










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.23 n.1 Bogotá jan./jul. 2005
Caracterización palinológica y viabilidad polínica de Physalis peruviana L. y Physalis philadelphica Lam.
Physalis peruviana L. and Physalis philadelphica Lam. palynological characterization and pollen viability
Tulio César Lagos1,Creuci María Caetano2,Franco Alirio Vallejo3Jaime Eduardo Muñoz4,Hernando Criollo5y Cristián Olaya 6
1Profesor Asistente, Facultad de Ciencias Agrícolas, Universidad de Nariño, Pasto. e-mail: tclagos3@yahoo.com2Consultora IPGRI Américas. e-mail: mrhcaetano@yahoo.com.br
3Profesor Titular, Facultad de Agronomía, Universidad Nacional de Colombia, Palmira.
4Profesor Asociado, Facultad de Agronomía, Universidad de Colombia, Palmira.
5Profesor Asociado, Facultad de Ciencias Agrícolas, Universidad de Nariño, Pasto. e-mail: hcriollo@hotmail.com 6Unidad de microscopia electrónica, Centro Internacional de Agricultura Tropical (CIAT), Cali.
Fecha de recepción: 2 de febrero de 2005. Aceptado para publicación: 27 de mayo de 2005.
Resumen
Physalis peruviana L. y P. philadelphica Lam. son dos solanáceas de importancia económica para Colombia y México. Existen grandes vacíos en el conocimiento de la palinología y viabilidad polínica de estas dos especies. Por lo tanto, el objetivo del presente trabajo fue caracterizar su grano de polen y determinar su viabilidad bajo condiciones contrastantes de climas frío y cálido. La caracterización del polen se hizo a través de la técnica de acetólisis (en microscopía de luz) y por microscopía electrónica de barrido. Se determinaron variables como el eje polar, el diámetro ecuatorial, el número, posición y carácter de las aberturas (NPC), lado de apocolpo/apoporo, forma polínica, contorno en vista polar y en vista ecuatorial, tipos de área polar y de abertura, y el tipo polínico. Estas variables permitieron describir y caracterizar el grano de polen de los genotipos estudiados. Con el empleo de técnicas de fijación de flores en antesis y tinción con acetocarmín, se hizo un análisis de la viabilidad polínica bajo microscopía de luz. Las dos especies presentan un grano de polen zono-tricolporado, de tamaño mediano en P. peruviana y pequeño en P. philadelphica. Los ambientes marginales reducen la viabilidad del polen pero no afectan su fertilidad.
Palabras clave adicionales: Solanáceas, morfología polínica, tinción, acetólisis, microscopía de luz.
Abstract
Physalis peruviana L. and P. philadelphica Lam. represent two Solanaceae species which are economically important for Colombia and Mexico, respectively. Little is currently known regarding P. peruviana and P. philadelphica palynology and pollen viability. This work was thus aimed at characterising these two species' pollen grains and determining pollen viability in contrasting cold and warm climate conditions. Acetolysis (light microscopy) and scanning electron microscopy were employed for pollen characterisation. The following traits were determined: polar axis, equatorial diameter, aperture number, position and character (NPC), apocolpus/apoporus side, pollen morphology, axis and equatorial contour, polar area and aperture and pollen types. This led to describing and characterising the pollen grains of the genotypes being studied. Flower fixation during anthesis, followed by acetocarmin staining, was used for establishing pollen viability; observations were made by light microscope. The two species displayed a medium-sized pollen grain zono-tricolporate in P. peruviana and a small-sized one in P. philadelphica. Marginal environments reduced pollen viability but did not affect fertility.
Additional key words: Solanaceae, pollen morphology, staining, acetolysis, light microscopy.
Introducción
EL GéNERO PHYSALIS fue establecido por Linneo en 1753. Comprende aproximadamente 100 especies entre herbáceas anuales y perennes. Cinco de éstas se cultivan por su fruto: P. peruviana y P. pruinosa L. se consumen en fresco y en mermeladas; P. ixocarpa Brot. y P. philadelphica se utilizan como hortalizas, mientras que P. alkekengi L. es ornamental. En algunos casos, se ha ubicado a P. philadelphica bajo la sinonimia de P. ixocarpa, pero son distintas. P. peruviana y P. philadelphica han sido señaladas como de importancia económica para Colombia y México. Sin embargo, los estudios básicos para el mejoramiento y el manejo de los recursos genéticos de las dos especies, principalmente para P. peruviana, son escasos (Menzel, 1951; Moriconi et al., 1997 y McCain, 1993).
Existen grandes vacíos respecto a la viabilidad y caracterización del grano de polen de P. peruviana y P. philadelphica. La cantidad y calidad del polen producido por una flor es uno de los aspectos más importantes en estudios evolutivos y en la adaptación de los genotipos. Por lo tanto, es necesario establecer la viabilidad y caracterizar el polen en las dos especies.
El grano de polen es la fase discreta y móvil del gametofito masculino de las plantas superiores. En un sentido amplio, se considera al proceso de polinización como la transmisión del grano de polen al gametofito femenino. A su vez, la germinación y crecimiento del gametofito masculino hace parte de la fertilización o fecundación. Los mecanismos de polinización y fecundación tienen una relación decisiva sobre los procesos de fitomejoramiento y producción de cultivos (Frankel y Galun, 1977).
Las fallas en la formación y desarrollo del polen son una causa primaria de inviabilidad de especies o de híbridos. En este caso, la inviabilidad del polen puede deberse a gametos no reducidos (de tamaño más grande), o que no tienen un número cromosómico balanceado (de menor tamaño). Las causas no genéticas que afectan negativamente al polen incluyen la temperatura, la humedad, la fertilidad del suelo, las enfermedades, las plagas y la edad (Delph et al., 1997; Caetano et al., 2003). El estudio del polen recibe el nombre de palinología (del griego palynein que significa dispersar). Erdtman (1952) indica que la palinología es una ciencia que estudia las paredes de las esporas y los granos de polen sin tener en cuenta su contenido protoplásmico.
Dentro de la caracterización morfológica, el estudio y el análisis de los descriptores relacionados con el polen son vitales para diferenciar y relacionar especies, debido a que la forma y la ornamentación de la exina son características genéticamente determinadas y, por tanto, muy conservadas (Sasakuma y Yamada, 1988).
La gran diversidad de formas, tamaños, aperturas, caracteres estructurales y esculturales de la exina es una manifestación de la adaptación, que asegura que el polen pueda llegar hasta el estigma de la flor de la misma especie por varios vectores como la gravedad, el viento, el agua, y los insectos, entre otros (Erdtman, 1986; Uribe y Fonnegra, 1979).
En muchos casos el tipo de polen de un taxón es característico y constante, siendo denominado 'estenopalinológico'. En otros taxones, los tipos de polen varían considerablemente en abertura, escultura, estratificación de la exina, tamaño o forma. éstos conforman el taxón 'euripalinológico'. Los grupos europalinológicos son los más frecuentes, por tanto, de gran importancia para la clasificación taxonómica y sistemática, como también para caracterizar los recursos genéticos de un género o de una especie (Fonnegra, 1989).
Según Erdtman (1986) los granos de polen de la familia Solanaceae, en donde se ubica el género Physalis, son (2-)3-5(-6)-colpado, -colporoide, -colporado, algunas veces inaperturado y oblato-prolato. La longitud de los ejes oscila entre 14,5 µm (Solanum dulcamara) y 61 µm (Nierembergia frutescens).
El trabajo se llevó a cabo con el objeto de caracterizar el grano de polen de P. peruviana y P. philadelphica, y determinar la viabilidad polínica bajo condiciones contrastantes de clima frío y cálido de las dos especies.
Materiales y métodos
Descripción del material experimental
Para la caracterización palinológica y la viabilidad del polen (porcentaje de granos de polen viables) se utilizaron las accesiones UN-40 (Nariño), UN-49 (Nariño), UNPU054 (Felidia, Valle), UNPU099 (Silvania, Cundinamarca) y Kenya (Bogotá) de P. peruviana y Tepehuan (Estados Unidos) de P. philadelphica. Las principales características de estos genotipos son:
UN-40. Forma silvestre colectada en el municipio de Potosí (Nariño) a 2.669 m.s.n.m. Presenta una ramificación basal dicotómica y un hábito de crecimiento erguido. La pubescencia es escasa en el tallo y abundante en las hojas. Según la Norma ICONTEC (1999), presenta frutos calibre C con un diámetro de 20,05 mm. Esta norma clasifica los frutos de P. peruviana en calibres A (=15 mm de diámetro), B (15,1-18 mm), C (18,1-20 mm), D (20,1-22 mm) y E (>23 mm). Su rendimiento promedio es de 581 g/planta y semana. La pulpa del fruto maduro (estado 6 de maduración en la norma citada) contiene 13,6° Brix, 1,58% de ácido cítrico (AC) y pH de 4,6 (Hejeile e Ibarra, 2001; Criollo et al., 2001).
UN-49. Colectada en el municipio de Yacuanquer (Nariño) a 2.755 m.s.n.m., cultivada en huerto. Su ramificación es basal dicotómica con hábito de crecimiento erguido. La pubescencia es abundante, tanto en el tallo como en las hojas. Presenta frutos de calibre E con un diámetro de 25,64 mm. Su rendimiento promedio es de 587 g/planta y semana. La pulpa del fruto estado 6 contiene 14,2° Brix, pH de 4,6 y 1,54% de AC (Hejeile e Ibarra, 2001; Criollo et al., 2001). UNPU054. Colectada en la vereda Felidia (Valle del Cauca) a 1.720 m.s.n.m. Se encontró como una forma espontánea en jardín. Sus frutos son de calibre A con un diámetro de 14 mm. El peso del fruto oscila entre 2,4 y 3,8 g; su pulpa contiene entre 12 y 14° Brix (Bonilla y Espinosa, 2003).
UNPU099. Cultivar comercial colombiano proveniente de Silvania (Cundinamarca). El peso promedio del fruto es de 4 a 5 g. Los frutos presentan una mejor coloración y un mayor contenido de azúcares, cualidades que los hacen apetecibles en el mercado.
Kenya. Se introdujo a finales de los años ochenta al departamento de Boyacá (Almanza y Fischer, 1993). Es una planta de porte bajo, con hojas grandes. El fruto tiene un peso de 6 a 10 g, con menos contenido de azúcares que el genotipo comercial de Granada y Silvania.P. philadelphica var. Tepehuan. Tallo herbáceo, hoja oval-lanceolada, flores solitarias con presencia de lóbulos en la corola de los cuales cinco tienen una longitud menor que la mitad del largo de la corola. El cáliz es pubescente, con diez aristas longitudinales y un aspecto inflado a la madurez del fruto. El diámetro del fruto oscila entre 4 a 8 cm. Las semillas son redondeadas (Rodríguez y Azurdia, 1996).
Localización
Las accesiones de P. peruviana se sembraron en la Granja Experimental Botana de la Universidad de Nariño y en el Centro Experimental de la Universidad Nacional de Colombia, sede Palmira (CEUNP). Botana está localizada a 2.820 m.s.n.m., 01°09'12" latitud N y 77°18'31" longitud O, en el Altiplano de Pasto (Nariño), con una temperatura promedio de 13°C y precipitación promedia anual de 840 mm (Lagos et al., 2001). CEUNP está ubicado a 927 m.s.n.m., 2° 06' latitud N y 65° 03' longitud O, en el corregimiento El Carmelo, municipio de Candelaria (Valle). La precipitación promedio anual es de 1.100 mm, la temperatura promedia de 24°C y humedad relativa de 70% (Gutiérrez, 1997). El análisis de las muestras se realizó en el laboratorio de microscopía de la Unidad de Virología del Centro Internacional de Agricultura Tropical (CIAT), localizado en Palmira, Valle del Cauca.
Caracterización y viabilidad del grano de polen
En la caracterización del grano de polen se emplearon los genotipos UN-49 y Kenya de P. peruviana, y Tepehuan de P. philadelphica. Al polen se le tomaron fotografías a través del microscopio electrónico de barrido (MEB). Igualmente, se utilizó la técnica de acetólisis (Erdtman, 1952) para la observación bajo el microscopio de luz, en donde se evaluaron diez campos por placa, que correspondieron a cincuenta observaciones por accesión. La viabilidad polínica se determinó en todos los genotipos. De cada accesión se tomaron cinco flores en antesis y con cada flor se preparó una placa según la metodología convencional. La tinción se hizo con acetocarmín al 2%. Se observó en un microscopio de luz marca Aristoplan® con el objetivo de 25/0,65. En cada una de las placas se realizó el conteo de los granos de polen viables (teñidos de rojo) y no viables (incoloros).
Variables evaluadas
Con base en las fotografías del MEB y en las observaciones de polen acetolizado bajo microscopía de luz, se determinó el eje polar (P), el diámetro ecuatorial (E), E en vista polar (EP), y la relación entre P y E (P/E). El eje P corresponde la longitud del polen de un polo a otro en el plano ecuatorial. El diámetro E es la longitud de la línea ecuatorial del grano de polen.
Se describieron las características estructurales de la exina, la forma del grano de polen, el número de aberturas y la distancia promedia entre aberturas. Todas las mediciones se hicieron en micrómetros (µm). Además, se estableció el índice de área polar (IAP) y los tipos de área polar y de abertura según el IAP (relación entre la distancia entre dos aberturas y el diámetro ecuatorial en vista polar).
Análisis de la información
Para el análisis de los descriptores de caracterización, se utilizaron las medidas de tendencia central y de dispersión. Los datos sobre viabilidad polínica se sometieron al análisis de varianza (ANDEVA) y a la prueba de significancia de medias (DMS).
Resultados y discusión
Caracterización palinológica de P. peruviana y P. philadelphica Bajo la MEB, el polen P. peruviana y de P. philadelphica se presenta como una mónada isopolar con simetría radial. Las accesiones UNPU099, UN-49, Kenya y Tepehuan presentan polen tricolporado, zonoaperturado (figura 1), aunque en el material Kenya se pueden encontrar granos tetracolporados. Esto se debe a la presencia de microsporas no reducidas o con mayor número de cromosomas.
Acorde con las accesiones estudiadas, la morfología polínica del género Physalis corresponde a un taxón estenopalinológico, monomórfico excepto para la accesión Kenya, similar a lo observado por Benítez de Rojas y Magallanes (1998). Lo mismo es válido también para P. angulata L., P. cordata Millar. Diet., P. lagascae R. et S., P. pubescens L. y P. peruviana. Estas especies se distribuyen en Venezuela principalmente como vegetación adventicia, producto de la intervención antrópica, con amplitud de distribución en cuanto a niveles de temperatura y pisos altitudinales (Benítez de Rojas y Magallanes, 1998).
La ornamentación de la exina en vista superficial es granulosa y escabrosa para P. philadelphica, mientras que es granulosa y equinada (erizada o espinosa) en P. peruviana (Figura 1). Contrariamente, Benítez de Rojas y Magallanes (1998) indican que la esculturación de la exina es de tipo verrugoso para las especies P. angulata, P. cordata, P. lagascae, P. pubescens y P. peruviana.
Las formas polínicas, según el índice P/E para P. peruviana y P. philadelphica, corresponden a prolato-esferoidal y prolato, en su orden. El contorno en vista polar es circular para UN-49 y angular-triangular-agudo-convexo para Kenya. En su vista ecuatorial es rómbico para ambas. El polen de P. philadelphica es angular-triangular-obtuso-convexo en el ámbito polar y elíptico en su contorno ecuatorial (tab. 1). Similarmente, Benítez de Rojas y Magallanes (1998) reportan que el polen de las cinco especies ya mencionadas, en su vista polar, es triangular-obtuso-convexo y elíptico-acuminado-obtuso. En la tab. 2 se muestran las medidas del eje polar (P) y del diámetro ecuatorial (E) en polen acetolizado de las accesiones UN-49 y Kenya de P. peruviana y Tepehuan de P. philadelphica. En general, el polen de P. peruviana es de mayor tamaño que el P. philadelphica.
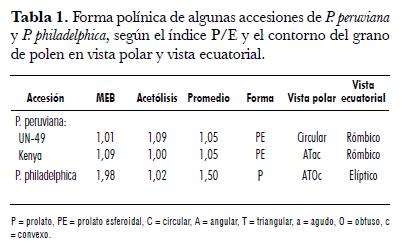
El P para la primera es de 31,6 a 33,03 µm y para la segunda de 21,3 µm; el E es de 30 a 31,7 µm y 20,8 µm, en su orden. En las accesiones de P. peruviana, el P oscila entre 27 y 37,8 µm, presentando un tipo polínico mediano. En cambio, el tipo polínico de P. philadelphica es pequeño, debido a que su rango de P es de 16,2 a 24,3 µm (tab. 2).En la tab. 2se muestran las medidas del eje polar (P) y del diámetro ecuatorial (E) en polen acetolizado de las accesiones UN-49 y Kenya de P. peruviana y Tepehuan de P. philadelphica. En general, el polen de P. peruviana es de mayor tamaño que el P. philadelphica. El P para la primera es de 31,6 a 33,03 µm y para la segunda de 21,3 µm; el E es de 30 a 31,7 µm y 20,8 µm, en su orden. En las accesiones de P. peruviana, el P oscila entre 27 y 37,8 µm, presentando un tipo polínico mediano. En cambio, el tipo polínico de P. philadelphica es pequeño, debido a que su rango de P es de 16,2 a 24,3 µm (tab. 2).
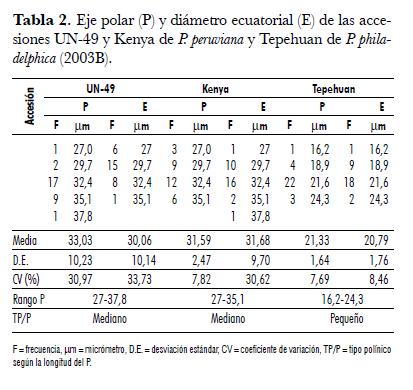
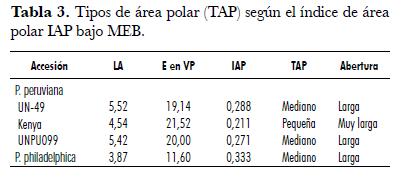
Los valores para P. peruviana son mayores a los obtenidos por Benítez y Magallanes (1998), quienes encontraron un P de 17,70 µm y un E de 11,83 µm. Esto indica que dentro de la especie P. peruviana puede existir cierta variabilidad genética relacionada con la distribución geográfica. Por otro lado, el tipo de área polar (TAP) va desde pequeña hasta mediana, con aberturas largas a muy largas en P. peruviana, y mediana con abertura larga para P. philadelphica (tab. 3).
Viabilidad polínica de P. peruviana L. y P. philadelphica Lam.
Los porcentajes de viabilidad polínica para P. peruviana y P. philadelphilca, obtenidos bajo condiciones del Altiplano de Pasto (Botana, Nariño) y CEUNP (Candelaria, Valle del Cauca) fueron estadísticamente diferentes entre las dos localidades y entre las accesiones a través de las dos localidades. La interacción genotipo por ambiente fue significativa; por lo tanto, existe un comportamiento diferencial de cada genotipo en cada una de las localidades (tab. 4).
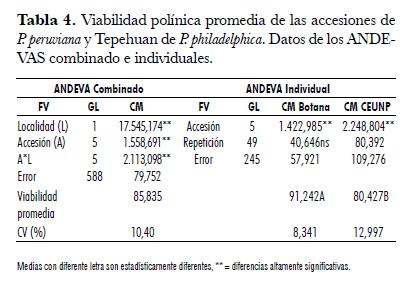
El mayor porcentaje de viabilidad se obtuvo en Botana con un 91,24%. Esta localidad presenta diferencias altamente significativas con CEUNP (80,43%). A pesar de la menor viabilidad observada en CEUNP, los rangos obtenidos se consideran normales, puesto que no afectan la fertilidad de la especie. Dentro de cada localidad, se obtuvieron diferencias altamente significativas entre las accesiones (tab. 4).
En Botana, los porcentajes de viabilidad oscilaron entre 81,7 y 98%. UN-49 presentó el máximo promedio de viabilidad con diferencias altamente significativas respecto a las demás accesiones (tab. 5). Le siguen en su orden UNPU054 (93,5%), UN-40 (92,2%), UNPU099 (91%) y Kenya (91%), las cuales no presentaron diferencias significativas entre ellas. Tepehuan, con 81,7% de viabilidad, fue la de menor promedio. Los altos porcentajes de viabilidad presentados por las accesiones de P. peruviana en Botana se deben a su alto grado de adaptación para las condiciones ambientales de esta localidad. Con respecto al rango de adaptación, la especie crece en zonas altas desde Chile hasta Venezuela como planta silvestre y semisilvestre, entre los 1.500 y los 3.000 msnm (Fischer, 2000 y Zapata et al., 2002).
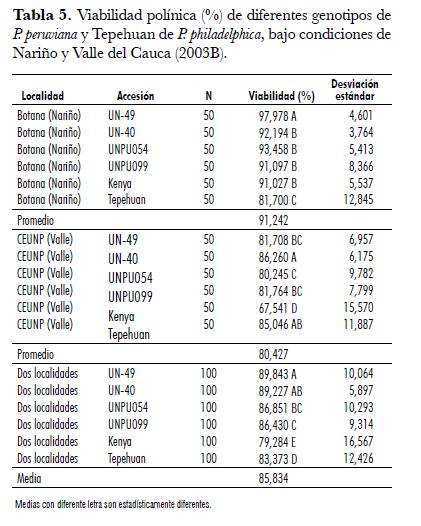
El porcentaje de viabilidad polínica obtenido en CEUNP fue de 80,4 % (tab. 5). A pesar de que los valores obtenidos son bajos, no afectan la fertilidad de la especie. Puesto que el crecimiento y desarrollo de las plantas fue normal y la viabilidad del polen no fue afectada por las condiciones ambientales extremas del trópico seco (altitud de 1.000 m.s.n.m., temperatura promedia de 24°C y una humedad relativa del 70%), es probable que el rango de adaptación de P. peruviana puede ser más amplio. Es posible también que en regiones como la zona cafetera existan ambientes potenciales para desarrollar cultivos comerciales, lo que requiere previa evaluación y selección de genotipos adecuados. Todos los genotipos presentaron menor viabilidad en CEUNP, variando de 67,5% para Kenya hasta 86,26% para UN-40 (tab. 5). Esto se puede explicar por una respuesta de adaptación a condiciones ambientales extremas, como alta temperatura y baja humedad relativa, conforme lo explican Delph et al. (1997) y Caetano et al. (2003).
Al igual que P. ixocarpa, P. philadelphica se encuentra en forma silvestre en la Costa del Pacífico, desde California hasta Centro América (Peña y Santiaguillo, 1999). A pesar de su origen (regiones templadas y semitempladas), el porcentaje de viabilidad de Tepehuan no fue afectado por las condiciones ambientales del trópico de altura de Botana (Nariño). Esto permite ubicarla como un componente del pool genético primario de P. peruviana, para efectos de utilizarla en programas de mejoramiento, como una fuente opcional de variabilidad genética. Al respecto, California Rare Fruit Growers (CRFG, 2003) indica que las especies relacionadas de P. peruviana son: Physalis heterophylla, P. ixocarpa, P. philadel phica, P. pruinosa, P. pubescens y P. viscosa. Se conocen los híbridos entre estas especies.
En comparación con Botana, Tepehuan obtuvo una mayor viabilidad polínica en CEUNP (85,05%), en donde no presentó diferencias significativas con el promedio más alto obtenido por UN-40 (86,26%). Además, superó significativamente a las otras accesiones de P. peruviana. Con base en los resultados obtenidos, se puede establecer que P. philadelphica puede adaptarse más fácilmente a condiciones de trópico seco que a las condiciones de trópico de altura. Conclusiones Las dos especies poseen un grano de polen zono-tricolporado, aunque en el cultivar Kenya se pueden encontrar polen tetracolporado. El polen es mediano en P. peruviana y pequeño en P. philadelphica. Las formas polínicas corresponden a prolato-esferoidal en P. peruviana y prolato en P. philadelphica. Existen pocas variaciones en la ornamentación de la exina como en la forma en su vista polar y ecuatorial.
Los genotipos evaluados de P. peruviana y de P. philadelphica presentaron una alta viabilidad de polen. Ambientes marginales como el del Valle del Cauca reducen esta viabilidad, sin alterar la fertilidad de las especies.
Literatura citada
Almanza, P. y G. Fischer. 1993. Nuevas tecnologías de la uchuva Physalis peruviana L. Agro-Desarrollo 4(1-2), 292-304. [ Links ]
Benítez de Rojas, C. y A. Magallanes. 1998. El género Physalis (Solanaceae) de Venezuela. Acta Botánica 21(2), 11-42. [ Links ]
Bonilla, M. y K. Espinosa. 2003. Colección, caracterización fenotípica, molecular de poblaciones de uchuva Physalis peruviana L. Trabajo de grado. Facultad de Agronomía. Palmira, Universidad Nacional de Colombia, Palmira. 94 p. [ Links ]
Caetano, C.M.; G. Coppens; N.M. Stenzel; C.A. Olaya; J.A. Arroyave; J.A. Campo; D.G.C. Nunes; B.R.C. Nunes y J. Vega. 2003.Estimativa da viabilidade polínica em quatro subgéneros de Passiflora: eficiencia de diferentes métodos. Arquivos da Apadec 7(Supl.), 26-27. [ Links ]
Criollo, H.; T.C. Lagos; P. Criollo y M. Guerrero. 2001. Caracterización de materiales de uvilla o uchuva (Physalis peruviana L.) por sus características de calidad. Revista de Ciencias Agrícolas 18(2),168-180. [ Links ]
CRFG. California Rare Fruit Growers. 1997. Cape gooseberry: Physalis peruviana L. En: http://www.crfg.org/pubs/ff/cape-gooseberry.html Consulta: diciembre, 2004. [ Links ]
Delph, L.F.; M.H. Johannsson y A.G. Stephenson. 1997. How environmental factors affect pollen performance: ecological and evolutionary perspectives. Ecology 78(6), 1623-1639. [ Links ]
Erdtman, G. 1952. Pollen morphology and plant taxonomy: angiosperms. Almqvist and Wiksall, Estocolmo. 450 p. [ Links ]
Erdtman, G. 1986. Pollen morphology and plant taxonomy: angiosperms: An introduction to palynology. Leiden, New York. 553 p. [ Links ]
Fischer. G. 2000. Crecimiento y desarrollo. En: Flórez V.J.; G. Fischer y A.D. Sora (eds.). Producción, poscosecha y exportación de la Uchuva (Physalis peruviana L.). Unibiblos, Universidad Nacional de Colombia, Bogotá. 175 p. [ Links ]
Fonnegra, C. 1989. Introducción a la palinología. Métodos de estudio palinológico. Medellín, Universidad de Antioquia. pp. 11-43. [ Links ]
Frankel, R. y E. Galun. 1977. Pollination mechanisms, reproduction and plant breeding. Springer-Verlag, New York. 281 p. [ Links ]
Gutierrez, J.A. (ed). 1997. Programa de investigación en hortalizas: semillas UNAPAL. Universidad Nacional de Colombia, Palmira. 62 p. [ Links ]
Hejeile, H. y A. Ibarra. 2001. Colección y caracterización de recursos genéticos (Physalis peruviana L.) en algunos municipios del sur del departamento de Nariño. Trabajo de grado. Facultad de Ciencias Agrícolas, Universidad de Nariño, Pasto. 123 p. [ Links ]
ICONTEC. 1999. Norma técnica Colombiana NTC 4580: Frutas frescas: uchuva. Instituto Colombiano de Normas Técnicas y Certificación. Bogotá. 15 p. [ Links ]
Lagos, T.C.; H. Criollo y C. Mosquera. 2001. Evaluación preliminar de cultivares de uvilla o uchuva (Physalis peruviana L.) para escoger materiales con base en la calidad del fruto. Revista de Ciencias Agrícolas 18(2), 82-94. [ Links ]
McCain, R. 1993. Goldenberry, passionfruit & white sapote: Potential fruits for cool subtropical areas. pp. 479-486. En: Janick, J. y J.E. Simon (eds.). New crops. Wiley, New York. [ Links ]
Menzel, M. 1951. The cytotaxonomy and genetics of Physalis. Proceedings of the American Philosophical Society 65(2), 132-183. [ Links ]
Moriconi, D.N. Rush, M.C. y H. Flores. 1990. Tomatillo: A potential vegetable crop for Louisiana. pp. 407-413. En: Janick, J. y J.E. Simon (eds.). Advances in new crops. Timber Press, Portland, OR. En: http://www.hort.purdue.edu/newcrop/proceedings1990/v1-407.html#Fig. 1; consulta: diciembre, 2004. [ Links ]
Peña, A. y F. Santiaguillo. 1999. Variabilidad genética de tomate de cáscara en México. Boletín Técnico No. 2. Programa Nacional de Investigación en Olericultura, departamento Fitotecnia, Universidad Autónoma Chapingo, México. 26 p. [ Links ]
Rodríguez, F. y C. Azurdia. 1996. Distribución geográfica y características agromorfológicas de los cultivares de Miltomate (Physalis spp.) de Guatemala. En: Azurdia, C. (ed.) Lecturas en recursos fitogenéticos. Instituto de Investigaciones Agronómicas, Facultad de Agronomía, Universidad de San Carlos, Guatemala. pp. 72-73. [ Links ]
Sasakuma, I.Y. y Y. Yamada. 1988. Pollen: illustrations and scanning of electronmicrographs. Kodansha, Tokyo and Springer-Verlag. 198 p. [ Links ]
Uribe, F. y R. Fonnegra. 1979. Importancia de la forma, estructura y tamaño del polen en la determinación de la familia Melastomataceae. Actual. Biol. 8(27-28), 3-9. [ Links ]
Zapata, J.L; A. Saldarriaga; M. Londoño y C. Díaz. 2002. Manejo del cultivo de la uchuva en Colombia. Boletín Técnico 14. CORPOICA, PRONATTA, Rionegro, Antioquia. 40 p. [ Links ]

