










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.24 no.2 Bogotá July/Dec. 2006
Ángela María Bernal1, Jorge Enrique Arias2, José Dilmer Moreno3, Iván Valbuena4 y Luis Ernesto Rodríguez5
1 Ingeniera agrónoma, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
e-mail: ambernalva@unal.edu.co
2 Ingeniero agrónomo, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
e-mail: jeariasr@unal.edu.co
3 Investigador, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Centro de Investigación Tibaitatá, Mosquera (Colombia).
e-mail: jdmoreno@corpoica.org.co
4 Investigador, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Centro de Investigación Tibaitatá, Mosquera (Colombia).
e-mail: ri.valbuena@yahoo.com
5 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
e-mail: lerodriguezmo@unal.edu.co
Resumen:
Dentro de una colección de germoplasma, la presencia de materiales duplicados amenaza su identidad genética, enmascarando su variabilidad y aumentando los costos de manejo y mantenimiento. Por tal razón, con el fin de identificar los materiales candidatos a duplicados dentro de la Colección Central Colombiana de papa Solanum tuberosum ssp. andigena, se llevó a cabo la caracterización morfológica con variables cualitativas. La investigación se realizó durante el primer y segundo semestre de 2004 en el Centro de Investigación Tibaitatá de la Corporación Colombiana de Investigación Agropecuaria (Corpoica). El análisis factorial de correspondencias múltiples (ACM) permitió identificar los caracteres color primario y secundario de piel de tubérculo, color secundario de carne de tubérculo y color primario y secundario de flor, como los que mejor discriminan la colección, y mediante el análisis de conglomerados, se obtuvo un dendrograma que permitió definir siete grupos y cuatro subgrupos en los que se identificaron los posibles duplicados bajo un índice de nivel arbitrario que admitiera rigor estadístico y amplitud biológica. En total, se candidatizaron 64 materiales para una confirmación molecular posterior.
Palabras claves adicionales: recursos genéticos, germoplasma, conglomerados
Abstract:
Presence of duplicated material in a germplasm core collection can threat its genetic identity, hide its real variability and increase the cost of management and maintenance. Because of this reason, with the aim of identifying candidate materials for duplicates in Colombian Center Collection of potato Solanum tuberosum ssp. andigena, the morphologic characterization with qualitative variables was carried out. The investigation was made during the first and second semester of 2004, in Corpoica CI Tibaitatá (Mosquera, Cundinamarca). The multiple correspondence analysis (ACM) permited to identify characters of primary and secondary tuber skin color, secondary tuber meat color, and primary and secondary flower color, as the ones that better discriminated the collection and, by means of the cluster analysis, a dendrogram was obtained, which defined seven groups and four subgroups, from which the possible duplicates under an arbitrary level index were identified that admitted statistic rigor and biologic amplitude. Totally were recognized 64 materials for a later molecular confirmation.
Additional key words: genetic recourses, germplasm, conglomerates
Introducción
LA DIVERSIDAD GENÉTICA DE LA PAPA se concentra en la zona Andina de Suramérica, que incluye un amplio rango de hábitat que va desde ambientes muy fríos y sujetos a heladas hasta condiciones muy secas de semidesierto; este hecho ha colaborado con la adaptación de estos recursos al estrés abiótico y con el desarrollo de resistencias a varias plagas y enfermedades, mostrando cómo los recursos genéticos constituyen un reservorio de adaptabilidad genética que actúa como un cojinete frente a los efectos potencialmente dañinos del ambiente y los cambios económicos (Bonierbale et al., 2004).
La estructura taxonómica de la papa es muy compleja; Hawkes (1990) nomina más de 200 especies entre silvestres y cultivadas y ocho especies cultivadas en la zona Andina, mientras que Huamán y Spooner (2002) no reconocen tales especies como independientes sino como unas pocas o una sola especie, siendo S. stenotomum el grupo más primitivo y S. tuberosum ssp. andigena la papa cultivada más importante de los Andes, mientras que, en la región litoral del sur de Chile, la papa cultivada más importante pertenece a la especie tetraploide S. tuberosum ssp. tuberosum.
En Colombia, la Colección Central de papa hace parte del Sistema nacional de bancos de germoplasma, a cargo de Corpoica; concentra gran parte de la variabilidad genética de la especie y está constituida por 664 accesiones de la subespecie andigena, 83 de la subespecie tuberosum, además de un grupo importante de materiales silvestres e introducciones provenientes de Estados Unidos, Alemania, Holanda y los países andinos Bolivia, Perú, Ecuador y Venezuela (Corpoica, 2004).
El aprovechamiento de los recursos genéticos implica la disponibilidad del material, su debida caracterización y evaluación morfológica, agronómica y molecular, relaciones taxonómicas dilucidadas y el estudio completo de su variabilidad (Hidalgo, 2003). En este sentido, la Colección se encuentra caracterizada morfológicamente de acuerdo con los descriptores para la caracterización básica de colecciones de papa del Centro Internacional de la Papa CIP-1994 (Ligarreto et al., 1997); por precocidad, azúcares reductores y gravedad específica (Avella y Parra, 1997) y por resistencia a gota Phytophthora infestans (Mont) de Bari (Méndez y Rojas, 1997).
El tamaño de la colección, analogías entre los datos de pasaporte, el tipo de propagación por tubérculo-semilla y el manejo y manipulación del material son factores que favorecen la presencia de duplicados, que, en su conjunto, enmascaran la variabilidad real de la colección, estrechan su base genética y amenazan la identidad que se pretende conservar.
Siguiendo los principios básicos del fenetisismo a nivel operacional, se pueden establecer similitudes al interior de un grupo de unidades (poblaciones, especies, individuos) al ser comparadas entre sí con la ayuda de metodologías estadísticas, legítimamente suscritas a las técnicas del análisis multivariado de datos (Crisci y López, 1983; Díaz, 2002). Estas metodologías precisan la ejecución de dos grandes grupos de procedimientos: ordenación (análisis de correspondencias múltiples) y agrupamiento (análisis de conglomerados). Con la ordenación se busca reducir el número original de variables a un grupo pequeño y compacto y con el agrupamiento se propone repartir los individuos en grupos claramente delimitados, excluyentes o no, según un esquema jerárquico o no jerárquico, respectivamente (Crisci y López, 1983; Crossa et al., 1985; Hidalgo, 2003).
La identificación propia de duplicados está en función de la valoración de los caracteres genéticos de una población o accesión. Para establecer los elementos comparativos y definir la situación de duplicados, se deben realizar todas las valoraciones necesarias, caracterizaciones y evaluaciones, en función de los objetivos de la preservación del recurso. El procedimiento, en el sentido estricto de la genética de poblaciones, consiste en encontrar ciertas similitudes o igualdades entre dos accesiones, aunque no sean exactamente duplicados (Chávez, 2003).
La posibilidad de detectar estos duplicados tendría, no sólo un efecto directo en la reducción del costo que genera el mantenimiento de la colección en campo, sino que representaría una contribución al conocimiento de las relaciones taxonómicas entre accesiones y de su valor potencial en el aprovechamiento genético de la subespecie.
El objetivo de este trabajo fue detectar los materiales que sean posibles duplicados, a través de una caracterización morfológica analizada estadísticamente mediante un análisis factorial de correspondencias múltiples y, posteriormente, un análisis de conglomerados.
Materiales y métodos
Como materiales se utilizaron 464 accesiones de la especie Solanum tuberosum ssp. andigena conservadas ex situ en la CCC, debidamente identificadas con el código asignado en el Banco de germoplasma. El trabajo se desarrolló durante 2004 en el Centro de Investigación Tibaitatá de Corpoica (Mosquera, Cundinamarca). Se sembraron 10 plantas de cada una de las accesiones a 0,45 m entre plantas en surcos sencillos de 4,5 m de longitud.
La caracterización morfológica se basó en los descriptores de papa (Solanum tuberosum) para caracterización básica de colecciones propuestos por el Centro Internacional de la Papa (CIP, 1994). Se evaluaron 24 caracteres cualitativos categóricos correspondientes a los diferentes órganos de la planta (tabla 1). Las variables seleccionadas para este trabajo fueron todas cualitativas por ser las que tienen menor influencia ambiental (Vallejo y Estrada, 2002).
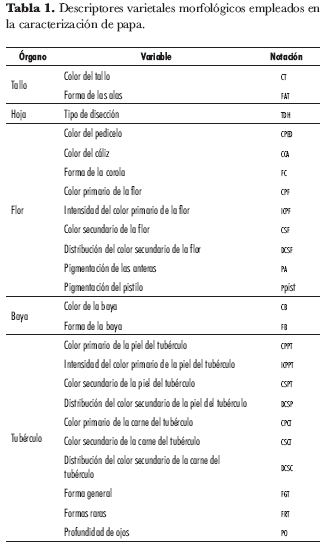
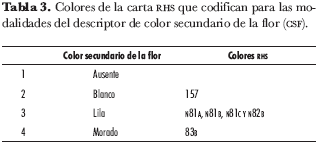
El color primario de flor se tomó con la carta de colores de la Royal Horticultural Society (RHS). Se identificaron 30 tonalidades diferentes, que se asociaron en nueve grupos que describen color e intensidad y codifican dentro de las modalidades de los descriptores de los caracteres color primario e intensidad de color primario de la flor (tabla 2); así mismo, para color secundario se observaron 6 tonalidades, asociadas en cuatro grupos, incluyendo ausencia de color secundario (tabla 3).

Los datos para caracteres de tallo, hoja, flor y baya fueron observados en tres plantas por accesión; para datos de tubérculo, la información se tomó de 20 tubérculos y en ambos casos se registraron las modas.
El análisis para la matriz de datos (completa para 316 accesiones) se dividió en dos etapas: 1) análisis de correspondencias múltiples (en adelante, ACM) y 2) análisis de conglomerados (AC). El ACM encuentra asociaciones entre variables de tipo categórico a través de las respectivas modalidades de éstas (Díaz, 2002) y busca establecer el valor discriminatorio de los caracteres con respecto a las relaciones que se definan entre las unidades bajo estudio (Crisci y López, 1983). A su vez, el AC se propone definir la estructura natural de un conjunto de objetos, partiéndolo en grupos, de tal forma que los objetos de un mismo grupo sean similares y los objetos de grupos diferentes sean disímiles (Díaz, 2002).
El análisis para el caso de variables categóricas múltiples (ACM) considera la nube de puntos fila (n puntos) y la nube de puntos columna (p puntos). En el primer caso, los ejes son las variables y en el segundo, los individuos. De acuerdo a Lebart et al. (1995), los principios del ACM son: transformar la tabla de datos en perfiles fila y perfiles columna; ajustar los datos o puntos ponderados por sus perfiles marginales (fila o columna) y dar la distancia entre perfiles por la ji-cuadrado.
La distancia ji-cuadrado (χ2) pondera las distintas coordenadas de manera que se le da más ‘importancia’ a las categorías con menor frecuencia y menos ‘importancia’ a las que tengan alta frecuencia (Díaz, 2002).
Las respectivas distancias ji-cuadrado, entre modalidades y entre individuos, aplicada a una tabla disyuntiva completa6 están dadas por:
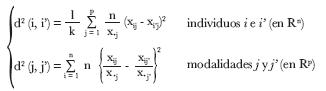
donde xij, observación para el individuo i-ésimo de la modalidad j-ésima; k, número total de variables; n, número total de individuos; p, número total de modalidades; x.j = ∑i=1 xij y R, espacio vectorial sobre los números reales.
Mediante el paquete estadístico SPAD versión 4.51, se procesaron los datos y se obtuvieron los factores, de tal forma que cada uno explica una parte de la asociación total entre variables y las coordenadas métricas para cada accesión (por convención cada accesión se nota como un individuo).
En la clasificación de individuos (a través del análisis de conglomerados), el sistema de análisis de datos SPAD utiliza algoritmos de construcción de grupos que trabajan con las coordenadas factoriales de un análisis previo (Morineau, 1994), en este caso, con las coordenadas resultantes del ACM, y utiliza el criterio de agregación de Ward, cuya principal ventaja es que minimiza la varianza dentro de grupos y maximiza la varianza entre grupos (Franco et al., 1997a; Crossa et al., 1995).
Resultados y discusión
Las variables tipo de disección de hoja (TDH) y pigmentación de anteras (PA) fueron constantes para las modalidades de ‘hoja diseccionada’ y ‘sin antocianinas’, respectivamente, hecho que permitió excluirlas del análisis; según Crisci y López (1983), es preciso excluir los caracteres que no varían en las unidades bajo estudio, ya que carecen de poder discriminatorio y, por lo tanto, de valor taxonómico.
De otra parte, la variable forma rara de tubérculo (FRT) fue asumida como una variable suplementaria, pues siempre apareció complementando la información de la variable forma general de tubérculo (FGT), más común que la primera, que tuvo una baja frecuencia de aparición.
El ACM arrojó 56 ejes o factores que reúnen el 100% de la varianza. La varianza ligada a cada valor propio estuvo entre 0,1948 y 0,0015 para el valor propio más grande y el más pequeño, respectivamente. De la misma forma, el primer valor propio explica un 6,96% de la variación total y el segundo, un 5,11%, mientras que el décimo valor propio explica el 2,79% de la varianza total (tabla 4). Esto coincide con la naturaleza del ACM, pues en éste, a diferencia del análisis de componentes principales, los primeros ejes, aun en forma creciente, explican una pequeña parte de la variabilidad total (Lebart et al., 1995).
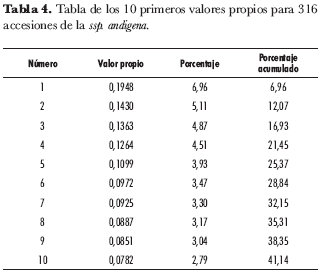
Al explorar en detalle las contribuciones absolutas de todos los ejes factoriales para cada variable, se encontró que a partir del décimo eje las variables con mayor contribución se empezaron a repetir, hecho que permitió seleccionar de los primeros 10 ejes las variables que mejor discriminan la ssp. andigena. En la tabla 5 se presentan las contribuciones de cada variable para la construcción de los primeros 10 factores. Los ejes que se omitieron contienen valores máximos de contribuciones para variables que resultaron significativas en factores anteriores.
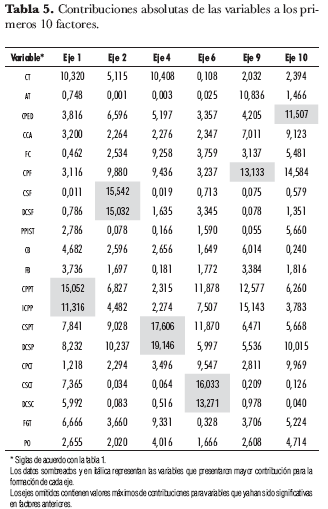
Las variables que discriminan la CCC de papa (S. tuberosum ssp. andigena) son, en orden de importancia: color primario de piel de tubérculo y su intensidad, color secundario de flor y su distribución, color secundario de piel de tubérculo y su distribución, color secundario de carne de tubérculo y su distribución, color primario de flor y color de pedicelo. Estas variables corresponden parcialmente con lo encontrado por Ligarreto et al. (1997), quienes proponen las variables color primario de flor y color primario y secundario de piel de tubérculo dentro de las más significativas. Esta correspondencia fue evidente con lo observado en campo, al menos al nivel de estas dos variables; sin embargo, el análisis estadístico incluyó variables cuya ingerencia no fue fácilmente percibida al momento de hacer las observaciones: es el caso de los caracteres color secundario de flor y color secundario de carne del tubérculo. Esto se explica en gran medida por la naturaleza del método de análisis utilizado, que precisa la ponderación de los valores (Díaz, 2002); así, la moda registrada para estas dos variables (tabla 6) permitiría diferenciar este grupo de n individuos en dos grandes grupos alternativos: aquéllos con color secundario de flor blanco y aquéllos que no presentan esta característica, o aquéllos con color secundario de carne de tubérculo violeta y aquéllos que carecen de este carácter.
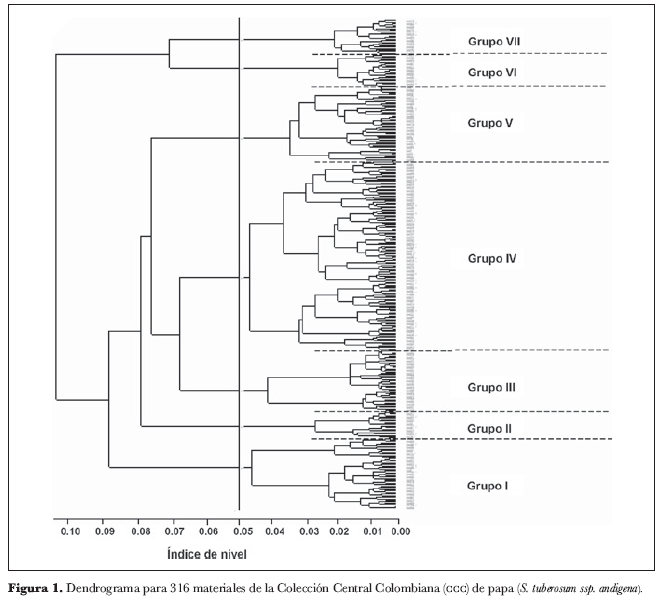
Los valores propios no nulos fueron p - k = 55 (número de modalidades - variables activas) (Jobson, 1992), cuyas coordenadas de individuos se utilizaron para la construcción del dendrograma (figura 1). Al interior de éste último se discriminaron siete grupos y cuatro subgrupos, de acuerdo al índice de nivel7 en que cada uno de ellos se diferenció del resto; en el índice 0,10697 se diferenciaron los grupos 6 y 7 del resto; en 0,09004, el grupo 1; en 0,08010, el grupo 2; en 0,07660, el grupo 5 de los grupos 3 y 4; en 0,07074 se diferenció el grupo 6 del grupo 7 y finalmente en el nivel de 0,06771 lo hizo el grupo 4 del 3.
Aunque se dispone de un amplio número de estrategias para decidir sobre el número de conglomerados para construir, en este trabajo el criterio decisivo ha sido la homogeneidad ‘media’ alcanzada dentro de los conglomerados (Díaz, 2002), ya que en muchas situaciones existen grupos ‘naturales’, separados unos de los otros, para ser descritos por el esquema de clasificación; además, muchas de las clasificaciones dependen del grado de refinamiento de la diferenciación, y la estructura general obedece más a un árbol complejo, con ramas a diferentes ‘alturas’ (como ocurre en los dendrogramas de análisis jerárquico) (Hampel, 2002).
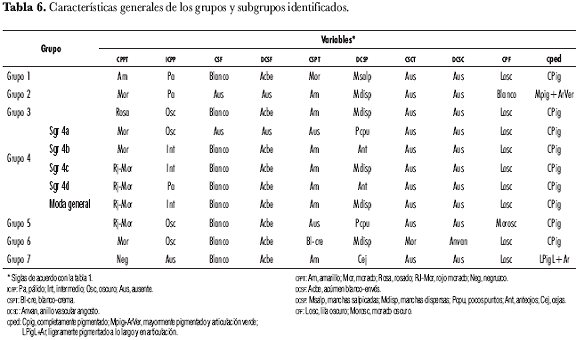
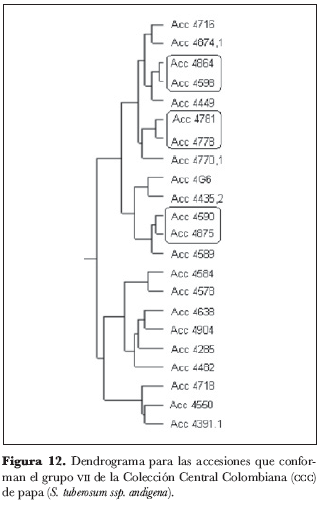
Sin embargo, la decisión sobre el número de grupos para considerar es, por lo general, de la incumbencia del especialista asociado con el estudio en consideración (Díaz, 2002). La selección de forma arbitraria de siete grupos y cuatro subgrupos en el dendrograma (figura 1) obedece a: (i) la percepción global al hacer un examen visual del esquema; (ii) la homogeneidad que se evidencia al interior de los conglomerados y (iii) la identificación de los duplicados que se ve facilitada; esta designación arbitraria también entra en juego a la hora de establecer el criterio para la elección de los duplicados. La polémica radica en la imposibilidad de determinar en qué nivel dos materiales son equivalentes, similares o iguales (Chávez, 2003). Las características generales de cada grupo se presentan el la tabla 6 y corresponden a las modas de las variables más discriminatorias (modalidad más frecuente).
Las figuras 2-12 presentan la ubicación jerárquica a mayor escala de las accesiones; los grupos se describen de derecha a izquierda –o bien, de abajo hacia arriba– en el dendrograma completo. En un dendrograma se pueden identificar tanto núcleos como grupos; los primeros no son más que las parejas formadas por los individuos más cercanos, mientras que un grupo es el producto de la integración de núcleos con accesiones individuales o con otros núcleos, en índices de nivel mayores (Crisci y López, 1983); de esta manera, se propusieron como candidatos a duplicados todos los núcleos y grupos de máximo tres accesiones por debajo de un índice de nivel de 0,00248, designado arbitrariamente. En este nivel, se tiene un eje por encima del cual no sería sensato postular candidatos a las pruebas moleculares –en aras de la precisión– y por debajo se estarían excluyendo materiales potencialmente duplicados; es decir, se buscó aquel punto que diera oportunidad al mayor número de eventos favorables posibles.
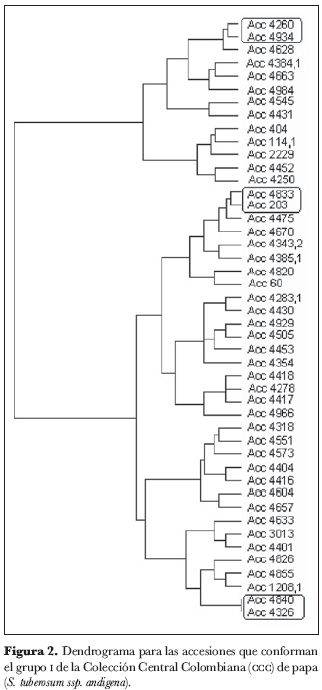
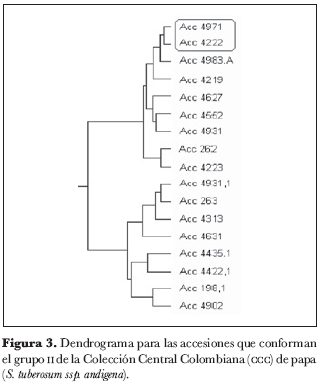
Dentro del grupo 1 (figura 2) se identificaron los núcleos 4934 y 4260, 4833 y 203, 4840 y 4326; en el grupo 2 (figura 3), el núcleo 4971 y 4222; en el grupo 3 (figura 4), los núcleos 4810 y 4237, 4899 y 4852, 4369 y 4765, 4220 y 4671, 4388 y 4624, 4906 y 4714 y el grupo 4905, 4937 y 4480.
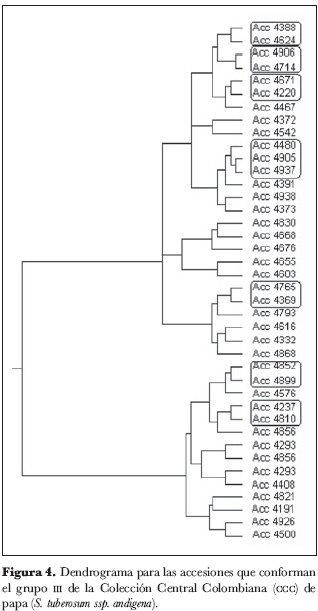
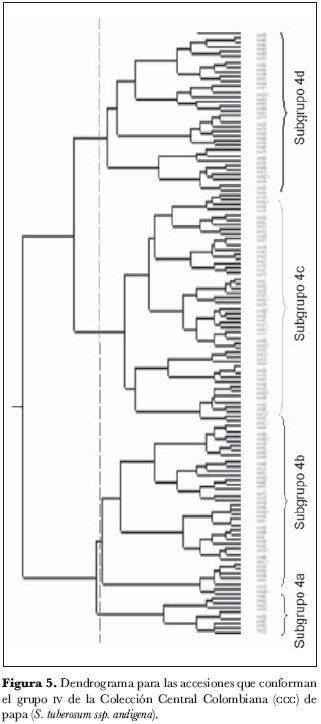
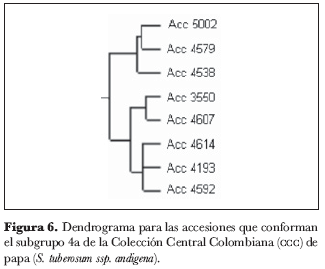
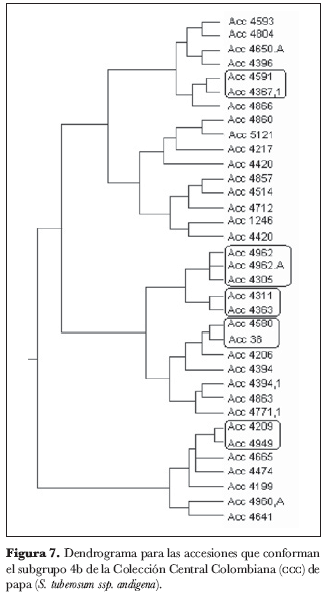
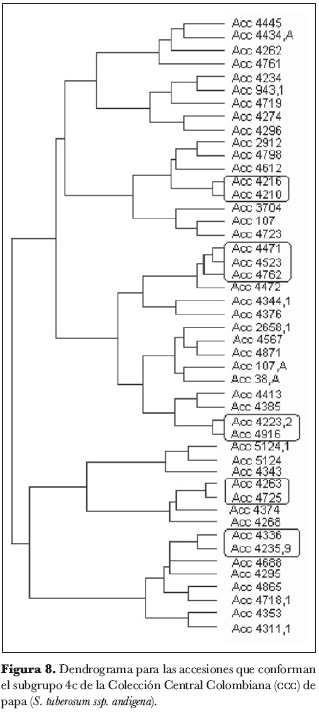
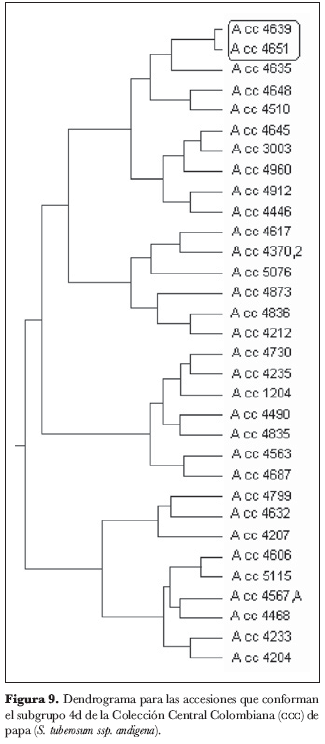
El grupo 4 (figura 5) se subdividió en cuatro subgrupos: 4a, 4b, 4c y 4d. En el subgrupo 4a (figura 6) no se identificaron núcleos ni grupos; el subgrupo 4b (figura 7) está conformado por los núcleos 4209 y 4949, 38 y 4580, 4363 y 4311, 4367.1 y 4591 y por el grupo 4305, 4962.A y 4962; del subgrupo 4c (figura 8) forman parte los núcleos 4235.9 y 4636, 4725 y 4263, 4916 y 4223.2, 4210 y 4216 y el grupo 4471, 4523 y 4762; finalmente, en el subgrupo 4d (figura 9) aparece el núcleo 4651 y 4639.
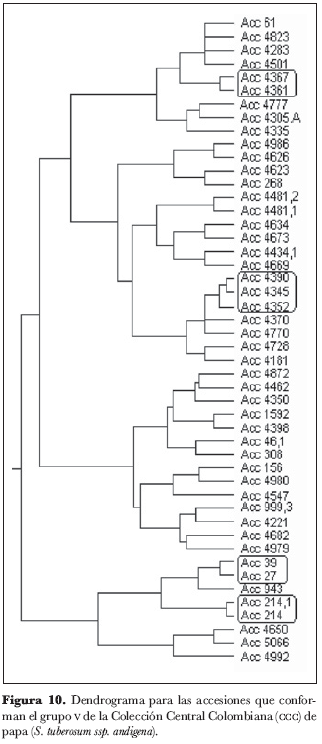
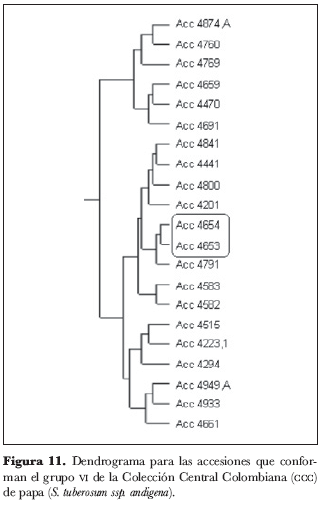
En el grupo 5 (figura 10) se encontraron los núcleos 214 y 214.1, 27 y 39, 4367 y 4361 y el grupo 4390, 4345 y 4352; el único núcleo del grupo 6 (figura 11) está conformado por las accesiones 4653 y 4654, así como los tres núcleos del grupo 7 (figura 12) están conformados por las accesiones 4875 y 4590, 4778 y 4781, 4598 y 4864.
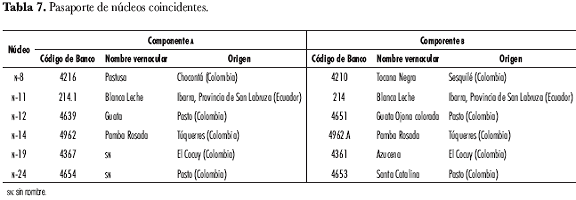
La postulación de materiales duplicados se basó en el análisis estadístico de la caracterización; sin embargo, los datos de origen y consecutivos de los códigos designados por el Banco fueron registrados para confirmar la hipótesis de igualdad de materiales, resultando que seis de los núcleos se relacionaron desde su origen mismo (tabla 7). Estos núcleos que coincidieron en sus datos de pasaporte constituyeron un fuerte argumento de respaldo a la postulación de 64 materiales como candidatos a duplicados, desde el punto de vista morfológico; no obstante, es necesario someter estos materiales a pruebas moleculares antes de tomar una decisión sobre su descarte o permanencia en la colección.
Conclusiones
En este trabajo se logró: la identificación de los caracteres cualitativos que mejor discriminan la CCC de papa S. tuberosum ssp. andigena a nivel morfológico; la definición de un grupo importante de materiales candidatos a duplicados y el avance en el conocimiento de la estructura morfológica de este acervo.
Para trabajos posteriores se propone seguir conservando los candidatos a duplicados hasta que la evidencia molecular no demuestre lo contrario y, en cuanto a las accesiones que no se incluyeron en este trabajo, se sugiere completar la caracterización morfológica para su posterior inclusión en alguno de los grupos definidos y, una vez consolidados estos grupos, hacer estudios más específicos dentro de cada uno de ellos, tales como la evaluación y caracterización del material, incluyendo variables morfológicas, agronómicas, fisiológicas e industriales.
6 Una tabla disyuntiva completa es una matriz cuyas columnas están compuestas por variables presencia-ausencia (1,0), representando las categorías (pi) de las k variables.
7 El índice de nivel es un valor de disimilaridad derivado de la matriz de distancias, al cual se presentan diferentes niveles de agregación y está representado gráficamente en el eje horizontal de un dendograma.
Literatura citada
Avella, A. y Parra, H. 1997. Evaluación de la Colección Central Colombiana de papa (Solanum tuberosum ssp. andigena) por los parámetros de precocidad y calidad industrial en el centro de investigaciones Tibaitatá, municipio de Mosquera. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 74 p. [ Links ]
Bonierbale, M., W. Amoros, J. Espinoza, E. Mihovilovich, W. Roca y R. Gómez. 2004. Recursos genéticos de la papa: don del pasado, legado para el futuro. Suplemento Revista Latinoamericana de la Papa 2004. [ Links ]
Chávez, J.L. 2003. Análisis estadístico de datos de caracterización morfológica, pp. 72-77. En: Franco, T. y R. Hidalgo (eds.). Análisis estadístico de datos de caracterización morfológica de recursos fitogenéticos. Boletín técnico IPGRI No 8. International Plant Genetics Resources Institute (IPGRI). Cali, Colombia. 89 p. [ Links ]
Centro Internacional de la Papa [CIP]. 1994. Descriptores de la papa para caracterización básica de colecciones nacionales. Centro Internacional de la Papa (CIP), Lima, Perú.
Corporación Colombiana de Investigación Agropecuaria [Corpoica]. 2004. Informe anual 2003. Documento inédito. Programa de recursos genéticos y biotecnología vegetal, Corporación Colombiana de Investigación Agropecuaria (Corpoica).
Crisci, V. y M.F. López. 1983. Introducción a la teoría y práctica de la taxonomía numérica. Secretaría General de la Organización de los Estados Americanos (OEA), Programa regional de desarrollo científico y tecnológico. Washington D.C. 132 p. [ Links ]
Crossa, J., I.H. DeLacy y S. Taba. 1995. The use of multivariate methods in developing a core collection, pp. 77-92. En: Hodgking, T., A.H.D. Brown, Th.J.L. van Hintum y E.A.V. Morales (eds.). Core collections of plant genetic resourses. International Plant Genetics Resources Institute (IPGRI). Cali, Colombia. 269 p. [ Links ]
Díaz, L.G. 2002. Estadística multivariada: inferencia y métodos. Facultad de Ciencias, Universidad Nacional de Colombia. 529 p. [ Links ] Franco, J., J. Crossa, J. Villaseñor, S. Taba y S.A. Eberhart. 1997a. Classifying Mexican maize accessions using hierarchical and density search methods. Crop Sci. 37, 972-980. [ Links ]
Hampel, F. 2002. Some thouthgs about classification. pp. 5-26. En: Jajuga, K., A. Sokolowski y H. Bock (eds.). Classification, clustering and data analysis. Springer-Verlag. 492 p. [ Links ]
Hawkes, J.G. 1990. The potato-evolution, biodiversity and genetic resources. Belhaven Press, Oxford. 259 p. [ Links ]
Hidalgo, R. 2003. Variabilidad genética y caracterización de especies vegetales. pp. 2-26. En: Franco, T. y R. Hidalgo (eds.). Análisis estadístico de datos de caracterización morfológica de recursos fitogenéticos. Boletín técnico IPGRI No 8. International Plant Genetics Resources Institute (IPGRI). Cali, Colombia. 89 p. [ Links ]
Huamán, Z. y D.M. Spooner. 2002. Reclassification of landrace populations of cultivated potato (Solanum sect. Petota). Amer. J. Bot. 89,947-965. [ Links ]
Jobson, J.D. 1992. Applied multivariate data analysis, volume II: categorical and multivariate methods. Springer-Verlag, New York. 768 p. [ Links ]
Lebart, L., A. Morineau y M. Piron. 1995. Statistique exploratoire multidimensionnelle. Dunod, Paris. 439 p. [ Links ]
Ligarreto, G., I. Valbuena y D. Fajardo, D. 1997. Caracterización morfológica de la colección central colombiana de papa. En: Memorias. V Congreso Sociedad colombiana de fitomejoramiento y producción de cultivos. [ Links ]
Méndez-Heredia, R. y L. Rojas-Sepúlveda. 1997. Evaluación de la Colección Central Colombiana de Solanum tuberosum ssp andigena por resistencia a Phytophthora infestans y calidad industrial en la localidad de San Jorge. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 56 p. [ Links ]
Morineau, A. 1994. Análisis de correspondencia. Simposio de estadística. CISIA (Francia) y Facultad de Ciencias, Departamento de Matemáticas y Estadística, Universidad Nacional de Colombia, Bogotá. 67 p. [ Links ]
Vallejo, F.A. y E.I. Estrada. 2002. Mejoramiento genético de plantas. Universidad Nacional de Colombia, Palmira. 67 p. [ Links ]


{kind=link}