










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.23 no.1 Bogotá Jan./July 2005
Inducción de organogénesis indirecta en Abarco (Cariniana pyriformis Miers.) (Juz. et Buk.)
Inducing indirect organogenesis in Abarco (Cariniana pyriformis Miers.)
Mary Luz Yaya1,Olga Lucía Rodríguez2,William Usaquén3y Alejandro Chaparro4
1Bióloga, Laboratorio de Biología Molecular, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá. e-mail: mlyayal@unal.edu.co2Investigadora, Unidad de Biotecnología, Instituto SINCHI, Bogotá. e-mail: olgarod@softhome.com
3Instructor Asociado, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá. e-mail: wusaquenm@unal.edu.co
3Profesor Asistente, Laboratorio de Biología Molecular, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá. e-mail: achaparrog@unal.edu.co
Fecha de recepción: 14 de septiembre de 2004 Aceptado para publicación: 27 de mayo de 2005
Resumen
Se indujeron procesos primarios de morfogénesis de novo vía organogénesis indirecta en segmentos de hipocótilo de abarco, provenientes de plántulas in vitro de dos semanas de edad. El material fuente se obtuvo por germinación de embriones en Woody Plant Médium (WPM) diluido a la mitad y sin reguladores de crecimiento. La formación de callo organogénico se estimuló mediante la adición de ácido indolacético (AIA) conjuntamente con 6-bencilaminopurina (BAP). Después de cuatro semanas el mayor porcentaje de formación de callo (75%) se observó con la combinación AIA 1,21 µM y BAP 7,6 µM.
Palabras clave adicionales: Lecythidaceae, callo organogénico, brotes adventicios.
Abstract
Morphogenesis was induced in Cariniana pyriformis hypocotyl explants via indirect organogenesis. Explants were taken from two-week-old in vitro plants. The source material was obtained from germinating embryos in Woody Plant Medium (50%) without hormones. Callus formation was stimulated by IAA (indolacetic acid) and BAP (6-benzyl amino purine). The highest percentage (75%) of callus development (after four weeks) resulted from a combination of 1.21 µM IAA and 7.6 µM BAP.
Additional key words: Lecythidaceae, organogenic callus, adventitious bud.
Introducción
EL ABARCO (Cariniana pyriformis MIERS.) es una especie forestal que pertenece a la familia Lecythidaceae, la cual incluye aproximadamente 20 géneros y cerca de 440 especies de hierbas y árboles pantropicales (Morton et al., 1997). Los reportes bibliográficos indican que C. pyriformis se encuentra restringida a Panamá y Colombia, aunque existen especies muy similares en otros países. En nuestro país, su distribución se extiende a las tres cordilleras principalmente en la región del Magdalena Medio, Chocó, Urabá, las serranías de San Lucas, Abibe, San Jerónimo, Ayapel, Los Motilones y el Baudó. Su distribución natural corresponde a bosques húmedos y muy húmedos tropicales y a altitudes entre 500 y 800 m.s.n.m. (CONIF, 1996).
Debido a su alta calidad y durabilidad, la madera de abarco es muy apreciada, tanto en el mercado nacional como internacional, constituyéndose en una especie de alto potencial comercial para el país, razón por la cual hace parte de las especies vegetales promisorias incluidas en el convenio Andrés Bello (Bernal y Correa, 1994). Sus principales usos comprenden el área de construcción y fabricación de muebles; también es apta para construcciones navales, instrumentos de laboratorio y, en algunas regiones, es importante para la fabricación de canoas (PROEXPO, 1972; CONIF, 1996).
Además de su importancia como recurso maderable, C. pyriformis es una especie altamente promisoria en el área de reforestación de bosques nativos, obteniéndose buenos resultados en estudios con formaciones ecológicas de bosque húmedo tropical (Castro et al., 1993).
Diversas entidades nacionales, entre ellas el Instituto SINCHI, se han enfocado en estudiar y mejorar distintos procesos de propagación del Abarco. Respecto de los sistemas de propagación in vitro, hasta la fecha sólo se había reportado el trabajo realizado por Castro et al. (1993) en micropropagación de microestacas provenientes de material ex vitro.
Con el fin de inducir procesos morfogenéticos de novo como embriogenésis somática y organogénesis, se evaluó la respuesta de varios tipos de explantes juveniles procedentes de plántulas establecidas en condiciones in vitro, a distintas combinaciones de fitoreguladores. En el presente artículo se reportan los resultados obtenidos en los procesos de inducción de organogenésis indirecta.Materiales y métodos
Obtención de material fuente y germinación in vitro
Se extrajeron semillas a partir de frutos maduros las cuales, después de lavado con agua corriente, se sometieron a un proceso de desinfección mediante inmersión en polivinil-pirrolidona (Isodine®) al 10% por 1 h y posteriormente en hipoclorito de sodio al 5,25 % por una hora. Después de tres enjuagues con agua destilada estéril, y en condiciones asépticas, se aislaron los embriones que se sembraron, al igual que semillas completas, en medio Woody Plant diluido a la mitad, suplementado con sacarosa al 30% y agar al 6% (Castro et al., 1993). El medio previamente se ajustó a pH 5,8 con HCl 1N y NaOH 1N y se esterilizó en autoclave por 15 min a 20 lbs de presión.
Inducción de callo organogénico
A partir de plántulas de dos semanas de edad obtenidas por germinación in vitro de embriones, se aislaron segmentos de hipocótilos, cotiledones y hojas jóvenes, los cuales se inocularon en el medio anteriormente descrito, suplementado con las distintas combinaciones de fitoreguladores a evaluar (tab. 1).
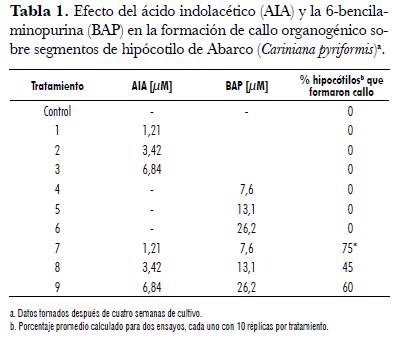
Tomando como referencia el trabajo de Vengadesan et al. (2000) en la especie leñosa Acacia sinuata, se evaluaron tres combinaciones auxina-citoquinina de ácido indolacético (AIA) y 6-bencilaminopurina (BAP) (1,21 y 7,6 µM; 3,42 y 13,1 µM; 6,84 µM y 26,2 µM, respectivamente), al igual que el efecto de cada una de las concentraciones individuales de los fitoreguladores sobre los explantes, estableciendo así un total de 9 tratamientos (Tabla 1). Por cada tratamiento se evaluaron 10 réplicas (segmentos de hipocotilo, cotiledón y hoja, respectivamente), y el ensayo completo se realizó dos veces. En cada tratamiento se evaluó el porcentaje de formación de callo organogénico (callo a partir del cual se formaron brotes caulinares y/o radicales) después de cuatro semanas. Las diferencias estadísticas entre los tratamientos se detectaron utilizando la prueba de los signos (Siecel, 1995) considerando un nivel de significancia a = 0,05.
Los explantes inoculados fueron ubicados en una cámara de crecimiento con un fotoperíodo 12/12 y temperatura de 26 ± 2° C.
Análisis histológicos
Los callos obtenidos fueron fijados en carnoy (3 etanol : 1 ácido acético glacial) por 48 horas, deshidratados a través de inmersión en una serie gradual de concentraciones de alcohol etílico, y por último, bañados en parafina. Se cortaron secciones de 7-10 µm en un micrótomo rotatorio, que se sometieron a tinción mediante la técnica fast green-safranina (Roth, 1964). Posteriormente las láminas fueron observadas al microscopio para evidenciar las estructuras morfogénicas.
Resultados y discusión
Germinación in vitro
En los ensayos realizados, solamente se obtuvo respuesta germinativa en los embriones con porcentajes de germinación entre 80 y 90% (datos no mostrados). Previamente, Castro et al. (1993) intentaron establecer procesos de germinación de embriones in vitro; sin embargo, la alta contaminación y efectos tóxicos de los antibióticos empleados para controlar el crecimiento de los agentes patógenos no les permitieron desarrollar el proceso. En nuestro caso, la contaminación fue muy baja (0-5%, datos no mostrados) y no representó un problema para la germinación y establecimiento del stock in vitro de plantas.
La velocidad del proceso de germinación in vitro fue mayor con respecto al proceso de germinación en suelo, que como lo reportan Diez y Moreno (1998) tiene una duración de 12 a 27 días. En condiciones in vitro, la germinación del embrión comienza a ser notable hacia el quinto día después de la siembra, y aproximadamente 25-30 días después, la plántula se encuentra bien desarrollada con cotiledones completamente expandidos y con las primeras hojas bien formadas. Probablemente, como lo afirma Pierik (1990), debido a la pérdida de inhibidores ocasionada al eliminar la testa, el proceso de germinación se acelera y la plántula se desarrolla rápidamente.
Organogénesis indirecta
De los tres tipos de explantes evaluados, sólo los segmentos de hipocótilo generaron respuesta callogénica. En todos los casos, el callo se originó en los extremos de los explantes y se extendió progresivamente por toda la superficie. En los tratamientos en los que se aplicó únicamente AIA (tratamientos 1-3, Tabla 1), la mayor parte de los explantes inoculados (80%, datos no mostrados) generaron callo compacto de color blanco-amarillo o amarillo, sin ninguna evidencia morfológica ni histológica de posibles estructuras organogénicas. Cuando se aplicó únicamente BAP (tratamientos 4-6, Tabla 1) se formó callo de tipo compacto de color marrón-verde (70% de los explantes inoculados, datos no mostrados), pero igualmente sin evidencias de estructuras organogénicas.
Independientemente de las concentraciones utilizadas, las combinaciones auxina-citoquinina evaluadas (tratamientos 7-9, Tabla 1) originaron callos de coloración marrón-verde de tipo organogénico (figura 1A), en los que se formaron, tanto yemas caulinares como radicales (figura 1B y C).
La combinación AIA (1,21 µM) + BAP (26,2 µM) generó un porcentaje de formación de callo organogénico altamente significativo (75%, p = 0,04122), mientras que las combinaciones AIA (6,84 µM) + BAP (26,2 µM) y AIA (3,42 µM) + BAP (13,1 µM) generaron porcentajes de callo organogénico de 60 y 45%, respectivamente (Tabla 1).
Los análisis histológicos confirmaron la formación de brotes adventicios a partir de los callos organogénicos. La Figura 1D muestra la formación de zonas meristemáticas (domos) y de primordios foliares a partir del tejido parenquimático del callo en el caso de brotes caulinares.
Puesto que la 'recalcitrancia' es una característica común de muchas especies leñosas, el proceso de establecimiento y propagación in vitro de gran parte de ellas se inicia tomando como base explantes juveniles provenientes de líneas germinales debido a su mayor potencial morfogénico (Segura, 1993).
En esta clase de explantes la respuesta callogénica de un tipo particular –como los hipocótilos–, puede indicar, como lo sugieren Sujatha y Mukta (1996), un efecto de predisposición del tejido a sufrir procesos de multiplicación celular a una velocidad mayor que otros, debido a que presenta distinto potencial morfogénico, y/o como sugieren Tonon et al. (2001), como resultado de diferencias en el contenido hormonal del tejido que afectan la interacción con los fitorreguladores exógenos aplicados.
La efectividad de la combinación AIA-BAP en la inducción de brotes adventicios ha sido reportada en varias especies leñosas: Acacia sinuata, Sesbania sesban, Prosopsis cineraria, Dalbergia sissoo, entre otras (Rao, 1988; Vengadesan et al., 2000), evidenciando el papel relevante que juega la interacción específica auxina-citoquinina dentro del proceso de desdiferenciación celular, adquisición de potencial morfogenético y neoformación de estructuras ocurridas en el callo (Street, 1979; De Klerk, 1997, Quoirin et al., 1998).
Recientes estudios moleculares realizados en Arabidopsis, han identificado genes relacionados con la regulación hormonal en los procesos organogénicos. El gen REV/IFL1 (Revoluta/Interfascicular fiberless) por ejemplo, es importante en el transporte polar de auxinas, en la formación de meristemos de brotes y en procesos de diferenciación celular. De otro lado, el gen ABP1 (Proteína 1 de ligamiento a auxina), al parecer está involucrado con la regulación de auxinas en procesos de expansión celular. Adicionalmente, se ha propuesto que elementos reguladores del ciclo celular como las ciclinas-D, juegan un papel relevante en los procesos de multiplicación celular que conllevan al desarrollo de callos, como resultado de su inducción por citoquininas (Mizuka- mi, 2001). Dentro de estos hallazgos, uno de gran interés es la respuesta callogénica inducida por la acción ectópica del gen Ant (Antegumenta) en tejidos diferenciados y sin la adición exógena de auxinas o citoquininas, lo que sugiere un papel primordial de este gen en el mantenimiento de la 'competencia meristemática', es decir, la capacidad celular de crecimiento y multiplicación que surge como respuesta a señales de crecimiento y desarrollo durante los procesos organogénicos vegetales (Mizukami, 2001).
La formación de un tipo determinado de estructura (brotes caulinares o radicales) es dirigida en gran parte por la proporción auxina/citoquinina aplicada (Har- tmann et al., 1997). Sin embargo, en nuestro estudio, la formación conjunta de brotes caulinares y radicales en el callo no nos permitió establecer un requerimiento específico de esta proporción para direccionar el desa- rrollo de uno u otro tipo.
Siguiendo el proceso realizado por Vengadesan et al. (2000), algunos callos se inocularon en medio suplemen- tado con AG3 (datos no mostrados), con el fin de inducir la elongación de los brotes caulinares; sin embargo, los fuertes procesos oxidativos presentados a lo largo de la primera semana después del cambio de medio, llevaron a su necrosis y pérdida total.
La oxidación es uno de los mayores problemas encon- trados durante el desarrollo de procesos in vitro con espe- cies leñosas. No obstante, en muchos casos se ha podido optimizados con éxito (George, 1993). Es necesario para el caso de abarco, enfocar esfuerzos en la búsqueda de tratamientos que solucionen el problema oxidativo, de manera que se pueda continuar y mejorar el desarrollo de un sistema de multiplicación por brotación adventicia. La formación de brotes adventicios, y su posterior desarrollo a plantas, constituye un sistema con altas tasas de multiplicación (Jones, 1983; Hartmann et al., 1997); y ha sido utilizado con éxito en la propagación de especies leñosas de alta importancia económica como Citrus sp. y palma de aceite (Jones, 1983; George, 1993). En el caso del abarco, la optimización de este tipo de sistema y de otros procesos de propagación in vitro, probablemente requieran de una ardua investigación. Sin embargo, por tratarse de una especie nativa de alto potencial económico y ambiental para el país, es importante continuar explorando todas las vías posibles para su multiplicación y futuro mejoramiento, paralelamente a investigaciones sobre su genética, fisiología y ecología, con el fin de generar el conocimiento necesario para aprovechar al máximo sus beneficios.
Conclusión
Sí bien el sistema de brotación adventicia en abarco necesita ser optimizado, la inducción de brotes lograda mediante la adición conjunta de AIA y BAP constituye un primer paso en el desarrollo de éste y otros procesos in vitro, que en un futuro contribuyan en los programas de propagación y mejoramiento de esta especie maderable nativa.
Agradecimientos
Los autores agradecen a la Gobernación del Guaviare y al Plan Nacional de Desarrollo Alternativo (PNDA) por la financiación brindada, así como al Instituto Amazónico de Investigaciones Científicas SINCHI, por facilitar los elementos necesarios para el desarrollo de este trabajo.
Literatura citada
Bernal, H. y J. Correa. 1994. Especies vegetales promisorias de los países del Convenio Andrés Bello. Tomo X. Editora Guadalupe, Bogotá. 325 p. [ Links ]
Castro, D.; C. Jiménez; D. Ríos; A. Restrepo y M. Giraldo. 1993. Utilización de las técnicas de cultivo de tejidos vegetales in vitro para la propagación y conservación de germoplasma de cuatro especies vegetales en vía de extinción en el oriente antioqueño: Comino (Aniba perutilis), abarco (Cariniana pyriformis), almedrón (Caryocar glabrum) y guayacán (Tabebura serratifolia). pp. 21-44. En: Cuadernos de Investigación y Desarrollo Regional CORNARE. El Santuario, Antioquia. 94 p. [ Links ]
Corporación Nacional de Investigación y Fomento Forestal (CONIF). 1996. Latifoliadas de zona baja. CONIF, Bogotá. 104 p. Díez, A. y M. Moreno. 1998. árboles de los bosques húmedos tropicales del sur oriente de Antioquia, Colombia. Revista Facultad Nacional de Agronomía Medellín 5(2), 9-25. [ Links ] [ Links ]
De Klerk, G.; B. Arnholdt-Schmitt; R. Lieberei y K. Newmann. 1997. Regeneration of roots, shoots and embryos: physiological, biochemical and molecular aspects. Biologia Plantarum 39(1), 53-66. [ Links ]
Fondo de Promoción de Exportaciones (PROEXPO), Colombia. 1972. Maderas colombianas. Editorial Itualgraf, Bogotá. 117 p. [ Links ]
George, E. 1993. Plant propagation by tissue culture: the technology. Segunda edición. Editorial Hardcover, New York. 574 p. [ Links ]
Hartmann, H.; D. Kester; F. Davies y R. Genere. 1997. Plant propagation, principles and practices. Sixth edition. Ed. Prentice Hall, New York. 770 p. [ Links ]
Jones, O. 1983. In vitro propagation of tree crops. pp. 139-159. En: Miantell, S. y H. Suiter (eds.). Plant biotechnology. Cambridge University Press, Cambridge. 582 p. [ Links ]
Mizukami, Y. 2001. A matter of size: developmental control of organ size in plants. Current Opinion in Plant Biology 4, 533-539. [ Links ]
Morton, C.; S. Mori; G. Prance; K. Karol y M. Chase. 1997. Phylogenetic relationships of Lecythidaceae: A cladistic análisis using RBCL sequence and morphological data. American Journal of Botany 84(4), 530-540. [ Links ]
Pierik, R. 1990. Cultivo in vitro de las plantas superiores. Editorial Mundi-Prensa, Madrid. 326 p. [ Links ]
Quoirin, M.; J. Bittencourt; F. Zanette y D. de Oliveira. 1998. Effect of growth regulators on indirect organogenesis of Acacia mearnsii tissues cultured in vitro. Revista Brasileira de Fisiologia Vegetal 10(2), 101-105. [ Links ]
Rao, A. 1988. In vitro culture studies on economically important tropical trees. pp. 9-195. En: The application of tissue culture techniques in economically important tropical trees. Biotrop. Special Publication N° 35. Editorial SEAMEO-BIOTROP. Indonesia. 229 p. [ Links ]
Roth, I. 1964. Microtécnica vegetal. Editorial Escuela de Biología, Facultad de Ciencias, Universidad Central de Venezuela, Caracas. 132 p. [ Links ]
Segura, J. 1993. Morfogénesis in vitro. pp. 240-325. En: Azcón-Bieto J. y M. Talón (eds.). Fisiología y bioquímica vegetal. Editorial McGraw Hill-Interamericana, Madrid. 581 p. [ Links ]
Siecel, S. 1995. Estadística no paramétrica. Ed. Trillos, Barcelona. 250 p. [ Links ]
Street, H.E. 1979. Embryogenesis y chemically induced organogenesis. pp. 123-153. En: Sharp, W.; P. Larsen; E. Paddock y V. Raghavan (eds). Plant cell and tissue culture, principles and applications. University Press, Columbus, Ohio. 650 p. [ Links ]
Sujatha, M. y N. Mukta. 1996. Morphogenesis and plant regeneration from tissue cultures of Jatropha curcas. Plant Cell, Tissue and Organ Culture 44, 135-141. [ Links ]
Tonon, G.; M. Capuana y A. Di Marco. 2001. Plant regeneration of Fraxinus angustifolia by in vitro shoot organogenesis. Scientia Horticulturae 87, 291-301. [ Links ]
Vengadesan, G.; A. Ganapathi; R. Premanand y V. Ramesh. 2000. In vitro organogenesis and plant formation in Acacia sinuata. Plant Cell, Tissue and Organ Culture 61, 23-28. [ Links ]

