 0,2) al comparar la palma de origen americano (GC 252) con el resto de palmas de origen africano.
El análisis de agrupamientos genera un dendrograma (figura 3) con una correlación cofenética alta (r = 0,92), lo cual indica que el fenograma representa fielmente los valores de la matriz de similitud (Crisci y López, 1983). En el árbol se puede evidenciar dos grandes agrupamientos: el primero reúne todas las palmas tipo Dura colectadas en la Zona Central (Plantación Monterrey), con excepción de CG 209, la cual se colectó en la Zona Norte, junto con algunas tipo Ténera. En el segundo grupo se ubican las restantes palmas Dura de la zona norte, junto con otras Ténera.
El agrupamiento de las palmas tipo Dura colectadas en la Zonas Central y Norte con las palmas tipo Ténera, se puede explicar al conocer el origen híbrido de las palmas Ténera, ya que provienen del cruce de Dura x Pisífera. Similares agrupamientos de materiales Dura con Ténera son reportados en los trabajos de Maizura y Rajanaidu (2003).
Los resultados del análisis de coordenadas principales de la figura 4, muestran la distribución de las muestras en tres coordenadas las cuales explican el 50,3 % de la variación. En dicha figura se puede corroborar de forma más precisa los agrupamientos evidenciados en el dendrograma de la figura 3, lo que permite afirmar que existe una interesante diferenciación genética en los individuos evaluados.
Por último, en las figuras 3 y 4 se puede observar que la palma de aceite de origen americano (GC 252) se agrupa aparte de las palmas de origen africano. Resultados similares son reportados en palma de aceite por Bilotte et al. (2001) y Barcelos et al. (2002) quienes usaron microsatélites, y AFLP y RFLP, respectivamente. Por el contrario, solamente el trabajo publicado por Arias y Rocha (2004) reporta una corta distancia genética entre la palma americana y la africana generando un dendrograma donde ambas especies se mezclan y evidenciando que los marcadores microsatélites, utilizados en dicho trabajo, no pueden distinguir entre E. oleifera y E guineensis o problemas el etiquetamiento del material.
Conclusiones
Para los objetivos del Programa de Fitomejoramiento de CENIPALMA la diferencia genética existente entre los grupos de las palmas tipo Dura es de gran interés, cuando se busca seleccionar materiales tipo Dura que sirvan para realizar cruzamientos DxD y así generar progenies segregantes con variabilidad genética que puedan brindar más ganancia en la selección de palmas madre. Además, la viabilidad de los cruzamientos interespecificos en palma hace que la especie E. oleifera constituya una de las estrategias para aumentar la base genética en los programas de mejoramiento de palma de aceite en Colombia.
Agradecimientos
Agradezco a CENIPALMA por acogerme como tesista y brindarme el apoyo logístico y económico necesario para esta investigación. Así mismo, a los Doctores Pedro Rocha y Leonardo Rey por la dirección, consejo, corrección y ayuda en la elaboración de este trabajo. De igual forma, al profesor Oscar Oliveros de la Universidad Nacional por sus correcciones y sugerencias.
Literatura citada
Anthony F.; M.C. Combes; C. Astorga; B. Bertrand; G. Graziosi y P. Lashermes. 2002. The origin of cultivated Coffea arabica varieties revealed by AFLP and SSR markers. Theoretical Applied Genetics 104(5), 894-900. [ Links ]
Arias, D. y P. Rocha. 2004. Análisis de la diversidad genética en materiales tolerantes y susceptibles a la pudrición de cogollo en palma de aceite mediante marcadores moleculares. Palmas 25(3), 11-27. [ Links ]
Ayala, L.; P. Gómez y C. Durán. 2000. Selección de progenitores Dura adaptados a las condiciones del Magdalena Medio colombiano. Palmas 21(1), 25-34. [ Links ]
Barcelos, E. 1998. étude de la diversité génétique du genre Elaeis (E. oleifera (Kunth) Cortés et E. guineensis Jacq.) par marqueurs moléculaires (RFLP et AFLP). Thèse de Docteur, Université de Montpellier II, Montpellier. 137 p. [ Links ]
Barcelos, E.; P. Amblard; J. Berthaud y M. Seguin. 2000. The genetic diversity of the American oil palm Elaeis oleifera (Kunth) Cortés, revealed by nuclear RFLP markers. Pesq. Agropec. Brasileira 37(8), 1105-1114. [ Links ]
Billotte, N.; A.M. Rusterucci; E. Barcelos; J.L. Noyer; P. Amblard y F.C. Baurens. 2001. Development, characterization and across-taxa utility of oil palm (Elaeis guineensis) microsatellite markers. Genome 44, 413-425. [ Links ]
Corley, R.H.V. y P.B. Tinker. 2003. The oil palm. Fourth edition. Blackwell, New York. 562 p. [ Links ]
Crisci, J. y M. López. 1983. Introducción a la teoría y práctica de la taxonomía numérica. Secretaría General de la Organización de los Estados Americanos OEA. Washington, D.C. 120 p. [ Links ]
FEDEPALMA. 2003. Anuario estadístico: La agroindustria de la palma de aceite en Colombia y el mundo. Fedepalma, Bogotá. Ferreira, M. y D. Grattapaglia. 1998. Introducción al uso de marcadores moleculares en el análisis genético. EMBRAPA, Brasilia. 221 p. [ Links ] [ Links ]
Julier, B.; S. Flajoulot; P. Barre; G. Cardinet; S. Santoni; T. Huguet y C. Huyghe. 2003. Construction of two genetic linkage maps in cultivated tetraploid alfalfa (Medicago sativa) using microsatellite and AFLP markers. BMC Plant Biology 3(9). En: http://www.biomedcentral.com/1471-2229/3/9 consulta: mayo 2005. [ Links ]
Li, Y.; Z. Su y F. Chen. 2002. Rapid extraction of genomic DNA from leaves and bracts of bove tree (Davidia involucrata). Plant Molecular Biology Reporter 20, 185a-185d. [ Links ]
Maizura, I. y N. Rajanaidu. 2003. Preliminary results on AFLP screening of the Malaysian advanced oil palm breeding population. Proceeding of the PIPOC 2003 International Palm Oil Congress. Kuala Lumpur, Malaysia. pp. 150-160. [ Links ]
Matthes, M.; R. Singh; S. Chean y A. Karp. 2001. Variation in oil palm (Elaeis guineensis Jacq.) tissue culture-derived regenerants revealed by AFLP's with methylation-sensitive enzymes. Theoretical and Applied Genetics 102, 971-979. [ Links ]
Mayes, S.; P. Jack; D. Marshall y R. Corley. 1997. The construction of a RFLP linkage map for oil palm. Genome 40, 116-122. [ Links ]
McGregor, C.; R. Treuren; R. Hoekstra y T. Hintum. 2002. Analysis of the wild potato germplasm of the series Acaulia with AFLP's: implications for ex situ conservation. Theoretical and Applied Genetics 104, 146-156. [ Links ]
Nei, M. y W. Li. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National Academy of Science 76, 5269-5273. [ Links ]
Purba, A.R.; J.L. Noyer; L. Baudouin; X. Perrier; S. Hamon y P.J. Lagoda. 2000. A new aspect of genetic diversity of Indonesia oil palm (Elaeis guineensis Jacq.) revealed by isoenzyme and AFLP markers and its consequences for breeding. Theoretical and Applied Genetics 101, 956-961. [ Links ]
Quenzar, B.; C. Hartman; A. Rode y A. Benslimane. 1998. Date palm mini-prepartin without liquid nitrogen. Plant Molecular Biology Reporter 16, 263-269. [ Links ]
Rajinder, S.; C. Suan-Choo y A. Rahimah. 1998. Generation of molecular markers in oil palm (Elaeis guineensis) using AFLPTM analysis. Plant Biotechnology 20, 26-27. [ Links ]
Rance, K.A.; S. Mayes; Z. Price; P.L. Jack y R.H.V Corley. 2001. Quantitative trait loci for yield components in oil palm (Elaeis guineensis Jacq.) Theoretical and Applied Genetics 103, 1302–1310. [ Links ]
Rey, L.; I. Ochoa; W. Delgado y P. Rocha. 2003. Colecta de material genético de palma Nolí Elaeis oleifera [H.B.K] Cortes en el Trapecio Amazónico. Ceniavances 101, 1-4. [ Links ]
Rocha, P. 2002. Teoría y práctica para la extracción del ADN de palma de aceite. Palmas 23, 9-17. [ Links ]
Rocha, P.; D. Árias y C. Galeano. 2003. Manual de protocolos del laboratorio de marcadores moleculares. CENIPALMA, documento interno de trabajo. [ Links ]
Rolhf, F.J. 2000. NTSYS–PC®. Numerical taxonomy and multivariate analysis system. Exeter Software. New York. Ubi, E.; R. Kölliker; M. Fujimori y T. Komatsu. 2003. Genetic diversity in diploid cultivars of rhodesgrass determined on the basis of Amplified Fragment Length Polymorphism markers. Crop Science 43, 1.516-1.552. [ Links ] [ Links ]
Villegas, V.; C. Durán y S. Beebe. 2000. Caracterización molecular de materiales Dura. Palmas 21, 35-40. [ Links ]
Vos, P.; R. Hogers; M. Bleeker; M. Reijas y T. Lee. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acid Research 23, 4407-4414. [ Links ]
Zeid, M. 2003. Analysis of genetic diversity based on molecular markers (AFLP) and the heterosis in faba bean (Vicia faba L). Ph.D. thesis. Faculty of Agricultural Sciences, Georg August University, Göttingen, Germany. [ Links ]
Zhu, J.; M. Gale; S. Quarrie; M. Jackson y G. Bryan. 1998. AFLP markers for study of rice biodiversity. Theoretical and Applied Genetics 96, 602-611. [ Links ]

