










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.23 no.2 Bogotá July/Dez. 2005
FITOMEJORAMIENTO, RECURSOS GENÉTICOS Y BIOLOGÍA MOLECULAR
Evaluación de la habilidad combinatoria y heterosis de siete progenitores de arveja Pisum sativum L.
Evaluating combinatory ability and heterosis of seven parental Pisum sativum L.
Natalia Espinosa1 y Gustavo Adolfo Ligarreto2
1 Ingeniera agrónoma, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. e-mail: ebayer@yahoo.com
2 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. e-mail: aligarretom@unal.edu.co
Fecha de recepción: 6 de abril de 2005 Aceptado para publicación: 21 de noviembre de 2005
RESÚMEN
En Colombia se carece de información referente al tipo de control genético en la arveja que gobierna la precocidad, el rendimiento y sus componentes. El objetivo de este trabajo fue evaluar la habilidad combinatoria y heterosis en cuatro líneas y tres variedades comerciales de arveja, para once variables agronómicas. Se desarrolló un diseño dialélico (Griffing, método 1, modelo 1) con siete materiales, bajo condiciones de la Sabana de Bogotá. Se hizo el análisis de varianza y se obtuvieron los valores estimados para habilidad combinatoria general (HCG) y específica (HCE), y se calcularon los valores de heterosis sobre la media parental (HMP) y sobre el mejor padre (HBP). La HCG fue altamente significativa para ocho de los caracteres evaluados y la HCE fue significativa para: nudos hasta la primera flor, longitud de vaina, número de semillas por vaina y peso de 100 semillas. Las variedades 'Pequinegra', 'Santa Isabel' e 'ICA Tominé' fueron los mejores combinadores generales. La variedad 'Pequinegra' tuvo efectos recíprocos altamente significativos en caracteres asociados con precocidad. En cuanto a la evaluación de heterosis, los híbridos F1 tuvieron un mejor comportamiento que la media de sus parentales en cinco de los componentes de rendimiento. Aún cuando sólo tres de las progenies en estudio fueron más precoces que el mejor padre, para la HMP se hallaron valores negativos que indican precocidad para más de la mitad de los cruces evaluados. Se encontró una asociación importante entre el tamaño de la vaina y el peso de 100 semillas.
Palabras claves adicionales: dialélico, precocidad, rendimiento, control genético, vigor híbrido.
ABSTRACT
There is a lack of information in Colombia about the type of genetic control that governs the precociousness, yield and components of peas. This study was aimed at evaluating eleven agronomic variables when combinatory ability and heterosis in four lines of pea and three commercial varieties. A complete diallel design (Griffing method 1, model 1) was developed with seven materials in the conditions existing on the Sabana de Bogotá. Variance was estimated and estimated values were used for calculating general combining ability (GCA) and specific combining ability (SCA) and heterosis cf mid-parent value (MPV) better parent value (BPV). GCA was highly significant for eight of the characteristics and SCA was significant for first flower node, pod length, number of seeds per pod and the weight of 100 seeds. The 'Pequinegra', 'Santa Isabel' and 'ICA Tominé' varieties were the best parents due to their high GCA in many of the evaluated characteristics. 'Pequinegra' presented highly significant reciprocal effects for characteristics related to precociousness. The F1 hybrid had better than average behaviour when evaluating heterosis cf mid-parent value for five yield components. Nevertheless, only three F1 were earlier than the better parent. Negative values were found for HMP suggesting precociousness for the half of the hybrids evaluated. There was important association between pod size and the weight of 100 seeds.
Additional key words: diallel design: precocious ness, yield, genetic control, hybrid vigour.
Introducción
SE ESTIMA QUE EN COLOMBIA, para el año 2000, la arveja fue la hortaliza con mayor área cosechada, con 24.620 ha, equivalentes a 27% del área hortícola cosechada total. Sin embargo, predominó el bajo rendimiento: en 2000, se obtuvieron 1,4 t· ha-1, valor que se mantuvo desde 1997 con rendimientos entre 1,0 y 1,4 t· ha-1 (Ministerio de Agricultura y Desarrollo Rural y Corporación Colombia Internacional, 2002). El conocimiento de la habilidad combinatoria ayuda en la formulación de un programa de mejoramiento exitoso para aumentar el rendimiento y sus componentes. La metodología de cruzamientos dialélicos es útil, suministrando la información de la habilidad combinatoria de padres y cruces y la naturaleza genética de los caracteres cuantitativos (Gupta et al., 1986).
En el mejoramiento genético de arveja se ha considerado el inicio de la floración como un indicador importante, de naturaleza cuantitativa, puesto que muestra una variación continua bajo condiciones de campo (Bourion et al., 2002). El control del momento de la floración se debe a un sistema poligénico aditivo con dominancia de la floración tardía. Se estableció que el gen Sn produce un represor denominado calisantina que retarda la floración, posiblemente por la destrucción de la hormona que la promueve (Khvostova, 1983). Durante los pasados años setentas se identificaron varias clases de comportamiento de la floración, distintas entre cultivares, llevando al reconocimiento de cuatro genes mayores en el control de la floración: Sn; Lf, en el control del nudo inherente mínimo hasta la floración; E y Hr, que reducen el efecto tardío de los días cortos sobre el momento de la floración (Bourion et al., 2002).
El número de nudos reproductivos, junto con la tendencia a producir ramas basales, fueron influidos por algunos genes mayores que controlan floración. Entre ellos Sn, que, por la demora en el inicio de la senescencia apical, causa un aumento altamente significativo en el período reproductivo y, por lo tanto, en el número de nudos reproductivos bajo condiciones de campo. Sn también tiene efectos pleiotrópicos sobre el hábito de ramificación; las líneas Sn tienen una marcada tendencia a producir ramas basales, particularmente bajo días cortos. Al mismo tiempo, el alelo Hr, que prolonga la actividad de Sn, aumenta el número de nudos reproductivos y el nacimiento de ramas basales en las líneas Sn a medida que disminuye la longitud de los días (Murfet, 1985, citado por Bourion et al., 2002).
Se considera que la naturaleza de la acción genética que determina el rendimiento del grano tiene penetración incompleta y diferencias en su expresividad, dependiendo del ambiente. Khvostova (1983), basada en una evaluación de híbridos obtenidos por cruzamientos de variedades distinguibles en la expresión de caracteres individuales (semillas por vaina, tamaño de la semilla y número de vainas), afirma que un incremento excesivo en un carácter está asociado negativamente con el desarrollo de los otros y, al mismo tiempo, una disminución en ese carácter es hasta cierto punto compensado por el aumento en otros caracteres. Se cree que cada uno de los componentes del rendimiento tiene su propio rango óptimo, más allá de sus interacciones. La correlación más alta se observa entre los caracteres de productividad de la semilla y el número de semillas por planta, una correlación un poco menor se encuentra entre productividad de la semilla y el número de vainas por planta y aún menor, entre productividad de la semilla y el peso de ésta.
La habilidad de los parentales para combinar bien depende de varias combinaciones genéticas complejas que no pueden ser juzgadas completamente por el desempeño fenotípico y las cualidades adaptativas. La habilidad combinatoria indica el comportamiento genético del material parental, por ello es recomendable seleccionar los padres para hibridización sobre la base de su habilidad combinatoria (Singh y Singh, 1987).
Una interpretación sencilla de heterosis, o vigor híbrido, es la diferencia entre el híbrido y la media de los dos padres. De acuerdo con Robles (1986), la heterosis se aplica principalmente a caracteres de herencia cuantitativa, cuya expresión es compleja por los efectos acumulativos de genes con dominancia, sobredominancia, aditividad y efectos epistáticos, es decir, la expresión final de heterosis se debe a la sumatoria de todos los genes que intervienen en todas las variantes de acción de genes inter-alélicos e intra-alélicos. La heterosis no ocurre de manera general en las especies vegetales. De hecho, algunas especies de plantas autógamas no expresan ninguna depresión endogámica ni heterosis. Sin embargo, se ha indicado la presencia del fenómeno de heterosis en autógamas, aun cuando es más frecuente y más intenso en alógamas (Wehner, 1999).
Gritton (1975) encontró que en arveja los caracteres: altura de la planta, vainas por planta, semillas por vaina y peso de la semilla; mostraban heterosis y depresión endogámica. La HMP de la F1 para rendimiento fue 55% y la HBP, 28%. En cuanto a HCG y HCE, las dos fueron importantes para días hasta la floración, altura de la planta, vainas por planta, semillas por vaina, semillas por planta, peso de la semilla y rendimiento de la semilla. Hubo efectos maternos para vainas por planta, semillas por planta y rendimiento.
Singh, et al. (1980) reportaron que el control genético aditivo fue importante en la expresión de los días hasta la floración, mientras que el período de floración y la altura de la planta, si bien mostraron ser influidos por la aditividad, también fueron influidos fuertemente por efectos dominantes. Dhillon y Chanal (1981) encontraron que los tipos de acción genética aditiva y no aditiva fueron importantes por igual para rendimiento, número de vainas y semillas por vaina. Las variables días hasta la floración y nudos hasta la primera flor fueron influidas sólo por efectos genéticos aditivos. Los efectos recíprocos fueron significativos para todos los caracteres, excepto para semillas por vaina.
Sarawat et al. (1994) hallaron que la mayoría de los híbridos fueron más rendidores que la media parental, pero el rendimiento fue menos estable a través de los ambientes. Hubo correlaciones significativas entre los híbridos F1 y el valor de la media parental para altura de la planta, vainas por planta y peso de 100 semillas, pero no para rendimiento. La heterosis para rendimiento de grano se debió principalmente a más vainas por planta en los híbridos F1. Tanto los efectos genéticos aditivos como los no aditivos fueron importantes en la expresión de todos los caracteres estudiados.
Bourion et al. (2002) estudiaron el determinismo genético de criterios adecuados para mejorar el rendimiento de la semilla y la estabilidad de éste en arveja. Se atribuyó el inicio de la floración a un gen dominante, posiblemente Lf, que controla el número de nudos hasta la primera flor. El grupo de parentales se dividió en dos grupos definidos por los nudos hasta la primera flor, indicando que el alelo del locus de floración tardía puede afectar la arquitectura y la productividad de la planta.
Teniendo en cuenta que a nivel nacional no hay trabajos en arveja que indiquen qué tipo de control genético gobierna los diferentes caracteres agronómicos, tales como precocidad y rendimiento, se propuso iniciar un programa de mejoramiento genético dirigido a sus variedades comerciales, buscando que conserven ciertas características apetecidas tanto por el productor como por el consumidor. El objetivo del presente trabajo fue evaluar las habilidades combinatorias general y específica, los efectos recíprocos y la heterosis entre cuatro líneas y tres variedades comerciales de arveja para once caracteres agronómicos.
Materiales y métodos
Se tomaron siete genotipos de arveja voluble, entre ellos el cultivar diferencial 'WSU 23', reconocido por su resistencia a la razas 1, 2 y 5 de Fusarium oxysporum f. sp. pisi (Van Hall) Snyd. & Hans (Kraft, 2001). Así como los materiales 'Alemania 23', 'Pequinegra' (material tradicional de Nariño, Colombia) y 'Australia 26' son resistentes en invernadero a la raza X del patógeno antes mencionado, el primero conserva su resistencia en campo y el segundo presenta una reacción intermedia entre resistente y susceptible. Con respecto a la raza Y de F. oxysporum f. sp. pisi, se reportó el material 'Tolima 3' con resistencia en invernadero y reacción intermedia en campo (Checa, 1993). Finalmente, se incluyeron la variedad comercial 'Santa Isabel' y la variedad mejorada 'ICA Tominé'; ésta última, con buen potencial de rendimiento, fue obtenida por hibridación ('Australia 23' x 'Bonza Boyacá 2') y posterior selección, en 1978, por el Instituto Colombiano Agropecuario [ICA] (Ligarreto, 1994). Estos materiales fueron cruzados entre ellos con todas las combinaciones posibles, a través de la técnica de hibridación artificial en arveja por emasculación (Ligarreto y Patiño, 2004).
Los siete materiales parentales y los 42 híbridos F1 resultantes, incluyendo los recíprocos, fueron evaluados bajo cubierta en invernaderos de la Sabana de Bogotá, en un diseño de bloques completos a azar con tres repeticiones. Las semillas se sembraron en surcos espaciados 1,5 m, en parcelas de 1,0 m de largo y 0,5 m entre parcelas. Se aplicó a la siembra una dosis de 200 kg· ha-1 del fertilizante NPK 10-30-10.
La evaluación de las diferentes variables se realizó sobre cinco plantas por parcela, y el análisis estadístico se desarrolló con promedios por parcela. Los once caracteres evaluados se agruparon en indicadores de precocidad –días hasta la floración, altura de la planta hasta la floración, nudos hasta la primera flor y días hasta la fructificación–, ancho de vaina, longitud de vaina, número de ramas basales y componentes del rendimiento –número de vainas por planta, número de semillas por vaina, rendimiento por planta y peso de 100 semillas (Medina et al., 1989).
Los valores estimados y el análisis de varianza de las habilidades combinatoria general y específica, así como los efectos recíprocos y maternos, se calcularon de acuerdo con la metodología de Griffing (1956), método 1, modelo 1, por el programa estadístico SAS, versión 8.1, con base en el modelo de programación sugerida por Zhang y Kang (2003). En las variables número de vainas por planta y rendimiento por planta, para corregir la normalidad del supuesto del modelo (εij = NI(0,σ2)) fue necesaria la transformación de los datos por medio de raíz cuadrada. Para la evaluación de heterosis se emplearon las siguientes fórmulas, reportadas por Sarawat et al. (1994), y se realizaron análisis de correlación de Pearson entre los valores de heterosis sobre la media parental y entre los valores de heterosis sobre el mejor padre:

Resultados y discusión
De acuerdo con el análisis de varianza (tabla 1), los híbridos F1 mostraron diferencias altamente significativas en todos los caracteres evaluados, excepto en el número de ramas basales, el número de vainas por planta y el rendimiento por planta. La HCG fue significativa para todas las variables en estudio y no se observó efecto alguno sobre el número de vainas por planta y el rendimiento, indicando la importancia de los efectos genéticos aditivos en la determinación de todos los caracteres, excepto de estos dos. Por su parte, la HCE sólo presentó efectos significativos para los caracteres: nudos hasta la primera flor, longitud de la vaina, número de semillas por vaina y peso de 100 semillas; lo que sugiere que en la determinación de tales caracteres se presentaron tanto efectos genéticos aditivos como no aditivos relevantes, siendo los aditivos los de mayor consideración. La poca importancia de los efectos de la habilidad combinatoria específica para los caracteres: días hasta la floración, altura de la planta hasta la floración, días hasta la fructificación, ancho de vaina, ramas basales, número de vainas por planta y rendimiento por planta; evidencia la baja variabilidad genética presente dentro del grupo de parentales para tales de variables.
En el presente estudio, los días hasta la floración parecen estar influidos sólo por una acción aditiva de los genes, en concordancia con lo hallado por Dhillon y Chanal (1981). Sin embargo, los reportes de varios autores coinciden en afirmar que la magnitud del efecto aditivo es mayor, aunque también son importantes los efectos no aditivos (Gritton, 1975; Singh et al., 1980; Singh et al., 1985; Bourion et al., 2002), de acuerdo con lo afirmado por Khvostova (1983) sobre un sistema poligénico aditivo con dominancia de la floración tardía. Por el contrario, Srivastava et al. (1986) hallaron que la relación de las varianzas de las dos habilidades combinatorias indicaba el predominio de los efectos no aditivos para días hasta la floración y lo corroboró con la alta heredabilidad del carácter.
En cuanto a los nudos hasta la primera flor, los resultados indican la acción conjunta de un sistema aditivo de genes y un efecto no aditivo, con superioridad de la aditividad. Dhillon y Chanal (1981) afirman que este carácter solamente fue influido por efectos genéticos aditivos. Entre tanto, Bourion et al. (2002) sugieren que la herencia del número de nudos hasta la primera flor está determinada principalmente por un gen mayor (posiblemente Lf) asociado con varios genes, actuando de manera aditiva. También, Srivastava et al. (1986) reportan una mayor incidencia de efectos no aditivos en el control genético de los nudos hasta la primera vaina. Para la altura de la planta hasta la floración, sólo fueron importantes los efectos genéticos aditivos. Se ha atribuido la determinación de este carácter tanto a efectos genéticos aditivos como a efectos dominantes, pero a los aditivos con mayor intensidad (Gritton, 1975; Singh et al., 1980; Singh et al., 1985; Singh y Singh, 1987; Srivastava et al., 1986). De igual manera, la aditividad fue el factor genético determinante para el número de ramas basales, coincidiendo con los resultados obtenidos por Bourion et al. (2002), Singh y Singh (1987) y Srivastava et al. (1986).
Los efectos genéticos para ancho y longitud de vaina fueron de tipo aditivo. Para ésta última, también fueron importantes los efectos no aditivos, aunque en menor proporción que los aditivos. De acuerdo con los reportes de Gupta et al. (1986) y Singh et al. (1985), la acción sobre el ancho de vaina es de carácter tanto aditivo como no aditivo, con preponderancia del primero. De igual modo, Singh y Singh (1987) y Gupta et al. (1986) sugieren que el mismo tipo de control genético ocurre en la determinación del largo de vaina. En cuanto al número de semillas por vaina y al peso de 100 semillas, hubo mayor efecto de la acción genética aditiva que de la no aditiva. Se observó el mismo comportamiento en trabajos realizados por Dhillon y Chanal (1981), Gritton (1975), Gupta et al. (1984), Sarawat et al. (1994), Singh et al. (1985), Singh y Singh (1987) y Srivastava et al. (1986).
Es de anotar que la HCG y la HCE no fueron significativas para el rendimiento por planta. Contrario a lo reportado en varios trabajos, en los que se encontró que el rendimiento está determinado tanto por efectos aditivos como no aditivos (Dhillon y Chanal, 1981; Gritton, 1985; Gupta et al., 1984; Sarawat et al., 1994; Singh et al., 1985; Singh y Singh, 1987; Srivastava et al., 1986), en tres de los cinco componentes del rendimiento evaluados hubo acción genética aditiva significativa, y dos de éstos también mostraron algún efecto no aditivo importante. Es posible que no se haya identificado algún tipo de acción genética para rendimiento, teniendo en cuenta lo sugerido por Cousin (1997) en relación con que, aun cuando hay una gran variabilidad genética para componentes de rendimiento –como número de vainas por planta, número de semillas por vaina y peso de 100 semillas–, al hacer cruces procurando mejorar estos caracteres hay a menudo reducción del rendimiento. Por el contrario, la reducción en el área foliar y la altura de la planta, de un lado, y el aumento en la ramificación y el peso de 100 semillas, del otro, favorecen un mayor rendimiento. Así mismo, Sarawat et al. (1994) afirman que el rendimiento del grano en arveja tiene pocas posibilidades de mejorarse por selección de los componentes individuales de rendimiento, ya que sugieren la presencia de compensación entre los diferentes componentes.
Los efectos recíprocos sólo fueron relevantes para los caracteres días hasta la floración y días hasta la fructificación. El primer caso parece deberse a efectos maternos, en tanto que para los días hasta la fructificación, los efectos recíprocos pueden ser explicados por la incidencia tanto de efectos maternos como no maternos. Dhillon y Chanal (1981) también observaron la presencia de efectos recíprocos para el carácter días hasta la floración (tabla 1).
Para el conjunto de genotipos evaluados, la acumulación de genes de carácter aditivo se considera la estrategia más adecuada, desde el punto de vista de mejoramiento genético, para los caracteres: días hasta la floración, altura de la planta hasta la floración, días hasta la fructificación, ancho de vaina y ramas basales. En el caso de nudos hasta la primera flor, longitud de la vaina, número de semillas por vaina y peso de 100 semillas, también se debe tener en cuenta el control genético no aditivo.
Los efectos estimados de la HCG de cada parental sobre los caracteres evaluados fueron contrastantes (tabla 2). Las variedades 'Pequinegra', 'ICA Tominé' y 'Santa Isabel', junto con las líneas 'Alemania 23' y 'Australia 26', tuvieron una incidencia significativa, tanto positiva como negativa, sobre gran parte de los caracteres estudiados. Los materiales que mostraron ser los mejores combinadores generales fueron las variedades 'Santa Isabel', 'ICA Tominé' y 'Pequinegra', por su efecto favorable sobre la precocidad de sus plantas F1 y sobre algunos de los componentes de rendimiento. Para estos genotipos, la primera flor emergió aproximadamente un nudo más abajo, de manera que disminuyó la altura de la planta hasta la floración entre 12 y 14 cm y, en consecuencia, la duración del período vegetativo se redujo en hasta 2,5 d. Este comportamiento está en concordancia por lo mencionado por Khvostova (1983), quien sostiene que por cada nudo vegetativo la floración tarda casi 2 d (tabla 2).
En las variables ancho y largo de vaina, todos los materiales parentales influyeron tanto positiva como negativamente. En el caso de los cruces cuyos paren-tales femeninos fueron 'Santa Isabel', 'ICA Tominé' y 'Pequinegra', hubo aumento del tamaño de la vaina, tanto en longitud como en ancho, resaltándose 'Santa Isabel' e 'ICA Tominé' como los parentales con mayor efecto positivo sobre estas variables. Por otra parte, 'WSU 23', 'Alemania 23' y 'Tolima 3' incidieron con reducciones sustanciales en el tamaño de la vaina. Es conveniente subrayar la influencia alta que tienen los caracteres determinantes del tamaño de la vaina (ancho y longitud) en el peso de la semilla, lo que indica que el tamaño del grano es factible de ser mejorado por medio del aumento en estas variables. Para tal efecto, las variedades 'Pequinegra', 'Santa Isabel' e 'ICA Tominé' son los parentales más indicados, con el fin de aprovechar su alta HCG asociada a estos caracteres. Este comportamiento puede explicar la amplia adaptación de estas variedades a las distintas condiciones climáticas de cultivo de arveja en Colombia.
Por el contrario, si bien los materiales 'Australia 26', 'Alemania 23' y 'Tolima 3' fueron buenos combinado-res generales, su repercusión no es la deseada para un programa de mejoramiento en arveja, cuyo objetivo sea obtener materiales precoces o de alto rendimiento, pero son convenientes por sus genes de resistencia a diferentes razas de F. oxysporum f. sp. pisi. Los cruces F1 de estas líneas fueron más tardíos, presentaron plantas más altas, lo que no es conveniente para las labores y la cosecha en una producción comercial; además presentaron vainas de menor tamaño y granos más livianos que los cruces provenientes de los materiales parentales 'Santa Isabel', 'ICA Tominé' y 'Pequinegra'.
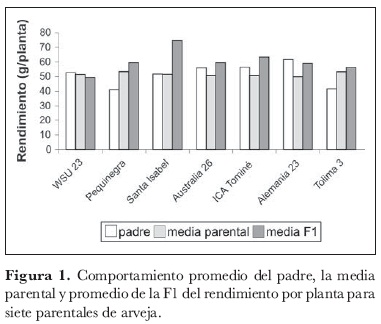
No obstante, algunos caracteres, como ancho y longitud de vaina, ramas basales y peso de 100 semillas, mostraron aumentos importantes. El rendimiento por planta per se sólo se incrementó en los híbridos F1 de la variedad comercial 'Santa Isabel', lo que sugiere que es posible obtener materiales mejorados de arveja que cumplan con las características comerciales del mercado colombiano (figura 1).
En la estimación de la HCE, para todas las variables hubo por lo menos un cruzamiento con algún efecto significativo, excepto para número de vainas y rendimiento por planta (tabla 3). De 21 cruces F1 directos, 13 presentaron efectos significativos de HCE, en algunos casos para más de una característica. Para las F1 que mostraron algún efecto importante, alguno o sus dos padres fueron buenos combinadores generales para dicha característica. Gupta et al. (1984 y 1986) encontraron este mismo comportamiento en cruces con efectos específicos significativos para todos los caracteres que estudiaron. En este trabajo, los cruzamientos específicos tuvieron progresos evidentes para algunos pocos caracteres, como tamaño de vaina, representado por su ancho y longitud, y peso de 100 semillas.
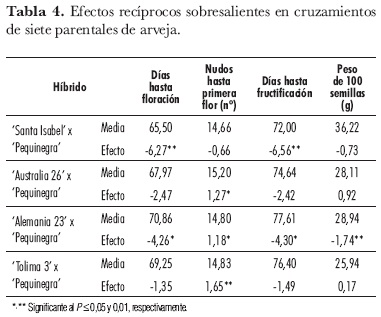
De los 21 cruzamientos recíprocos, cuatro presentaron efectos recíprocos significativos (tabla 4). Los materiales 'Santa Isabel', 'Australia 26', 'Alemania 23' y 'Tolima 3' cruzados con 'Pequinegra' como parental masculino tuvieron efectos importantes en las variables: días hasta la floración, días hasta la fructificación, nudos hasta la primera flor y peso de 100 semillas. La variedad 'Pequinegra' incidió en cruces precoces, con reducciones de hasta 6 d, al cruzarse con la variedad 'Santa Isabel', posiblemente por efectos maternos sobre estos caracteres.
Los resultados divergentes para el análisis de varianza de la HCG y la HCE, presentados en este documento, con respecto a los de otros trabajos pueden ser explicados, entre otros factores, por el hábito de crecimiento de los materiales, las condiciones ambientales que influyen la expresión de los caracteres y por el modelo fijo del diseño utilizado, en el que los resultados se restringen a la población de parentales involucrada. De acuerdo con Robles (1986), no es aceptable considerar a la heterosis sólo por su mayor expresión (heterosis positiva), ya que en algunos casos una menor expresión (heterosis negativa) es favorable para algunos caracteres. En la evaluación de heterosis para los caracteres indicadores de precocidad se considera propicio un valor negativo, también se tiene en cuenta el padre más precoz como el mejor padre; mientras que para el resto de variables, una heterosis positiva es la más apropiada (tabla 5).
Se observa un comportamiento heterótico de los cruzamientos para las variables rendimiento (18,45%), número de vainas por planta (16,44%) y ramas basales (15,69%), con respecto a la media parental, en tanto que para la HBP, aunque hubo valores altos de heterosis en distintos caracteres, éstos no son los más apropiados, ya que la norma fue plantas F1 más tardías y plantas de menor rendimiento que los mejores padres. Sin embargo, se debe anotar que la desviación estándar para rendimiento, número de vainas por planta y ramas basales fue elevada, lo que indica un amplio rango de variación en el comportamiento de estos caracteres, hecho que no es conveniente en un programa de mejoramiento genético, ya que no hay certeza del desempeño que tendrán los materiales. De otra parte, los caracteres que evidenciaron un desempeño más constante o una menor variación fueron: días hasta la floración, días hasta la fructificación, ancho de vaina, longitud de vaina y peso de 100 semillas (figura 2).

En cuanto a la correlación entre heterosis para los diferentes caracteres evaluados (tabla 6), los valores de la HMP para días hasta la floración y días hasta la fructificación estuvieron altamente correlacionados de manera positiva; lo mismo ocurrió con la altura de la planta hasta la floración y número de nudos hasta la primera flor, pero éstos dos grupos de variables no se correlacionaron entre sí. No obstante, sobresalió la precocidad de los híbridos F1, debida principalmente a la menor duración del período vegetativo con respecto a la media parental.
Se encontró que la heterosis para la altura de la planta hasta la floración influyó positivamente en la heterosis de varios componentes de rendimiento, como longitud de vaina, ramas basales, número de vainas por planta y número de semillas por planta, y hasta en el rendimiento mismo (r = 0,51), concordando con lo señalado por Singh et al. (1985) respecto a que la superioridad del rendimiento puede deberse al efecto combinado de los componentes de rendimiento. Otros valores de la HMP que afectaron de manera favorable la heterosis sobre el rendimiento fueron la longitud de vaina, ramas basales, número de semillas por vaina, pero el carácter más relevante fue el número de vainas por planta (r = 0,95), tal como lo referencia Sarawat et al. (1994). Estos autores también concluyen que este componente del rendimiento es altamente susceptible al ambiente, mientras que el número de semillas por vaina y el peso de la semilla están más determinados por el genotipo.
Literatura citada
Bourion, V., G. Fouilloux, C. Le Signor e I. Lejeune-Hénaut. 2002. Genetic studies of selection criteria for productive and stable peas. Euphytica 127, 261-273. [ Links ]
Checa, O. 1993. Determinación de razas del hongo Fusarium oxysporum f. sp. pisi y su efecto en materiales de arveja en el departamento de Nariño. Tesis de maestría. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira. [ Links ]
Cousin, R. 1997. Peas (Pisum sativum L.). Field Crops Res. 53, 111-130. [ Links ]
Dhillon, G.S. y G.S Chanal. 1981. An analysis of combining ability and reciprocal effects in garden pea (Pisum sativum L.). J. Res. Punjab Agric. Univ. 18(4), 359-364. [ Links ]
Griffing, B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Bio. Sci. 9, 463-493. [ Links ]
Gritton, E. 1975. Heterosis and combining ability in a diallel cross of peas. Crop Science 15, 453-457. [ Links ]
Gupta, K.R., B.S Dahiya y K.P. Singh. 1986. Combining ability studies over environments in pea. Crop Improv. 13(2), 134-137. [ Links ]
Gupta, K.R., R.S. Waldia, B.S. Dahiya, K.P. Singh y D.R. Sood. 1984. Inheritance of seed yield and quality traits in peas (Pisum sativum L.). Theor. Appl. Genet. 69, 133-137. [ Links ]
Khvostova, V. 1983. Genetics and breeding of peas. USSR Academy of Sciences, General Biology Division. Oxonian Press, New Delhi. [ Links ]
Ligarreto, G.A. y M.P. Patiño. 2004. Hibridación artificial en arveja. Centro de Investigación y Extensión Rural (CIER). Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. Serie técnica Nº 1, 12 p. [ Links ]
Ligarreto, G.A. 1994. 'ICA Tominé', variedad de arveja voluble para clima frío. Instituto Colombiano Agropecuario (ICA), Corporación Colombiana de Investigación Agropecuaria (Corpoica), Federación Nacional de Cultivadores de Cereales y Leguminosas (Fenalce). Plegable divulgativo N° 275. [ Links ]
Medina, C., M. Escobar y M. Lobo. 1989. Evaluación primaria y caracterización de germoplasma de arveja Pisum sativum L. pp. 12-28. En: Instituto Interamericano de Cooperación para la Agricultura (IICA) y Prociandino. Boletín técnico N° 1. [ Links ]
Ministerio de Agricultura y Desarrollo Rural y Corporación Colombia Internacional. 2002. Manual del exportador de frutas, hortalizas y tubérculos en Colombia. Bogotá [ Links ].
Robles, R. 1986. Genética elemental y fitomejoramiento práctico. Limusa, México. 460 p. [ Links ]
Sarawat, P., F.L. Stoddar, D.R. Marshall y S.M. Ali. 1994. Heterosis for yield and related characters in pea. Euphytica 80, 39-48. [ Links ]
Singh, S. P. y R.P. Singh. 1987. Diallel analysis for combining ability in pea. Acta Agronomica Hungarica 36(1-2), 89-95. [ Links ]
Singh, V.S., U.S. Santoshi, K. Singh e I.B. Singh. 1985. Combining ability in pea. Crop Improv. 12(1), 52-54. [ Links ]
Singh, S., B.S. Dahiya y P.S. Sidhu. 1980. Genetic architecture of some morphological traits in peas (Pisum sativum L.). Genet. Agr. 34, 289-298. [ Links ]
Srivastava, P.L., U.S. Santoshi y H.G. Singh. 1986. Combining ability and heterosis in pea. Crop Improv. 13 (1), 20-23. [ Links ]
Wehner, T.C. 1999. Heterosis in vegetable crops. pp 387-397. En: Coors, J. y S. Pandey. The genetics and exploitation of heterosis. American Society of Agronomy, Madison, WI. [ Links ]
Zhang, Y. y M.S. Kang. 2003. Diallel-SAS: a program for Griffing's diallel methods. pp. 1-15. En: Kang, M.S. (ed.). Handbook of formulas and software for plant geneticists and breeders. Food Product Press, New York. [ Links ]

