










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.23 no.2 Bogotá July/Dez. 2005
FISIOLOGÍA DE CULTIVOS
Aspectos fisiológicos de estevia (Stevia rebaudiana Bertoni) en el Caribe colombiano: I. Efecto de la radiación incidente sobre el área foliar y la distribución de biomasa
Physiological aspects of stevia (Stevia rebaudiana Bertoni) in the Colombian Caribbean: I. Effects of attendant radiation on leaf area and biomass distribution
Alfredo Jarma1, Teresita Rengifo2 y Hermes Araméndiz-Tatis3
1 Profesor asociado, Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). e-mail: ajarma@sinu.unicordoba.edu.co
2 Asistente técnico, Organización de las Naciones Unidas para la Agricultura y Alimentación (FAO), Medellín (Colombia). e-mail: trengifo50@yahoo.es
3 Profesor asociado, Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). e-mail: haramendiz@hotmail.com
Fecha de recepción: 30 de agosto de 2005. Aceptado para publicación: 21 de noviembre de 2005
Resumen
Stevia rebaudiana Bertoni es uno de los 154 miembros del género Stevia. El componente edulcolorante de sus hojas se debe a glucósidos de diterpeno. Los principales glucósidos de esteviol son: esteviósido, rebaudiósido A, rebaudiósido C y dulcósido A. Este trabajo se realizó en Montería (Colombia), con el propósito de evaluar el efecto de cuatro niveles de radiación incidente sobre el comportamiento fisiológico de S. rebaudiana, bajo las condiciones ambientales del valle del Sinú, en el Caribe colombiano. Se planteó un diseño completo al azar con arreglo factorial, en el que los factores fueron los porcentajes de la radiación incidente en la región (19%, 24%, 56% y 100%) y los genotipos de estevia 'Morita 1' y 'Morita 2'. Los resultados indicaron que el área foliar de 'Morita 2' es superior a 'Morita 1', independientemente de la radiación. Los altos niveles de radiación incidente (100% y 56%) registraron la mayor acumulación de biomasa de las hojas, siendo superior en 'Morita 2'. La mayor proporción de la biomasa de las hojas, con respecto a la del tallo, en los primeros 60 d indica que la planta se dedicó a fortalecer su aparato fotosintético; luego, la migración de fotoasimilados se hizo en mayor proporción hacia los tallos, terminando con una tendencia estable hacia ambas demandas.
Palabras claves adicionales: esteviósidos, edulcolorantes, radiación incidente, materia seca, fotoasimilados.
Abstract
Stevia rebaudiana Bertoni is one of the Stevia genus' 154 members. The sweetening component of its leaves is due to dipterpene glycosides. The major steviol glycosides are: stevioside, rebaudioside A, rebaudioside C and dulcoside A. This research was carried out at Montería (Colombia); it evaluated the effect of four levels of attendant radiation in the climatic conditions found in the Sinú river valley on the physiological behaviour of S. rebaudiana. A completely random design was used, employing percentage of attendant radiation (19%, 24%, 56% and 100%) and Stevia genotypes ('Morita 1' and 'Morita 2') as factors. The results indicated that the leaf area of 'Morita 2' was always bigger than that of 'Morita 1' and radiation level did not influence this variable. The biggest accumulation of dry mass on leaves returned the highest levels of attendant radiation (100% and 56%). 'Morita 2' was better able to accumulate dry mass than 'Morita 1'. The fact that leaves accumulated more biomass than the stems during the first 60 d after being transplanted showed that plants were working to strengthen their photosynthetic ability during this period. This was followed by a greater migration of substances produced by photosynthesis towards the stems. The tendency stabilised toward both demands at the end of the period being studied.
Additional key words: steviosides, sweetener, attendant radiation, dry mass, photoassimilates.
Introducción
STEVIA REBAUDIANA BERTONI, o 'hierba dulce', es la principal productora de un edulcorante natural no calórico llamado esteviósido, cuyo poder es hasta 300 veces mayor que el de la sacarosa o azúcar de caña; es lo más parecido al azúcar entre todos los edulcorantes naturales, distinguiéndose de los edulcorantes artificiales por no tener sabor metálico y no ser cancerígeno (Schwebel, 2005; Galván y Guzmán, 2003). Es una planta herbácea perenne de la familia Asteraceae, que incluye plantas tan conocidas como el diente de león, el girasol, la achicoria y el crisantemo. Las hojas lanceoladas tienen aproximadamente 5 cm de longitud y 2 cm de ancho y se disponen alternadas, enfrentadas de dos en dos. En la naturaleza, la altura de las plantas varía entre 40 y 80 cm, pero cultivadas pueden llegar a 1,0 m de altura. Puede utilizarse para la producción comercial por un periodo de cinco o más años, dando varias cosechas anuales a partir de la parte aérea de la planta. Las raíces permanecen en el suelo y permiten el rebrote de la planta, aunque no macolla. Plantas con un metro de altura producen un peso seco medio de 70 g y el peso seco de las hojas puede variar entre 15 y 35 g/planta. Por tratarse de una planta que se reproduce sexualmente por fecundación cruzada (alógama), no debe extrañar la diversidad fenotípica (apariencia) que se observa en las poblaciones nativas en Paraguay; esto provoca una gran diferencia de contenido de edulcorantes entre las distintas plantas y/o clones, aunque en Colombia estas variaciones no se han referenciado (Secretaría de Agricultura de Antioquia, 2000).
Como resultado de los trabajos de laboratorio realizados, se conoce actualmente que la estevia tiene varios componentes edulcorantes, con una concentración en la hojas de los cuatro más importantes, así: esteviósido, 5%-10%; rebaudiósido A, 2%-4%; rebaudiósido C, 1%-2%, y dulcósido A, 0,5%-1,0% (Midmore y Rank, 2002). Estos componentes edulcorantes de las hojas de estevia son glucósidos de diterpeno sintetizados, al menos en los estados iniciales, usando la misma ruta del ácido giberélico (Geuns, 2003). Estos diterpenos se sintetizan a partir de unidades de acetato del acetil-coenzima A en la ruta del ácido mevalónico (MVA). Éste último ingresa al cloroplasto y se convierte en pirofosfato de isopentenilo (IPP); luego, se convierte en pirofosfato de geranilgeranilo, un compuesto de 20 carbonos que sirve como donador de todos los átomos de carbono de las giberelinas; más tarde, en pirofosfato de copalillo, con dos sistemas de anillos, y por último, en kaureno, un compuesto que tiene cuatro sistemas de anillos. El kaureno es el compuesto donde divergen los glucósidos de esteviol y las giberelinas. En estevia, el kaureno es convertido a esteviol -espina dorsal de los glucósidos de ditereno- en el retículo endoplásmico; este esteviol puede ser glucosilado o rhamnosilado en el aparato de Golgi, para formar los principales edulcorantes, como el esteviósido, los rebaudiósidos A y C y el dulcósido A, que van finalmente a las vacuolas (Totté et al., 2000 y 2003). El propósito de estos compuestos en la planta de estevia no está todavía claro, pero su alta concentración en la hoja y la conservación de la ruta dentro de las especies indicaría que, en algún momento del proceso evolutivo, su presencia confirió una ventaja significante a los individuos que los poseían. Algunos investigadores afirman que estos compuestos actúan para rechazar ciertos insectos y otros especulan que es un mecanismo detallado para controlar los niveles de ácido giberélico (Brandle et al., 2002; Brandle, 2005). No obstante su función dentro de la planta, algunos autores afirman que la calidad de los edulcorantes de las hojas de S. rebaudiana varía ampliamente debido a factores ambientales, como tipo de suelos, método de irrigación, prácticas de cultivo, proceso, almacenamiento y luz solar (Secretaría de Agricultura de Antioquia, 2000).
Para comprender cómo afecta la luz a la tasa fotosintética, hay que considerar inicialmente cuánta energía luminosa proporciona la luz del sol. En el límite superior de la atmósfera la irradiancia total es de 1.360 W· m-2 (constante solar), que varía ± 2% por la órbita elíptica de la Tierra e incluye las longitudes de onda ultravioletas e infrarrojas; sin embargo, cuando esta radiación atraviesa la atmósfera hasta la superficie terrestre, gran parte se pierde por la absorción y dispersión producidas por vapor de agua, polvo, CO2 y ozono, por lo que solamente llegan a las plantas alrededor de 900 W· m-2, dependiendo de la época del año, hora del día, altitud, latitud, condiciones atmosféricas y otros factores. De este valor, cerca de la mitad corresponde al infrarrojo, un 5% es ultravioleta y el resto tiene longitudes de onda comprendidas entre 400 y 700 nm, capaces de inducir fotosíntesis. Esta radiación, expresada en unidades de energía, se conoce como radiación fotosintéticamente activa (RFA), cuyos valores generalmente pueden estar comprendidos entre 400 y 500 W· m-2, dependiendo de los factores mencionados.
La cantidad efectiva de luz en la fotosíntesis se conoce como flujo fotónico fotosintético (FFF). La luz solar se encuentra en el intervalo micromolar; en un día despejado, equivale a un valor comprendido entre 2.000 y 2.300 µmol· m-2· s-1. En cualquier entorno, la irradiancia varía dentro de una misma cobertura vegetal. Entre 80% y 90% del FFF lo absorbe una hoja representativa, aunque este valor varía de manera considerable con la edad y la estructura de la hoja. El resto, que en su mayor parte lo constituyen longitudes de onda del verde y del rojo lejano, se transmite a las hojas inferiores o al suelo o se refleja hacia los alrededores. Por tanto, de la radiación que se absorbe y que tiene la capacidad potencial de inducir la fotosíntesis, es habitual que más de 95% se transforme en calor, capturándose durante la fotosíntesis menos de 5% (Salisbury y Ross, 2000).
Los mismos autores indican que por encima de cierto nivel de irradiancia, conocido como saturación lumínica, el incremento en la luz ya no causa incremento alguno en la fotosíntesis y, entre la oscuridad y el nivel de saturación, existe una irradiancia en la que la fotosíntesis está en equilibrio con la respiración (el intercambio neto de CO2 es cero), conocido como punto de compensación luminosa. Este punto varía con la especie, con la irradiancia durante el crecimiento, con la temperatura a la que se efectúen las mediciones y con la concentración de CO2, aunque en hojas que crecen al sol suele ser alrededor de 2% de la luz solar plena. El incremento en la materia seca puede ocurrir sólo cuando la irradiancia se encuentra por encima del punto de compensación luminosa.
En trabajos realizados en alfalfa sobre la fijación neta de CO2 en la fotosíntesis, así como de la liberación de CO2 en la respiración, se observó que, en primer lugar, la máxima fijación de CO2 se presentaba hacia el mediodía, cuando la irradiancia estaba en su punto más elevado; sin embargo, cuando las plantas se expusieron a sombras de corta duración causadas por nubes, las tasas de fijación de CO2 disminuyeron (Salisbury y Ross, 2000).
Investigaciones realizadas en 1997 por Páez et al. sobre aclimatación de Panicum maximum a diferentes regímenes lumínicos (10%, 30% y 100%) indicaron un incremento lineal de la altura de las plantas a través del tiempo, pero los valores más altos se registraron cuando se expusieron a 30% de luz ambiental (sombra parcial). El número de hojas se incrementó a medida que la irradiancia se incrementaba, y el área foliar, la materia seca de hojas y la biomasa fueron mayores en las plantas expuestas a 100% de radiación ambiental.
Páez et al. (1998) reportaron que plantas de Barleria lupulina crecidas en la sombra (30% de luz solar), en relación con las expuestas a 100% de luz solar, presentaron una mayor área foliar. No se encontraron diferencias significativas en la biomasa de hojas y tallos y la biomasa total de las plantas crecidas bajo 100% de luz solar, en relación con las crecidas bajo sombra parcial, a los 20, 50 y 200 d; sin embargo, la biomasa de las raíces fue significativamente mayor en las plantas crecidas al sol a los 100 d.
Conover y Flohr (2003) investigaron el efecto de tres niveles de luz (100%, 70% y 50%) sobre el crecimiento y rendimiento del genotipo de tomate 'Pixie Hibryd II'. Los resultados indicaron que la altura de planta fue estadísticamente mayor en el máximo nivel de sombra (50%). El número de frutos totales producidos por planta, el peso total de frutos por planta y el peso de cada fruto no presentaron diferencias estadísticas para los niveles de luz evaluados.
En el país se tienen informes sobre la introducción de S. rebaudiana en los departamentos de Valle del Cauca y Antioquia a comienzos de la década de los noventas, desconociéndose su procedencia y año de introducción. En 1994, la Corporación para la Diversificación del Ingreso Cafetero (Cordicafé) y la Secretaría de Agricultura de Antioquia suscribieron un convenio de cooperación con la empresa japonesa Jaido, en convenio con Morita Co., para evaluar la adaptación al trópico de la variedad 'Morita 1'. Este genotipo registró baja tolerancia a Septoria sp., por lo que en 1995 se importaron esquejes de Japón de una nueva selección hecha por el señor Morita, denominada 'Morita 2', que registraba un mejor comportamiento ante el patógeno.
En la actualidad hay registros de la siembra de esta especie en los departamentos de Antioquia (Bajirá), Córdoba (Valencia y Planeta Rica), Tolima, Huila, Valle del Cauca y Meta; sin embargo, las técnicas de manejo, los genotipos utilizados y el desconocimiento del cultivo han sido limitantes para una producción sostenible y eficiente (Universidad Eafit, 2004). En la actualidad, se desconoce el comportamiento de esta especie bajo los diversos factores ambientales que caracterizan a la región Caribe, fundamental si los agricultores, principalmente los pequeños, establecen cultivos intercalados con estevia, como es lo usual. Por otro lado, el interés emergente sobre la planta radica en el uso intensivo de mano de obra para su producción, dado que utiliza al año alrededor de 245 jornales permanentes/ha, lo que representa una alternativa para generar empleo, además de contribuir a la diversificación de los renglones agrícolas de la región y el país (Secretaría de Agricultura de Antioquia, 2000).
Ante la inexistencia de trabajos de investigación en el país referentes al comportamiento de esta especie frente a las variables ambientales, este estudio tuvo como finalidad evaluar el efecto de cuatro niveles de radiación incidente sobre el crecimiento de dos genotipos de Stevia rebaudiana en el valle del río Sinú, en el Caribe colombiano.
Materiales y métodos
El trabajo se realizó en los campos experimentales de la Facultad de Ciencias Agrícolas de la Universidad de Córdoba, con sede en Montería (Colombia), durante el período comprendido entre julio de 2002 y abril de 2003. El campo se encuentra ubicado a 13 msnm, 8° 52' de latitud norte y 76° 48' de longitud oeste, y tiene una temperatura promedio anual de 28 °C, humedad relativa promedio de 80%, precipitación promedio anual de 1.200 mm, en una zona de transición entre Bosque Húmedo y Bosque Seco Tropical, según Holdridge. Para evaluar el efecto de los cuatro niveles de radiación incidente sobre el comportamiento de la materia seca y el área foliar de los dos genotipos de S. rebaudiana, se utilizaron mallas polisombra negras de polietileno de alta densidad, con aditivo UV para resistencia a la intemperie, que permitían el paso de 19%, 24% y 56% de radiación incidente, y un testigo sin polisombra, que representó el 100% de la radiación incidente.
Al disponer en el área experimental de condiciones de suelo y ambientales homogéneas, se utilizó un diseño completamente aleatorizado con estructura de tratamiento factorial, en el que intervinieron dos factores: radiación incidente (factor A) y genotipos (factor B). El factor A estuvo compuesto por cuatro niveles (19%, 24%, 56% y 100%) y el factor B, por dos niveles ('Morita 1' y 'Morita 2'), para un total de ocho tratamientos distribuidos en cuatro repeticiones. La unidad experimental (UE) estuvo constituida por 80 plantas sembradas en bolsas plásticas negras con capacidad para 5 kg, ubicadas una contigua a la otra. El número total de plantas evaluadas en la investigación fue 2.560. Las lecturas se realizaron cada 15 días en tres plantas escogidas al azar por UE durante ocho meses, y los análisis se hicieron en tres épocas: 60, 150 y 255 días después del transplante (ddt).
La información fue analizada estadísticamente a través del Statistical Analysis System (SAS) de la Facultad de Agronomía de la Universidad de Córdoba. Se utilizó como prueba de comparación de medias a la diferencia mínima significativa (DMS), en niveles significantes de 5% y 1%. Las interacciones que resultaron significativas o altamente significativas se descompusieron gráfica y numéricamente a través de un Anova, con la tasa de varianza F significante al 5%.
Las mediciones de la radiación incidente se realizaron con una barra cuántica Li-Cor 184, con la que se tomaron mensualmente lecturas en la parte superior, media e inferior del dosel de las plantas a diferentes horas del día (cada hora, entre las 08:00 y las 14:00 h).
Variables evaluadas
Área foliar: Se determinó con el método gravimétrico, utilizando un sacabocado y una balanza electrónica (Scientech Sl 5000); la lectura se tomó en centímetros cuadrados y correspondió a la relación entre el peso de la muestra de área foliar y el peso del total del área foliar de la planta.
Materia seca de hojas: El total de las hojas de cada planta muestreada se sometió a secado en estufa (Memmert) a 70 °C durante 72 h; después de este tiempo se pesó el material vegetal seco.
Materia seca total del dosel: Se consideró como la materia seca total de la parte aérea por planta, calculada como la suma de las biomasas de hoja y tallo.
Distribución de la materia seca en el dosel: Para cada genotipo y en cada nivel de sombreamiento, se evaluó la distribución de la materia seca de las hojas y el tallo, a 60, 150 y 255 ddt. Desafortunadamente, en este estudio no se consideró el sistema radical.
Resultados y discusión
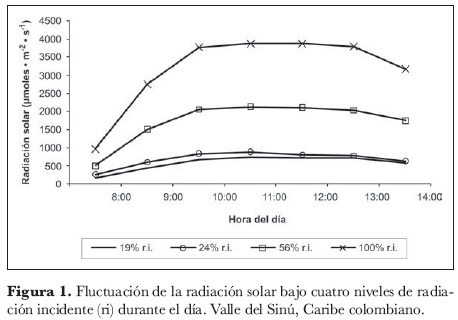
La figura 1 registra la fluctuación de la radiación solar incidente que recibieron las plantas a través del día. En ella se detalla este parámetro bajo cada uno de los tres tratamientos con polisombra, así como la del testigo (sin polisombra), que se asumió como 100% de radiación incidente. El comportamiento en todos los estratos de luz fue similar, registrando una tendencia creciente en horas de la mañana, estable hacia el mediodía y con tendencias a declinar hacia las horas de la tarde. Las radiaciones más altas se registraron alrededor de las 11:00 h para todos los tratamientos, observándose los valores más altos en el testigo con 3.885 µmol· m-2· s-1; en segundo lugar, la poli-sombra que permitía el paso del 56% de radiación incidente, registró una máxima radiación de 2.132 µmol· m-2· s-1; y por último, las más bajas radiaciones se presentaron en las polisombras de 24% y 19% de radiación incidente, con 891,5 y 741,3 µmol· m-2· s-1, respectivamente.
Área foliar
El análisis de varianza realizado a los 60, 150 y 255 ddt indicó que no se presentaron diferencias significativas para la interacción genotipo x radiación. Al analizar el efecto individual de los factores, se observaron de modo consistente diferencias estadísticas (P ≤ 0,05) entre geno-tipos en las tres épocas, señalando que 'Morita 2' presenta mayor área foliar, comparado con 'Morita 1', independientemente del nivel de radiación. A los 60 ddt la radiación no mostró, al parecer, efectos importantes sobre esta variable, pero a los 150 y 255 ddt sí se encontraron diferencias significativas (P ≤ 0,05). La DMS realizada para radiación (α = 0,05) a los 150 ddt señala que el área foliar de las plantas de estevia expuestas a niveles de radiación incidente de 100% y 56% fue estadísticamente más alta respecto a los niveles sombreados de 19% y 24%. A los 225 ddt se observó la misma tendencia, aunque entre los niveles de 24% y 100% no se presentaron diferencias estadísticas (figura 2).
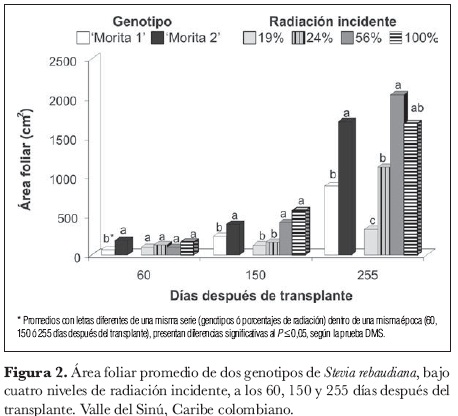
Los anteriores resultados sugieren que los niveles de radiación más altos, independientemente del genotipo, producen un incremento en el área foliar de la estevia. Esta respuesta podría explicarse en parte por el mayor número de hojas por planta (resultado no discutido en este trabajo), que registró valores de 1.061, 1.142, 715 y 119 hojas por planta para los porcentajes de 100%, 56%, 24% y 19% de radiación incidente, respectivamente (Rengifo, 2003). Es posible que este incremento de la superficie fotoasimilatoria se haya traducido en una mayor intercepción de la luz y una mayor fijación de CO2, sugiriéndose que las plantas expuestas a estas radiaciones realizarían mayores tasas de fotosíntesis, en comparación con las que crecen en niveles de menor radiación. Valores más altos de radiación pudieran haber causado una fotoinhibición de la fotosíntesis, con el consecuente efecto negativo sobre el desarrollo del área foliar; sin embargo, esto no pudo ser demostrado en este trabajo.
Paytas (2005), trabajando con poblaciones en algodón, afirma que el índice de área foliar (IAF) óptimo no es estático para un determinado cultivo, sino que cambia de acuerdo con las variaciones de la intensidad de la luz y que, a medida que el IAF se incrementa, aumenta la intercepción de luz y la fotosíntesis neta hasta alcanzar un valor crítico, más allá del cual no hay incremento de la fotosíntesis. De acuerdo con Salisbury y Ross (2000), es evidente el hecho de que la luz muchas veces limita la fotosíntesis, demostrándose en la disminución de las tasas de fijación de CO2 cuando las plantas se exponen a sombras (baja radiación incidente). Esta afirmación es discutible, si se considera que el área foliar responde de manera plástica con la variación de las especies.
En cuanto al factor genotipo, los resultados evidenciaron que 'Morita 2' presentó durante todas las fechas analizadas una mayor área foliar que 'Morita 1', de manera independiente al efecto de la radiación. Esta ventaja tal vez se deba a una condición genética que puede ser 'potencializada' a favor de este genotipo, en la medida en que reciba suficiente radiación solar para expresar los máximos valores de esta variable, máxime cuando los valores del número de hojas por planta fue estadísticamente similar para los dos recursos genéticos (Rengifo, 2003).
Los resultados mencionados son compatibles con investigaciones realizadas sobre aclimatación de Panicum maximum (Páez, 1997), sometida a diferentes regímenes lumínicos, en las que el área foliar fue mayor en las plantas expuestas a 100% de irradiancia, presentándose reducciones de 17% y 93,5% para los tratamientos con 30% y 10% de luz ambiental, respectivamente; aunque es importante considerar que ésta es una planta con metabolismo fotosintético C4. Estas respuestas, sin embargo, no siempre son en el mismo sentido, ya que el efecto de la radiación sobre este parámetro puede variar con la especie. En algunos casos, la planta reduce su lámina foliar ante niveles altos de radiación, como un mecanismo para evitar pérdidas excesivas de agua por transpiración; respuestas de este tipo fueron observadas en habichuela por Jarma et al. (1999), quienes reportaron un incremento del área foliar en niveles de penumbra, sugiriendo que en esta especie la planta procura la expansión máxima de la lámina foliar para captar con mayor eficiencia la luz disponible. Así mismo, otros factores de cultivo pueden afectar la expresión del área foliar en función de la radiación interceptada. Tewolde et al. (2005), en trabajos con algodón, reportaron que el índice de área foliar se redujo en la medida en que hubo menor radiación debida a la reducción en la distancia entre los surcos. De manera similar, Mansab et al. (2003), al evaluar cultivos intercalados de soya y trigo, encontraron que el área foliar de las plantas de soya en las fases tempranas de crecimiento fue menor cuando el cultivo estuvo intercalado, debido a que la RFA interceptada fue menor.
La importancia del área foliar para la distribución de la luz y el uso eficiente de la radiación interceptada ha sido enfatizada recientemente por Rahimian y Zand (2004) en trabajos desarrollados con cultivares antiguos y modernos de trigo. Aunque en estevia no se tienen registros, es importante mencionar que el comportamiento observado en esta especie muestra una tendencia similar a las plantas de sol, que se saturan a niveles muchos más altos de luz que las plantas de sombra, en las que estos niveles podrían causar fotoinhibición, ya que la asimilación neta positiva se logra a niveles de luz mucho menores que los requeridos por las plantas de sol para alcanzar su compensación (Salisbury y Ross, 2000).
Materia seca de hojas
La figura 3 registra el comportamiento de la biomasa de las hojas. Los análisis de varianza realizados 60, 150 y 255 ddt, indicaron que en cada una de estas épocas se presentan diferencias altamente significativas para la interacción genotipo x radiación; la descomposición de la interacción (figura 4) mostró que estas diferencias están explicadas de manera consistente en las tres épocas por el comportamiento diferencial de 'Morita 2' respecto a 'Morita 1', siendo 'Morita 2' la que registra acumulación de materia seca de hojas significativamente mayor cuando es expuesta a 100% de radiación incidente.
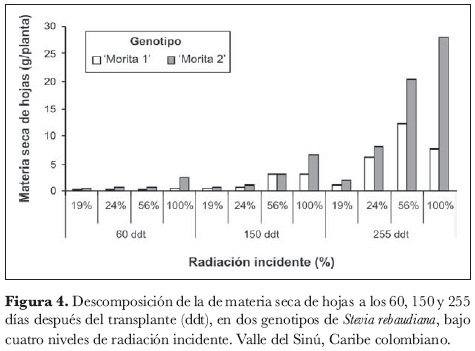
En razón a que los resultados indican que la mayor acumulación de materia seca de hojas se obtuvo cuando 'Morita 2' fue expuesta a 100% de radiación incidente, es posible que este genotipo responda de una manera más eficiente a la fijación de CO2 por unidad de superficie fotosintética y al envío de fotoasimilados a la hoja, respecto a 'Morita 1', lo que sería consistente con la expansión del área foliar, como se evidenció en el punto anterior. La relación entre el área foliar y la materia seca de las hojas muestra una relación positiva directamente proporcional, ya que el aumento en la primera afectó de manera positiva a la segunda. Esta condición es totalmente relevante si se tiene en cuenta que son las hojas los órganos de mayor interés económico de esta planta, pues son las que contienen mayor proporción de glucósidos de diterpeno, responsables de su gran poder edulcorante.
Páez (1997) encontró resultados similares en investigaciones sobre aclimatación de P. maximum, sometida a diferentes regímenes lumínicos (10%, 30% y 100%), reportando que la materia seca de hojas fue superior para el tratamiento expuesto a 100%, con reducciones de 50,1% y 96,4% para los tratamientos expuestos a niveles de radiación de 30% y 10%, respectivamente, en la primera fecha de evaluación, y de 57% y 97,1% en la segunda fecha. Taiz y Zeiger (1998) mencionan que la cantidad de luz y CO2 determinan la respuesta fotosintética de las hojas y que, en algunas situaciones, esta respuesta es limitada por un inadecuado suministro de luz. Aunque esto no siempre es así pues existen otros factores involucrados, éste podría ser el caso para los dos genotipos de estevia, en los que las deficiencias en la cantidad de luz afectaron probablemente de manera importante la capacidad fotosintética de las hojas, respuesta que, según Salisbury y Ross (2000), es muy plástica dependiendo de los recursos disponibles.
De igual manera que para el área foliar, la acumulación de biomasa en las hojas como respuesta a estímulos de variación en la radiación recibida es un factor que se modifica con la especie. Walcroft et al. (2002), en trabajados realizados con melocotón, no encontraron una relación lineal entre la RFA y la biomasa de las hojas. Los autores afirman que la variación en la capacidad de los árboles para hacer fotosíntesis es debida a la plasticidad del dosel para adaptarse a las variaciones de la RFA. En este mismo sentido, Lee y Heuvelink (2003) encontraron que el área foliar específica de hojas nuevas de crisantemo mostró un aumento lineal significante con el inverso de la RFA diaria. Por otra parte, Zermeño et al. (2005) informan que, de acuerdo a mediciones en laboratorio, en la hoja la eficiencia fotosintética es mayor con niveles bajos de radiación debido a que las hojas se saturan de radiación cuando están expuestas directamente al sol, lo que ocurre en todas las hojas de plantas con metabolismo fotosintético C3. Dicen los mismos autores que, aunque las hojas de las plantas del tipo C4 no alcanzan la saturación, la eficiencia de la fotosíntesis también es mayor con niveles bajos de radiación.
Materia seca total del dosel
La figura 5 muestra la biomasa acumulada en el dosel de los genotipos durante el ciclo de desarrollo. La tendencia de la acumulación total de biomasa mostró, de manera general, un comportamiento creciente a medida que el nivel de sombreamiento disminuyó, de manera independiente al genotipo. Sin embargo, al final del ciclo, cuando 'Morita 1' fue expuesta a un 100% de radiación, la biomasa total del dosel decreció respecto al nivel de sombreamiento inmediatamente anterior (56%), sugiriendo que a este nivel ya ha sobrepasado posiblemente su punto de saturación lumínica, a partir del cual los incrementos en la luz no producen aumentos en la fotosíntesis. Es posible, entonces, que la reducción en la acumulación de la biomasa de 'Morita 1' bajo altos niveles de irradiancia sea una consecuencia de los diferenciales entre las tasas de fotosíntesis y respiración, ésta última cada vez más creciente con el aumento de la temperatura y la radiación. Además, es importante considerar que a altas temperaturas -que se registran normalmente en los niveles altos de radiación- la proporción de O2 respecto al CO2 cloroplástico disuelto es mayor que a temperaturas más bajas, de manera que la fijación de O2 sea más rápida e, indirectamente, la fotorrespiración frene el crecimiento de manera diferencial entre 'Morita 1' y 'Morita 2'. Sin embargo, es interesante reseñar que los tratamientos más sombreados (19% y 24%) mantuvieron un registro de biomasa muy inferior con respecto a los niveles de radiación alta, independientemente de la variedad.
Los análisis estadísticos indicaron que sobre la biomasa del dosel existió un efecto importante de la interacción genotipo x radiación a los 60 y 255 ddt (P ≤ 0,05), en tanto que a los 150 ddt, el efecto más importante lo registró el nivel de radiación, registrándose los mayores valores cuando las plantas estuvieron irradiadas en un 100%. El análisis de las interacciones indicó, de manera consistente con la época, que 'Morita 2' presenta valores superiores de materia seca total, respecto a 'Morita 1', cuando está expuesta a 100% de radiación incidente. Esta respuesta de 'Morita 2' es consistente con los resultados del área foliar y de la biomasa de las hojas, sugiriendo que, bajo las condiciones ambientales de la región Caribe, ésta es más eficiente fisiológicamente en la síntesis de asimilados que 'Morita 1'.
Mariscal et al. (2000), en trabajos desarrollados con árboles jóvenes de olivo, informaron que la producción de materia seca total se correlacionó linealmente con la interceptación de la RFA acumulada. Estos resultados muestran una tendencia similar a la hallada en trabajos realizados en lechuga por De Grazia et al. (2001), quienes señalan que las plantas cultivadas bajo 65% de sombra alcanzaron un peso seco total (PST) significativamente menor que el resto. Moreno (1997) encontró que el com-portamiento fotosintético del cuángare (Otoba gracilipes), bajo distintos regímenes de iluminación, presenta las tendencias típicas de las plantas de sombra: su capacidad fotosintética (tasa máxima de asimilación) se reduce sustancialmente cuando crece a alta radiación y se dispara a medida que aumenta el sombreamiento. La tendencia del sajo (Campnosperma panamensis) es opuesta, típica de las especies heliófitas: su capacidad fotosintética aumenta cuando crece bajo radiación alta; además, la fotosíntesis en las hojas que crecen a la sombra se satura bajo valores de radiación menores. Los resultados anteriores corroboran el que las respuestas de los vegetales ante estímulos de luz sean muy variadas y que la alta heterogeneidad de estas respuestas obedezca principalmente a la especie y a otras condiciones ambientales, como temperatura y disponibilidad de agua, entre otras.
Distribución de la materia seca en el dosel
Las tendencias de distribución de la materia seca entre tallos y hojas registró un comportamiento similar para las dos variedades (figura 6). En términos generales, se encontró que al inicio del ciclo las plantas tienden a acumular biomasa en las hojas como un mecanismo de fortalecimiento de su aparato fotosintético. Teniendo en cuenta que son éstas las fuentes de fotoasimilados por excelencia, puede deducirse que, en este período de rápido crecimiento, son las hojas, sobre todo las nuevas, las principales demandantes de estos compuestos, ya que para esta época la planta se encuentra en la fase vegetativa, cuando sólo compiten por las sustancias asimiladas las raíces, los tallos y las hojas. Es probable que el análisis de la biomasa de la raíz pueda dar más claridad en la discusión, pero desafortunadamente este parámetro no fue calculado en este estudio. Resultados similares fueron reportados en habichuela por Jarma et al. (1999), para plantas que crecieron bajo niveles altos y bajos de radiación.
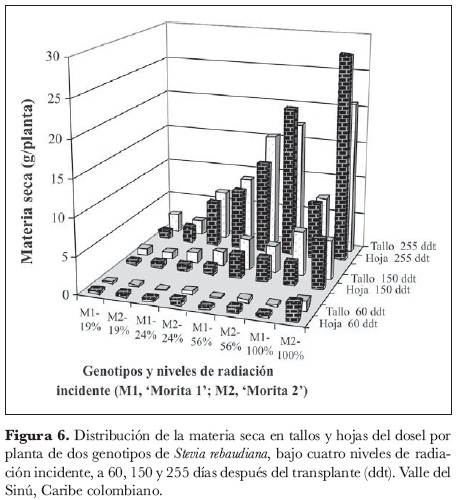
A mediados del ciclo ocurre un cambio notorio en la distribución de la materia seca total para todos los tratamientos, cuando la materia seca del tallo está en mayor proporción que la materia seca de las hojas, indicando que para esta época el tallo se convierte en el principal órgano demandante de fotoasimilados y las hojas pasan a un segundo plano. Por último, se establece una tendencia equilibrada entre tallos y hojas.
En las plantas que crecieron bajo mayores niveles de sombra no es muy claro cuál sea el órgano de demanda predominante, mientras que para los niveles máximos de radiación incidente la materia seca de hojas se presenta en mayor proporción. De acuerdo con lo anterior, es posible presumir que la planta se encarga primero de fortalecer su aparato fotosintético, expandiendo su área foliar, para luego fortalecer su tallo principal y ramas; una vez logrado este objetivo, tiende a establecer un equilibrio en la relación fuente-demanda
Conclusiones
Bajo las condiciones del presente ensayo, el área foliar de la variedad 'Morita 2' fue superior al de la variedad 'Morita 1', independientemente de la radiación, lo que sugiere que aquella variedad posee probablemente características morfofisiológicas y genéticas que le permiten ser más eficiente. Sin embargo, el incremento de la radiación genera incrementos en el área foliar de la especie, siendo más favorables los niveles de radiación de 56% y 100%.
Los altos niveles de radiación inducen un incremento de la materia seca de las hojas y materia seca total en 'Morita 2', efecto que no se evidenció para 'Morita 1'.
La mayor proporción de materia materia seca de las hojas, respecto a la del tallo, durante los primeros 90 ddt indica que la planta durante esta época se dedica a la expansión foliar, fortaleciendo su aparato fotosintético. Después de este tiempo, la migración de los fotoasimilados se da en mayor proporción hacia los tallos y, por último, termina con una tendencia estable hacia ambos órganos de demanda.
Literatura citada
Brandle, J.E., A. Richman, A. Swanson y B. Chapman. 2002. Leaf ESTs from Stevia rebaudiana: a resource for gene discovery in diterpene synthesis. Plant Mol. Biol. 50, 613-622. [ Links ]
Brandle, J.E. 2005. Stevia, nature's natural low calorie sweetener. En: Agriculture and Agri-Food Canada, http://res2.agr.ca/london/faq/stevia_e.htm; consulta: junio 2005. [ Links ]
Conover, C. y R. Flohr. 2003. Light, fertilizer and cultivar selection affect growth and yield of containerized patio tomatoes. Research Report RH-96-1. Commercial Follage Research Reports (CFRR). University of Florida Research and Education Center, Apopka, 81. [ Links ]
De Grazia, J., P. Tittonell y A. Chiesa. 2001. Efecto de la época de siembra, radiación y nutrición nitrogenada sobre el patrón de crecimiento y el rendimiento del cultivo de lechuga (Lactuca sativa L.). Invest. Agr. Prod. Prot. Veg. 16(3). [ Links ]
Galván, L. y J. Guzmán. 2003. Determinación de los requerimientos nutricionales de Stevia rebaudiana Bert. en el Sinú Medio. Trabajo de grado. Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). 106 p. [ Links ]
Geuns, J.M. 2003. Stevioside. Phytochemistry 64, 913-921. [ Links ]
Jarma, A., C. Buitrago y S. Gutiérrez. 1999. Respuesta del crecimiento de la habichuela (Phaseolus vulgaris L. var. Blue Lake) a tres niveles de radiación incidente. Revista Comalfi 26(1-3), 62-73. [ Links ]
Lee, J. y E. Heuvelink. 2003. Simulation of leaf area development based on dry matter partitioning and specific leaf area for cut chrysanthemum. Ann. Bot. 91(3), 319-327. [ Links ]
Mansab A., D. Jeffers y P. Henderlong. 2003. Interrelationship between leaf area, light interception and growth rate in a soybeanwheat system. Asian J. Plant Sci. 2181, 605-612. [ Links ]
Mariscal, M., F. Orgaz y F. Villalobos. 2000. Radiation-use efficiency and dry matter partitioning of a young olive (Olea europaea) orchard. Tree Physiol. 20, 65-72. [ Links ]
Midmore, D.J. y A.H. Rank. 2002. A new rural industry -Stevia- to replace imported chemical sweeteners. RIRDC Report 02/022, 55 p. [ Links ]
Moreno, F. 1997. Fotosíntesis en plántulas de sajo (Campnosperma panamensis) y cuángare (Otoba gracilipes) bajo diferentes ambientes lumínicos. Revista Crónica Forestal y del Medio Ambiente 12(1). [ Links ]
Páez, A., M. González y J. Villasmil. 1997. Acclimation of Panicum maximum to different light regimes: effect of subsequent defoliation. Rev. Fac. Agron. (UZ) 14, 625-639. [ Links ]
Páez, A., M. González, J. Urdaneta, D. Paredes, D. Tissue y T. Tschaplinski. 1998. Índices de crecimiento y formación de compuestos orgánicos en Barleria lupulina sometida a dos condiciones de luminosidad. Rev. Fac. Agron. (UZ) 15, 515-525. [ Links ]
Paytas, M. 2005. Evolución del índice de área foliar en distintas densidades y distancias de siembra en el cultivo de algodón. En: Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Reconquista, Argentina http://www.inta.gov.ar/reconquista/info/documentos/agricultura/agric_tecnica/info_indice_area_foliar_algodon.pdf. pp. 1-5; consulta: octubre 2005. [ Links ]
Rahimian, H. y E. Zand. 2004. Canopy profile distribution of leaf area, light and nitrogen in some Iranian winter wheat cultivars released during the last 50 years. En: Proc. 4th International Crop Science Congress. Brisbane, Australia. 26th sept. - 1st oct. 2004. [ Links ]
Rengifo, T. 2003. Análisis de crecimiento de Stevia rebaudiana Bertoni bajo cuatro niveles de radiación incidente en el valle del Sinú. Tesis de maestría. Convenio Universidad Nacional de Colombia y Universidad de Córdoba. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 216 p. [ Links ]
Salisbury, F.B. y C.W. Ross. 2000. Fisiología de las plantas. 2. Bioquímica vegetal. Ed. Paraninfo, Madrid. 523 p. [ Links ]
Schwebel, R. 2005. Stevia, el edulcorante natural sudamericano con cero calorías. En: http://www.fm.unt.edu.ar/ftp/Health_I_G_News/Dic1998.doc; consulta: junio 2005. [ Links ]
Secretaría de Agricultura de Antioquia. 2000. Informe preliminar sobre adaptación de la especie Stevia rebaudiana en la región tropical. Gobernación de Antioquia, Medellín (Colombia). 25 p. [ Links ]
Taiz, L. y E. Zeiger. 1998. Plant physiology. Second edition. Sinauer Associates, Sunderland, MA. pp. 228-229. [ Links ]
Tewolde H., K. Sistani, D. Rowe, A. Adeli y T. Tsegaye. 2005. Estimating cotton leaf area index nondestructively with a light sensor. Agron. J. 97, 1158-1163. [ Links ]
Totté, N., L. Charon, M. Rohmer, F. Compernolle, I. Baboeuf y J.M.C. Geuns. 2000. Biosynthesis of the diterpenoid steviol, an entkaurene derivative from Stevia rebaudiana Bertoni, via the methylerythritol phosphate pathway. Tetrahedron Letters 41, 6407-6410. [ Links ]
Totté, N., W. van den Ende, E.J.M. van Damme, F. Compernolle, I. Baboeuf y J.M.C. Geuns. 2003. Cloning and heterologous expression of early genes in gibberellin and steviol biosynthesis via the methylerythritol phosphate pathway in Stevia rebaudiana. Can. J. Bot. 81, 517-522. [ Links ]
Universidad Eafit. 2004. Inteligencia de mercados internacionales de Stevia rebaudiana. Departamento de Negocios Internacionales, Medellín (Colombia). 71 p. [ Links ]
Walcroft A., X. Le Roux, A. Díaz-Espejo, N. Dones y H. Sinoquet. 2002. Effects of crown development on leaf irradiance, leaf morphology and photosynthetic capacity in a peach tree. Tree Physiol. 22(13), 929-938. [ Links ]
Zermeño, A., J. Montemayor, J. Munguía, L. Ibarra y M. Cadena. 2005. Reflectividad y absortibidad de la radiación en tres densidades de planta y su relación con el rendimiento de maíz (variedad Cafime). Agrociencia 39, 285-292. [ Links ]

