










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.23 no.2 Bogotá July/Dez. 2005
Influencia de la fertilización foliar con Ca sobre la pudrición apical en tomate (Lycopersicon esculentum Mill.)
Influence of the foliar fertilization Ca on the blossom-end rot (BER) in tomato (Lycopersicon esculentum Mill.)
Carlos Cardona1, Harvey Arjona2 y Hermes Araméndiz-Tatis3
1 Profesor titular, Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). e-mail: ccardona@sinu.unicordoba.edu.co
2 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. e-mail: hearjona@unal.edu.co
3 Profesor asociado, Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). e-mail: haramendiz@hotmail.com
Fecha de recepción: 01 de febrero de 2005. Aceptado para publicación: 21 de noviembre de 2005
Resumen
Uno de los problemas en la producción de tomate (Lycopersicon esculentum Mill.), en campo abierto o en invernadero, es la pudrición apical de los frutos asociada con la deficiencia localizada de calcio. Para evaluar el efecto de la aplicación foliar de Ca sobre la pudrición apical y la calidad de los frutos de tomate, se aplicaron tres concentraciones de quelato de calcio: 0,05%, 0,10% y 0,15% e calcio activo, aplicados una, dos y tres veces, según las combinaciones de tratamientos, a los 5, 10 y 15 d post-antesis, en el área experimental de la Universidad de Córdoba (Unicor) y en la finca Ucrania, localidades del municipio de Montería (Colombia). Se utilizó un diseño completamente aleatorizado con arreglo factorial 3 x 3, un control y cuatro repeticiones. En Unicor no se presentó el desorden fisiológico, pero el efecto del Ca se manifestó al incrementarse la firmeza y la concentración de Ca en los frutos, mientras que los sólidos solubles totales, el pH y la acidez no fueron afectados consistentemente por los tratamientos. En Ucrania el efecto se hizo evidente al reducir la incidencia del desorden a niveles mínimos, en tanto que las características físicas y químicas de los frutos no fueron afectadas por los tratamientos, con la notable excepción del contenido de Ca en frutos. Éste resultó altamente significativo en ambas localidades, de tal manera que la relación funcional entre el Ca aplicado y el Ca en frutos puede explicarse con modelos cuadráticos.
Palabras claves adicionales: desorden fisiológico, deficiencia, nutrición mineral, quelato.
Abstract
Blossom-end rot is a major problem in tomato (Lycopersicon esculentum Mill.) production and it has been associated with Ca deficiency. Three Ca concentrations (0.05%, 0.10% and 0.15%) were evaluated at three different times (5, 10 and 15 days after anthesis) for evaluating the effect of Ca sprays on blossom-end rot and fruit quality in tomatoes. A complete randomised design was used with four replicates for each treatment and two repetitions in two different localities (Universidad de Córdoba [Unicor] and Ucrania at Montería, Colombia). The results revealed that there was no physiological disorder in locality 1 (Unicor) but an effect of the Ca sprays was observed on the fruit texture (increased firmness) and fruit Ca content. No effect was observed on soluble solids, pH and acidity. The disorder was seen to be reduced in locality 2 (Ucrania) when Ca sprays were used; however, physical and chemical characteristics were not affected except for the amount of Ca found in the fruit. The results suggest that the functional relationship between Ca sprays and amount of Ca in the fruit can be explained using quadratic models.
Additional key words: physiological disorder, deficiency, mineral nutrition, chelation.
Introducción
EL TOMATE DE MESA (LYCOPERSICON ESCULENTUM MILL.) es el fruto de mayor consumo a nivel mundial. La producción de tomate bajo cubierta y en campo abierto ha crecido significativamente durante la última década en muchos países, entre ellos Colombia, con los consecuentes aumentos en la demanda de los conocimientos relativos a su sistema de producción (Arjona et al., 1998). Uno de los principales problemas en la producción de tomate es la pudrición apical de los frutos asociada con deficiencia de calcio (Lazcano, 2000). En el departamento de Córdoba se han reportado pérdidas de la producción de frutos del orden de 43% (Boada, 1998).
Varios desórdenes fisiológicos de las plantas, caracterizados por el colapso de células y necrosis de los tejidos, se han relacionado con la deficiencia localizada de Ca. La pudrición apical en tomate presenta una necrosis progresiva de grupos de células, con la subsiguiente deshidratación parcial del área afectada. El desorden comienza en el fruto inmaduro, durante la etapa de rápido crecimiento (Shanon et al., 1996).
Las zonas meristemáticas de raíces, tallos y hojas, en donde la división celular es más activa, son más susceptibles a la deficiencia de Ca, quizá debido a que éste se requiere para formar una nueva lámina media en el fragmoplasto entre las células hijas y para satisfacer las necesidades de las membranas en formación (Salisbury y Ross, 1994).
La planta de tomate con deficiencia de Ca presenta hojas nuevas con márgenes necróticas y, en plantas jóvenes, las hojas se doblan hacia arriba formando una copa. Además, se reduce la tasa de crecimiento y las partes nuevas de la planta no crecen. La punta de la raíz muere y la radícula se ramifica. El síntoma más conocido de la deficiencia de Ca en tomate es la pudrición apical de la fruta y obedece al incremento en la demanda de Ca para la rápida expansión celular, haciendo deficitario el suministro de Ca hacia los tejidos susceptibles del fruto (Marcelis y Ho, 1999; Lazcano, 2000).
La cantidad de Ca que llega al fruto durante el período de rápida expansión celular es más importante que la cantidad de Ca que toma la planta. El período crítico se presenta aproximadamente dos semanas después de la antesis, cuando la tasa de crecimiento del fruto es mayor (Ho et al., 1993).
En el tomate, la susceptibilidad de una variedad a la pudrición pistilar está directamente relacionada con la cantidad de tejido xilemático desarrollado en la parte distal del fruto y con la tasa de expansión del fruto. El transporte del Ca disminuye con la distancia al pedúnculo, de manera que los tejidos de la parte distal siempre contienen menos Ca que los tejidos proximales, al tener la parte distal del fruto menos haces vasculares y menor relación xilema/ floema (Ho et al., 1993; Marcelis y Ho, 1999).
Un fruto de tomate es 95% agua, y más de 85% de esa agua proviene de la savia floemática. De aquí se desprende el que el aporte de agua al fruto vía xilema sea muy pequeño, pero es el único canal por el cual se mueve el Ca hacia el fruto. De otro lado, menos de 3% del Ca absorbido por el cultivo va a los frutos, a pesar de que el crecimiento de los frutos representa 90% del crecimiento del cultivo (Ho et al., 1993).
A plena exposición solar, una gran cantidad de Ca se dirige hacia las hojas. En éstas la tasa de transpiración es más alta que en los frutos. Sólo en la noche, cuando no hay transpiración o es reducida, una proporción de flujo xilemático incrementada se desvía hacia los frutos. Así, la transpiración se da a gran velocidad con luz brillante, en particular si la humedad ambiental es baja. Estas condiciones estimulan el crecimiento de la fruta pero no el suministro de Ca. Por otra parte, las altas conductividades eléctricas reducen el ascenso y el transporte de Ca hacia los frutos (Ho et al., 1993).
El Ca debe estar por encima de 0,12% para evitar el riesgo de la aparición de la pudrición apical. El 70% del Ca total de la planta se retiene en las hojas, mientras los frutos sólo contienen un 5% y, a diferencia del potasio, la translocación del Ca al fruto, una vez asimilado por las hojas, es muy escasa (Chamarro, 1995).
Las aplicaciones de Ca realizadas en precosecha son muy utilizadas, aunque es necesario hacer varias a lo largo de las etapas de crecimiento para obtener buenos resultados, lo que implica mayor costo (Perring, 1979; Glenn y Poovaiah, 1985). Sin embargo, el riesgo de dañar las hojas y frutos limita la concentración de sales que puede utilizarse en cada aplicación (Mason, 1976; Johnson, 1979).
Por otro lado, las aspersiones de Ca sobre hojas y frutos permiten uniformizar la distribución del Ca en el fruto, ya que ésta es desigual, disminuyendo progresivamente de la parte proximal hacia la parte distal (Ho et al., 1993).
Este estudio tuvo como finalidad evaluar la influencia de la aspersión de Ca activo sobre la incidencia de la pudrición apical y su efecto en la calidad de los frutos de tomate.
Materiales y métodos
El trabajo se realizó en el área experimental de la Facultad de Ciencias Agrícolas de la Universidad de Córdoba (en adelante, área Unicor) y en la finca Ucrania (corregimiento de Buenos Aires - La Manta), ambas localidades situadas en el municipio de Montería, departamento de Córdoba (Colombia), entre junio de 2001 y junio de 2002.
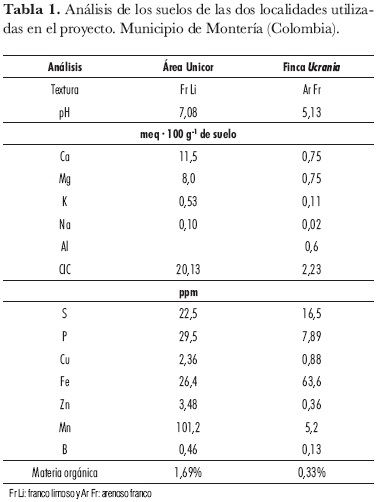
Los resultados de los análisis correspondientes a los suelos del área Unicor y de la finca Ucrania se muestran en la tabla 1.
El área experimental en cada localidad se sembró con la variedad de tomate 'Chonto Santacruz Kada', la más demandada por los cultivadores de tomate en la región y susceptible a la pudrición apical. Se evaluaron los efectos de tres concentraciones de quelato de Ca: 0,05%, 0,10% y 0,15% de Ca activo (120 g de Ca quelatado por kg de CaO), aplicados una, dos y tres veces, según las combinaciones de tratamientos, a los 5, 10 y 15 d post-antesis, sobre el follaje y frutos en la etapa de desarrollo, correspondiente al tercer racimo. Se utilizó un diseño completamente aleatorizado con arreglo factorial 3 x 3, un control y cuatro repeticiones. Se consideraron como variables de respuesta, además de la incidencia de pudrición apical, las características físicas y químicas de los frutos: masa fresca, firmeza, acidez, sólidos solubles totales y contenido de Ca. El cultivo se fertilizó de acuerdo con sus requerimientos y se implementó un manejo integrado de plagas y enfermedades.
Para determinar la firmeza (kg· 0,505 cm-2), se tomaron dos lecturas de cada fruto, cinco frutos por unidad experimental, con un penetrómetro (Bertuzzi FT 011) con punta simétricamente cónica (diámetro 0,80 cm y longitud 2,10 cm). El pH se midió con un potenciómetro digital (Metrohm 692); los sólidos solubles (en grados Brix), utilizando un refractómetro Atago N-1EBX y la acidez titulable, mediante el empleo de NaOH 0,1 N y fenolftaleína como indicador, expresando los resultados en porcentaje de ácido cítrico. El Ca total se determinó en frutos completos, después de mineralización húmeda, y se cuantificó por espectrofotometría de absorción atómica (Perkin Elmer 3110).
Para cada experimento se hizo el análisis de varianza correspondiente, así como contrastes ortogonales entre las medias de los tratamientos.
Resultados y discusión
Incidencia de la pudrición apical
El desorden fisiológico no se presentó en ninguna de las plantas sometidas a los tratamientos con Ca ni en las de control cultivadas en el área Unicor. La no aparición del desorden fisiológico obedeció a la presencia de condiciones ambientales favorables para el cultivo, como humedad relativa alta y brillo solar bajo que, asociados con niveles altos de Ca intercambiable (11,4 meq· 100 g-1 de suelo), conductividad eléctrica de 0,49 dS· m-1 y riego adecuados, favorecieron la toma de Ca y su translocación hacia los frutos, en concordancia con Sha et al. (1985), Maroto et al. (1995) y Paiva et al. (1998), quienes reportan que las condiciones climáticas no favorables al desorden fisiológico facilitan el transporte de Ca hacia los frutos, ya que este elemento parece incapaz de moverse por caminos simplásticos. Es claro que cuando se presenta irradiancia, temperatura alta y humedad relativa baja, la transpiración aumenta, provocando pérdidas excesivas de agua y soluto y desfavoreciendo el transporte de Ca hacia los frutos.
Por otro lado, en suelos con altas conductividades eléctricas, los frutos de tomate no importan el Ca esencial suficiente para la apropiada formación de la pared celular cuando las células de los frutos se están dividiendo rápidamente, ni cuando hay déficit hídrico. De esta forma, los frutos del tomate son más propensos a sufrir resquebrajamiento y necrosis en el área distal (Ho et al., 1993).
En contraste con la situación anterior, la pudrición apical se observó en la finca Ucrania en 5,9% de las plantas y en 3,1% de los frutos al iniciarse el cuajado de los primeros frutos, es decir, en los racimos correspondientes a la primera inflorescencia. Los frutos presentaron, en diferentes etapas de desarrollo, áreas necróticas en su parte apical.
En esta localidad se podría relacionar la aparición del desorden fisiológico con los contenidos de Ca bajos (0,75 meq· 100 g-1 de suelo) y, en general, de bases presentes en este tipo de suelo de textura arenosa-franca. La combinación de contenido de Ca bajo y permeabilidad alta del suelo, que en algún momento causó estrés hídrico, influyó desfavorablemente en la toma de Ca y su translocación hacia los frutos. Además, los valores de brillo solar fueron relativamente altos, registrándose una media de 7,10 h-sol durante los diez primeros días de crecimiento del fruto, que, según Ho et al. (1993), junto con la temperatura alta provoca una transpiración alta que ocasiona un flujo elevado de nutrientes hacia las hojas, en lugar de ir hacia los frutos.
Adicionalmente, las condiciones ambientales favorecieron un crecimiento acelerado de los frutos, situación que, según Ho et al. (1993) y Marcelis y Ho (1999), provoca un incremento en la demanda de Ca para la rápida expansión celular, haciendo insuficiente su suministro en los tejidos de la parte distal del fruto.
En el piso de producción evaluado (inflorescencia tres), post-aplicación foliar de Ca, la incidencia de pudrición apical se observó en el control, con una media de 1,23% ± 0,21% y, en menor grado, en el tratamiento correspondiente a la concentración de Ca más baja, aplicada una sola vez, con 0,45% ± 0,08%, disminuyendo notablemente en las plantas tratadas respecto a la incidencia observada al iniciarse el cuajado de los primeros frutos, lo que puede atribuirse, en parte, al efecto del Ca aplicado. Esta situación corrobora lo reportado por Borkorski (1984) y Ho (1999), quienes manifiestan que la aspersión de Ca al fruto de tomate en la etapa crítica del desarrollo, cuando crece con mayor rapidez, es efectiva para disminuir y prevenir la incidencia de pudrición apical. Con un mínimo de cuatro aspersiones de cloruro o nitrato de Ca al fruto, cada 3-5 d, se logra disminuir en cerca de seis veces el número de frutos afectados por pudrición apical (Borkorski, 1984).
Peso fresco de frutos
Los frutos presentaron en todos los tratamientos valores medios de peso fresco muy similares. El valor medio fue 66,07 ± 7,46 g en el área Unicor y 67,79 ± 9,99 g en la finca Ucrania.
El análisis de varianza no detectó diferencias significativas entre tratamientos en las dos localidades (P > F = 0,2222 y P > F = 0,3536). Esto sugiere, de algún modo, que la acumulación de biomasa es similar en todos los tratamientos, incluyendo el control, y no varía en función del Ca aplicado.
Firmeza de los frutos
Los frutos provenientes del área experimental Unicor presentaron valores medios de 3,06 ± 0,49 kg· 0,505 cm-2, mientras que para los de la finca Ucrania fueron 3,19 ± 0,61 kg· 0,505 cm-2. La tabla 2 presenta los valores de firmeza o consistencia de frutos sometidos a los tratamientos con Ca.
El análisis de varianza del experimento localizado en el área Unicor no reportó diferencias entre las medias de los tratamientos (P > F = 0,2828), mientras que el de Ucrania mostró diferencias significativas entre tratamientos (P > F = 0,0303). Aunque este atributo importante de calidad de los frutos de tomate parece estar bajo control genético (Al-Falluji et al., 1982), es evidente que varía en función del Ca aplicado.
La interacción concentración por número de aplicaciones (CCNL) resultó significativa, indicándose así que los efectos de la concentración de Ca sobre la firmeza cambian según el número de aplicaciones. En consecuencia, al resultar significativa la interacción CCNL (P > F = 0,0165), es posible conceptuar que el efecto cuadrático de la concentración de Ca cambia lineal-mente al cambiar los niveles del número de aplicación, o sea que la curvatura de la concentración de Ca en una aplicación es diferente a la correspondiente en tres aplicaciones (figura 1).
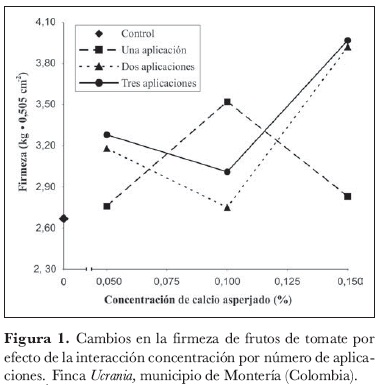
La prueba de Duncan para los diferentes promedios, desde el punto de vista de la concentración de Ca (comparación vertical utilizando α = 0,05), permite afirmar que el Ca, cuando se aplica una vez, no presenta efectos significativos; cuando se aplica dos veces, aumenta la firmeza, con la concentración de 0,15%, y finalmente, cuando se aplica tres veces, la respuesta es positiva, lográndose el mayor valor con el nivel correspondiente a 0,15%. Esto puede ser agronómicamente positivo para la no presencia o disminución de la pudrición apical en tomate y para el manejo postcosecha, en especial, en lo relacionado con manipulación, transporte, almacenamiento y mercadeo (Al-Falluji et al., 1982). Esta situación concuerda con los aumentos de firmeza reportados en otras especies, por ejemplo, en manzanas 'Golden delicious' (Poovaiah, 1988; Drake y Spayd, 1983) y en durazno 'Flordaprince' (Baez et al., 2000).
Concentración de calcio
Los frutos procedentes del área Unicor presentaron valores medios de 0,2617% ± 0,0406%, mientras que en la finca Ucrania la concentración de Ca fue 0,211% ± 0,041%.
El análisis de varianza permitió observar diferencias altamente significativas entre tratamientos en ambas localidades (P > F = 0,0001). Los contrastes ortogonales evidencian que los tratamientos con diferentes concentraciones del Ca asperjado superaron en conjunto al control, con una significancia alta (P > F = 0,0001). Este resultado sugiere que el Ca fue absorbido por la parte aérea de la planta y particularmente por los frutos y corrobora lo encontrado para la firmeza de fruto.
A través de comparaciones ortogonales, las respuestas lineal y cuadrática resultaron altamente significativas (P > F = 0,0001, para ambos efectos. Al resultar altamente significativos los efectos lineal y cuadrático de la concentración de Ca, la curva parabólica presenta una tendencia general de ascenso (figura 2).
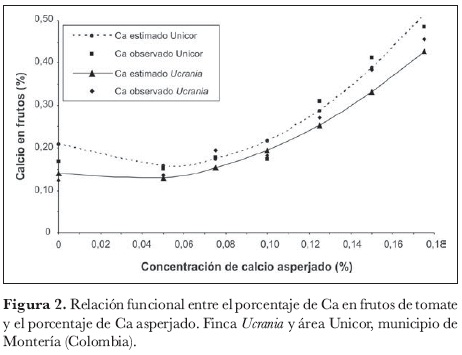
Según la ecuación estimada, se puede afirmar que, en el área Unicor por cada 0,01% de incremento de la concentración de Ca asperjado, el porcentaje de Ca en los frutos disminuye 2,105% con efecto lineal y aumenta 21,987% con efecto cuadrático, con errores estándar de 0,641% y 3,667%, respectivamente (figura 2).
Para la finca Ucrania, se afirma que, por cada 0,01% de incremento de la concentración de Ca asperjado, el porcentaje de Ca en los frutos disminuye 0,909% con efecto lineal y aumenta 14,63% con efecto cuadrático, con errores estándar de 0,582% y 3,33%, respectivamente (figura 2).
Báez et al. (2000) observaron los efectos de la aplicación foliar de Ca sobre su concentración endógena en el fruto de durazno, sin evidenciarse ninguna tendencia en la concentración de Ca en relación con el tipo de producto utilizado, la dosis o el número de aplicaciones realizadas.
Si bien las aspersiones foliares incrementaron el contenido de Ca en los frutos, tal aumento está influencia-do posiblemente por las condiciones climáticas, el contenido de Ca en el suelo y la disponibilidad de agua en él. Es notable la diferencia encontrada en los controles de las dos localidades: 0,195% y 0,119%, para Unicor y Ucrania, respectivamente.
Wada et al. (1996) observaron incrementos en la concentración de Ca en las hojas por efecto de la aplicación foliar de Ca sobre plantas de tomate. Con un mínimo de cuatro aspersiones de cloruro o nitrato de Ca cada 3-5 d, se ha incrementado hasta dos veces el contenido de Ca en los frutos de tomate (Borkorski, 1984).
Sólidos solubles totales
En el área experimental Unicor, el valor medio de los sólidos solubles totales (SST) resultó ser 5,03 ± 0,38 °Brix, mientras que en la finca Ucrania correspondió a 5,09 ± 0,36 °Brix. Mucha atención se ha dado a este componente de la calidad porque, no sólo afecta el sabor, sino también el rendimiento industrial del tomate para procesamiento (Vallejo, 1999).
Los valores de los SST presentaron pequeñas variaciones entre tratamientos, sin que se detectaran diferencias significativas entre ellos. Esto sugiere que los SST están presentes en cantidades muy similares en los frutos, como consecuencia del mismo estado de madurez en que fueron cosechados, y no varían en función de los tratamientos aplicados ni de la etapa de desarrollo de la planta; es decir, no existe un efecto aparente del Ca en las evaluaciones realizadas, confirmándose lo reportado por Wada et al. (1996), quienes, al evaluar el efecto de aplicaciones foliares de Ca en tomate, no encontraron diferencias en los SST frente al testigo. Estos valores variaron entre 5,4 ± 0,07 ºBrix y 6,1 ± 0,36 ºBrix.
Un resultado similar fue reportado por Báez et al. (2000) en durazno, al aplicar diferentes fuentes de Ca de una a cuatro veces; el contenido de sólidos solubles en los frutos tratados no presentó ninguna tendencia y los valores determinados no fueron diferentes del correspondiente al control.
Acidez
En el área experimental de Unicor, el valor medio de la acidez del fruto del tomate resultó ser 0,38% ± 0,06%, mientras que en la finca Ucrania correspondió a 0,42% ± 0,05%.
El análisis de varianza no reportó diferencias significativas entre tratamientos en las dos localidades. No existe pues un efecto del Ca en el porcentaje de ácidos orgánicos presentes en los frutos de tomate. Un resultado similar lograron Wada et al. (1996) al evaluar el efecto de la aplicación foliar de Ca en tomate; los tratamientos con Ca registraron valores entre 0,336% y 0,376%, mientras que el control registró 0,427%.
La media de acidez resultó más alta en la finca Ucrania que en el área Unicor (0,42% vs. 0,38%). Esta diferencia indica la presencia de una mayor reserva de ácidos orgánicos en las vacuolas de las células de los frutos procedentes de Ucrania, que puede ser movilizada para su empleo por las mitocondrias como sustratos oxidables en el ciclo de los ácidos tricarboxílicos (Wills et al., 1984).
Durante la maduración, los ácidos orgánicos son oxidados o convertidos en azúcares. Pueden ser considera-dos como una reserva energética más de la fruta, y se espera que su contenido decline en el período de actividad metabólica máxima durante el curso de la maduración (Wills et al., 1984). En consecuencia, es posible que la diferencia en la acidez de los frutos encontrada en las dos localidades se deba al período de actividad metabólica presente en el momento de la recolección y no, al efecto del Ca absorbido.
Conclusiones
• La incidencia de pudrición apical encontrada en los frutos de tomate estuvo relacionada con condiciones climáticas favorables a la aparición del desorden fisiológico, a deficiencia de Ca en el suelo y a estrés hídrico.
• La aplicación foliar de Ca resultó eficiente al disminuir la incidencia de pudrición apical en los frutos del tomate y al incrementar su firmeza, característica importante que influye en su resistencia potencial al transporte.
• La relación funcional entre el Ca aplicado y el Ca en frutos se explica bajo un modelo cuadrático.
• Las aspersiones de Ca no tuvieron efecto diferencial sobre el peso fresco, los sólidos solubles totales y la acidez de los frutos de tomate.
Literatura citada
Al-Falluji, R.A., D.H. Trinklein y V.N. Lambeth. 1982. Inheritance of pericarp firmness in tomato by generation mean analysis. HortScience 17(5), 763-764. [ Links ]
Arjona, H., H. Escobar y G. Herrera. 1998 Evaluación de la propagación vegetativa de tomate (Lycopersicon esculentum Mill.) mediante esquejes en la Sabana de Bogotá. Revista Comalfi 30(1), 9-12. [ Links ]
Báez, R., R. Troncoso, E. Bringas, J. Ojeda y A. Mendoza. 2000. Efectos de diferentes fuentes de Ca en melocotones. pp. 63-77. En: Memorias del Congreso iberoamericano de tecnología postcosecha y agroexportaciones. Bogotá [ Links ].
Boada, C. 1998. Determinación del efecto de la aplicación de Ca sobre la calidad y el rendimiento del tomate (Lycopersicon esculentum Mill.). Trabajo de grado. Facultad de Ciencias Agropecuarias, Corporación Universitaria de Ciencias Aplicadas y Ambientales (UDCA), Bogotá. 66 p. [ Links ]
Borkowski, J. 1984. Study on the calcium uptake dynamic by tomato fruits and blossom-end rot control. Acta Hort. 145, 72-75. [ Links ]
Chamarro, J. 1995. Anatomía y fisiología de la planta. pp. 83-84. En: Nuez, F. (ed.). El cultivo del tomate. Edición Mundi-Prensa, Madrid. [ Links ]
Drake, S.R. y S.E. Spay. 1983. Influence of calcium treatment on 'Golden Delicious' apple quality. J. Food Sci. 48, 403-405. [ Links ]
Glenn, G. y B. Poovaiah. 1985. Cuticular permeability to calcium compounds in 'Golden Delicious' apple fruit. J. Amer. Soc. Hort. Sci. 110(2), 166-171. [ Links ]
Ho, L., R. Belda, M. Brown, J. Andrews y P. Adams. 1993. Uptake and transport of calcium and the possible causes of blossomend rot in tomato. J. Exp. Bot. 44(259), 509-518. [ Links ]
Ho, L.C. 1999. The physiological basis for improving tomato fruit quality. Acta Hort. 487, 1-3. [ Links ]
Johnson, D. 1979. New techniques in the postharvest treatment of apple with calcium salts. Soil Sci. Plant Anal. 10(12), 373-382. [ Links ]
Lazcano, I. 2000. Deficiencia de Ca en tomate (Lycopersicon esculentum Mill). Informaciones Agronómicas, Instituto de la Potasa y el Fósforo de Canadá, 39, 7-8. [ Links ]
Marcelis, L. y L. Ho. 1999. Blossom-end rot relation to growth rate and calcium content in fruits of sweet pepper (Capsicum annum L.). J. Exp. Bot. 50(332), 357-367. [ Links ]
Maroto, J.V., S. López, A. Bardisi, B. Pascual y J. Alargada. 1995. Influence of irrigation dosage and its form of application on cracking response in cherry tomato fruits. Acta Hort. 379, 181-185. [ Links ]
Mason, J. 1976. Calcium concentration and firmness of stored 'Macintosh' apples increased by calcium chloride solution plus thicker. J. Science 11(5), 504. [ Links ]
Paiva, E., R. Arruda-Sampaio y H. Prieto. 1998. Composition and quality of tomato fruit cultivated in nutrient solutions containing different calcium concentrations. J. Plant Nutr. 21(12), 2653-2661. [ Links ]
Perring, M. 1979. The effects of environment and cultural practices on calcium concentration in the apple fruit. Soil Sci. Plant Anal. 10(1-2), 279-293. [ Links ]
Poovaiah, B.W. 1998. Molecular and cellular aspects of calcium action in plants. HortScience 23(2), 267-271. [ Links ]
Salisbury, F y C. Ross. 1994. Fisiología vegetal. Cuarta edición. Grupo Editorial Iberoamérica, México, DF. pp. 3-177. [ Links ]
Sha, L., B. McCown y L. Peterson. 1985. Occurrence and cause of shoot-tip necrosis in shoot cultures. J. Amer. Soc.Hort. Sci. 110(5), 631-634. [ Links ]
Shanon, S., J. Natti y D. Atkin. 1996. Relation of calcium nutrition to hypocotil necrosis of snap bean (Phaseolus vulgaris L.). New York State Agricultural Experiment Station for Publication as Journal Paper (1536). [ Links ]
Vallejo, F. 1999. Mejoramiento genético y producción de tomate en Colombia. Editorial Feriva, Cali (Colombia). pp. 52-53. [ Links ]
Wada, T., M. Ikeda y H. Furukawa. 1996. Effects of foliar calcium solutions on incidence of blossom-end rot of tomato fruit. J. Japan. Soc. Hort. Sci. 65(3), 553-558. [ Links ]
Wills, R.H.H., T.H. Lee, W.B. McGlasson, E.G. Hall y D. Graham. 1984. Fisiología y manipulación de frutas y hortalizas post-recolección. Editorial Acribia, Zaragoza. pp. 19-41. [ Links ]

