










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.24 n.2 Bogotá jul./dez 2006
SUELOS, FERTILIZACIÓN Y MANEJO DE AGUAS
Martha Viviana Torres1 y Luz Marina Lizarazo2
1 Bióloga, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja.
e-mail: vitoce@starmedia.com
2 Profesora asociada, Facultad de Ciencias Básicas, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja.
e-mail: lulizarazo@tunja.uptc.edu.co
Resumen:
La comunidad microbiana de los suelos agrícolas es un componente con múltiples funciones esenciales en la fertilidad del suelo. En este trabajo se hace un estudio de las poblaciones de bacterias implicadas en los ciclos biogeoquímicos del C, N y P, con el fin de evaluar su presencia en cultivos de cebolla (Allium ampeloprasum) y papa (Solanum tuberosum), así como también, de determinar posibles interacciones entre estos grupos, tanto en la rizosfera como el suelo total. Además, se realizó la determinación de la actividad enzimática de la fosfatasa ácida. La evaluación de los grupos funcionales se realizó mediante recuento en placa y la actividad de la fosfatasa ácida, utilizando el método de Tabatabai y Bremner (1969, 1977). Se encontró un mayor número de unidades formadoras de colonia (ufc) en el suelo correspondiente al cultivo de cebolla. Las bacterias solubilizadoras de P se encuentran en menor proporción que los grupos funcionales del ciclo del C y mantienen tamaños poblacionales similares a las bacterias fijadoras de N, tanto en rizosfera como en suelo total en las dos muestras de suelo. Según los resultados, la diferencia en la proporción de microorganismos entre rizosfera y el suelo circundante puede variar por las labores de adición de fertilizantes. Adicionalmente, se determinó que la actividad de la fosfatasa ácida pudo estar afectada por factores como el porcentaje de materia orgánica, la biomasa bacteriana y la disponibilidad de P.
Palabras claves adicionales: fosfatasa ácida, celulíticas, amilolíticas, proteolíticas, bacterias libres fijadoras de nitrógeno, solubilizadoras de fósforo
Abstract:
Microbial community of agricultural soils is a component with multiple functions that are essential for soil fertility. This research was undertaken in order to study bacteria populations participating in the biogeochemical cycles of C, N, and P, with the purpose of evaluating their presence in the cultivation systems and determining possible interactions among these groups in the rhizosphere and total soil. In a similar way, determination of the enzymatic activity of the acid phosphatase was done. The evaluation of the functional groups was carried out by recount in plate and estimation of the acid phosphatase activity using the method of Tabatabai and Bremner (1969, 1977). The phosphate solubilizing bacteria were present in smaller proportion than functional groups of the cycle of C and, in two soils, maintained population sizes similar to that of free-living nitrogen fixing bacteria in rhizosphere and total soil. According to the results obtained in these agricultural soils, the difference in the proportion of microorganisms between rhizosphere and surrounding soil can vary because of the fertilizer additions. Also, it was determined that activity of the acid phosphatase in these two soils could be affected by such factors as percentage organic matter, bacterial biomass, and phosphorus availability.
Additional keywords: acid phosphatase, celulolitics, amilolitics, proteolitics, free-living nitrogen fixing bacteria, solubilizers of phosphorus
Introducción
EL SUELO ES UN RECURSO NATURAL sin estándares de calidad definidos; por su variabilidad, es difícil establecer alguna medida física, química o biológica que pueda ser considerada lo suficientemente adecuada para reflejar la calidad, pues es necesario considerar todos los factores que pueden condicionar su funcionamiento, dificultándose así la regulación de la calidad de este recurso. Es necesario entonces medir aquellas propiedades del suelo que sean especialmente sensibles a los cambios generados por las prácticas de manejo y cuyos valores reflejarían la calidad del recurso (Bandick y Dick, 1999).
Según las diferencias en la exudación de las raíces y de la rizodeposición en diferentes zonas de la raíz, las comunidades microbianas de la rizosfera pueden variar en su estructura o en relación al tipo de suelo, especies de plantas, estado nutricional, edad, estrés, enfermedad y otros factores ambientales (Osorio y Pérez, 2001).
Las propiedades biológicas del suelo, como la biomasa, la actividad de microorganismos y la actividad enzimática, entre otras, podrían constituir una herramienta valiosa para establecer la calidad biológica de un suelo, siendo estas propiedades dependientes de la dinámica de la materia orgánica, las prácticas de manejo y los cambios ambientales (Bandick y Dick, 1999).
En este sentido se pueden utilizar grupos funcionales de organismos como bioindicadores para evaluar disturbios en el suelo o la fertilidad a nivel biológico (Andrade, 2004b). Los grupos funcionales considerados en este estudio son aquéllos que participan directa o indirectamente en los ciclos de los nutrientes y en la recirculación de la materia orgánica, teniendo en cuenta que éstos interaccionan entre sí, con las plantas y los demás factores físicos y químicos del suelo.
La actividad enzimática en los suelos cultivados también está relacionada con las prácticas de cultivo y puede asociarse de una manera directa con la disponibilidad de los nutrientes del suelo, debido a que éstas juegan un papel importante en cada uno de los ciclos biogeoquímicos de elementos requeridos para el crecimiento de las plantas.
Las fosfatasas detectadas en el suelo constituyen un grupo de enzimas de gran importancia en la dinámica de uno de los nutrientes más importantes para las plantas, como lo es el fósforo (P), pues participan en la mineralización del P orgánico, constatando su importancia si se tiene en cuenta que cerca de 90% del P del suelo esta presente en forma orgánica (Portilla et al., 1998).
Ahora bien, la disponibilidad de este elemento para las plantas también se puede relacionar con otra variable biológica importante como es la población de microorganismos solubilizadores del P, pues mediante la liberación de enzimas y ácidos orgánicos liberan formas de este elemento asimilables para las plantas (Osorio y Pérez, 2001).
El objetivo de este estudio fue medir algunas variables biológicas del suelo, como la actividad de la enzima fosfomonoesterasa ácida, la población de bacterias solubilizadores del P y otros grupos funcionales del suelo, tales como amilolíticos, proteolíticos, celulolíticos y fijadores libres de N, en dos suelos cultivados con cebolla y papa (rizosfera y suelo total); con el fin de establecer las diferencias que se puedan presentar en estas variables en dos suelos agrícolas.
Materiales y métodos
El estudio se hizo en condiciones de campo y laboratorio. El trabajo de campo se llevó a cabo en un cultivo de cebolla junca (Allium ampeloprasum) y otro de papa (Solanum tuberosum), con una extensión aproximada de 2 ha cada uno. En cada cultivo se tomaron 24 muestras, utilizando transectos en zig-zag, con una distancia de puntos de muestreo de 1,5 m, según metodología de Créptin y Johnson (2000). Las muestras, de aproximadamente 1 kg, fueron tomadas tanto en rizosfera (12 muestras) como en suelo total (12 muestras), se llevaron al laboratorio y se preservaron a 4 ºC hasta su análisis, que no fue posterior a 48 h.
Las áreas muestreadas están ubicadas en los municipios de Sogamoso y Aquitania (Boyacá), pertenecen a la provincia de Sugamuxi y son zonas que presentan una alta actividad agrícola. Sogamoso está localizado a una altitud de 2.400 msnm, con temperatura promedio de 17 ºC, humedad relativa promedio de 71% y precipitación promedio anual de 831 mm y Aquitania, a orillas del lago de Tota, a 3.046 msnm y con temperatura promedio de 12 ºC, humedad relativa promedio multianual de 82% y precipitación anual promedio de 910 mm.
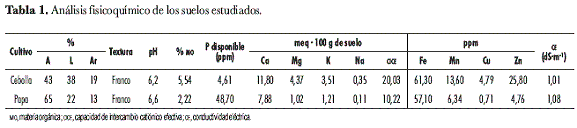
El análisis fisicoquímico de los suelos se realizó en el laboratorio de suelos de la Escuela de Agronomía de la UPTC, sede Tunja, utilizando para: textura, el método de Bouyoucos; pH, la relación 1:1 suelo:agua; acidez intercambiable, con KCL 1N; porcentaje de materia orgánica (MO), según Walkley-Black; conductividad eléctrica (CE), por extracto de saturación y conductivímetro; fósforo disponible, por el método de Bray II por colorimetría; Ca, Mg, K y Na, por extracción con NH4 Ac y absorción atómica; Fe, Mn, Cu y Zn, por extracción con DTPPA (siglas en inglés de ácido dietilen triamin pentametilenfosfónico) y absorción atómica; B, con agua caliente y S, con fosfato monocálcico (tabla 1).
Determinación de la actividad de la fosfatasa ácida
Se colocó 1 g de suelo en tubo, con 3 réplicas y 1 tubo control por muestra; a cada muestra se le adicionaron 1 mL de sustrato (solución de p-nitrofenil fosfato) y 4 mL de la solución tampón modificada a pH 6,5. Esta mezcla se incubó a 37 ºC por 1 h. Después de la incubación, a todos los tubos se les adicionaron 1 mL de CaCl2 0,5 M y 4 mL de NaOH 0,5 M. Se sirvió 1 mL de sustrato a los controles, posteriormente se filtró la suspensión y se midió la absorbancia a 400 nm. En forma simultánea, se prepararon blancos a los que se les añadió el sustrato después de la adición de CaCl2 0,5 M y NaOH 0,5 M.
Se preparó una solución de trabajo de 20 µg· mL-1 de p-nitrofenol fosfato y con ella se hizo una curva de calibración; se obtuvieron patrones de 0, 20, 40, 60, 80 y 100 µg de p-nitrofenol, de acuerdo con el método Tabatabai y Bremner (1969), modificado por Tabatabai y Eivazi (1977).
Recuento de grupos funcionales de microorganismos
Se cuantificaron los grupos funcionales de microorganismos relacionados con: el ciclo del P, las bacterias solubilizadoras de P (SP); el del C, las bacterias celulolíticas (CL), amilolíticas (AM) y proteolíticas (PR) y el del N, las bacterias libres fijadoras de N (FN) que utilizan la glucosa como fuente de C.
Para el recuento en placa se usó el medio de Silvestre- Bradley et al. (1982) modificado con fosfato tricálcico a pH de 6,5-7,0 para las bacterias SP; medio de Wood (1980) para las CL; medio de Pontecorvo et al. (1953) para las AM; medio de Wood (1980) para las PR y medio de Burk Wilson y Knight (1952) para las bacterias libres FN que utilizan la glucosa como fuente de C.
Las placas se incubaron a 28 ºC y las unidades formadoras de colonia (ufc) se contabilizaron luego de 3-5 d. Las ufc se expresaron por gramo de suelo seco. Las colonias consideradas positivas para actividad de solubilización de P, CL, AM, PR fueron aquéllas que presentaron un halo de degradación. En el caso de las bacterias libres FN se consideraron positivas todas las colonias que crecieron en el medio de cultivo.
Análisis estadístico
Con los datos de ufc determinados por recuento en placa de las muestras de rizosfera y suelo total, se realizó un análisis comparativo de las medias poblacionales para cada grupo funcional mediante prueba t-student para determinar si existen diferencias significativas en el tamaño de las poblaciones microbianas. Adicionalmente, se realizaron pruebas de correlación entre bacterias SP y los demás grupos funcionales para establecer interacciones.
En cuanto a la actividad de la fosfatasa ácida, se realizó una comparación de la actividad enzimática en la rizosfera y el suelo total en cada uno de los suelos estudiados y un análisis de correlación con los datos de ufc de las bacterias solubilizadoras de P. Todos los análisis se realizaron con la ayuda del programa estadístico Stat Graphics versión 4.0.
Resultados y discusión
Actividad de la fosfatasa ácida
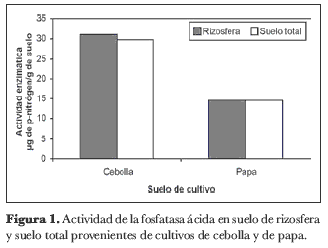
En las muestras de suelo correspondientes al cultivo de cebolla junca se observó una mayor actividad enzimática. Los valores de la enzima en estas muestras variaron entre 27,2 y 36,1 µg de p-nitrofenol por gramo de suelo para el total de las muestras, mientras que en las muestras del suelo con cultivo de papa se obtuvieron valores entre 8,7 y 20,6 µg de p-nitrofenol por gramo de suelo. En ninguno de los casos se presentó una diferencia significativa de la actividad de la fosfatasa ácida entre las muestras de rizosfera y de suelo total (figura 1).
Como los procesos de mineralización y solubilización determinan en gran medida la disponibilidad de P en el suelo, se aplicó una prueba de correlación lineal para establecer si la actividad de la fosfatasa ácida (Fa) está relacionada con la población de bacterias solubilizadoras de fósforo (SP), tanto en rizosfera como en suelo total. Los coeficientes de correlación encontrados entre SP y fosfatasa ácida (Fa) no fueron estadísticamente significativos y algunos de ellos mostraron valores demasiado bajos para indicar una relación estrecha entre la actividad de la fosfatasa y la población de solubilizadoras de P en el suelo (tabla 2).

La actividad de la fosfatasa ácida puede ser afectada por varios factores edáficos. Falih y Warinwright (1996), Martin (1984) y Weiss y Trespendorfer (1993), citados por Hysek y Sarapatka (1998), encontraron que la actividad de esta enzima es afectada por la adición de fuentes de C al suelo, la presencia de cobertura vegetal y la macrofauna del suelo.
Algunos de estos factores se evaluaron en este trabajo. Se determinó el porcentaje de materia orgánica en los suelos estudiados y se evaluó la actividad de la enzima en la rizosfera de cada cultivo. El suelo correspondiente al cultivo de cebolla presentó un mayor porcentaje de materia orgánica (5,54%) (tabla 1) y una mayor actividad de la enzima; este resultado también coincide con un mayor número de ufc por gramo de suelo en esta muestra, tanto para rizosfera como para suelo total (18,67· 103 ufc· g-1 de suelo). De acuerdo con esto, es probable que una mayor actividad de la fosfatasa ácida en el cultivo de cebolla esté asociada a un mayor contenido de materia orgánica y a una mayor cantidad de bacterias.
Existe otro factor que puede incrementar la actividad de las fosfatasas, según Cruz (1994) y Hadley (1994), citados por Portilla et al. (1998): la actividad de la fosfatasa ácida es más alta en la superficie de la raíz de la planta cuando existe deficiencia de P y, en el caso de los suelos estudiados, el suelo del cultivo de cebolla presentó una deficiencia de P disponible (4,61 ppm), coincidiendo también con una actividad alta de la enzima. En contraste, el suelo del cultivo de papa corrobora el efecto de los factores mencionados sobre la actividad de la enzima. En este suelo, como se mencionó arriba y como se puede observar en la tabla 1, se registró una actividad menor de la enzima, coincidiendo con un porcentaje menor de materia orgánica (2,22%), una cantidad menor de ufc en la totalidad de las muestras (10,92 · 103 ufc· g-1 de suelo) y un valor más alto de P disponible (48,7 ppm).
Al no encontrarse un coeficiente de correlación lo suficientemente considerable y estadísticamente significativo entre la actividad de la fosfatasa ácida y las bacterias solubilizadoras de P, se puede deducir que, aunque estos dos procesos determinen en gran medida la disponibilidad de fosfatos en el suelo, uno no está determinado por el otro, es decir, que se pueden considerar de forma independiente en este estudio.
Actividad microbiana
Microorganismos solubilizadores de P (SP)
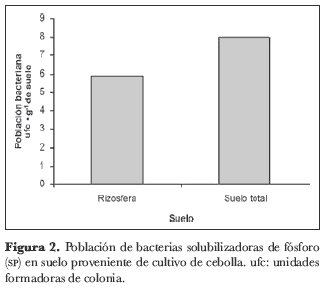
Para el suelo del cultivo de cebolla junca, se encontraron entre 0,5 y 22· 103 ufc· g-1 de suelo, en la totalidad de las muestras. En las muestras de rizosfera se hallaron entre 0,5 y 9,5· 103 ufc· g-1, mientras que en las muestras de suelo total los valores fluctuaron entre 4 y 22· 103 ufc· g-1 de suelo (figura 2).
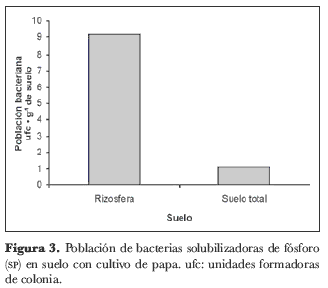
En cuanto al suelo del cultivo de papa, se registraron valores en la totalidad de la muestras entre 0,95 y 20,5· 103 ufc· g-1 de suelo. En las muestras de rizosfera se encontraron entre 3 y 20,5· 103 ufc· g-1 de suelo, mientras que en las muestras de suelo total se encontraron valores entre 0,95 y 1,47· 103ufc· g-1 de suelo (figura 3). Para los datos de este suelo se encontraron diferencias significativas en el número de ufc entre las muestras de rizosfera y de suelo total.
Los valores de ufc por gramo de suelo son superiores a los registrados por Adame (1996), citado por Linares (1999), quien indica que las bacterias solubilizadoras de fósforo (SP) existen efectivamente como parte de la microflora nativa de los suelos y sus niveles poblacionales fluctúan entre 1,7 y 1,58· 103 ufc· g-1 de suelo y de raíz.
El suelo como hábitat presenta muchas variables que pueden influenciar las poblaciones microbianas, de modo que éstas pueden presentar diferencias en sus niveles poblacionales entre un suelo y otro e incluso entre muestras de un mismo terreno. Sin embargo, estudios realizados en diferentes suelos por Carniero et al. (2004) y Barroti y Nahas (2000) indicaron que los suelos agrícolas pueden presentar valores de ufc más altos para SP que los suelos forestales y que uno de los factores que más influye sobre este parámetro es el pH del suelo modificado por la adición de fertilizantes en los suelos agrícolas; esta aplicación constante de químicos en los suelos agrícolas no sólo produce un aumento en el pH sino también un aumento en la disponibilidad de nutrientes, lo que puede favorecer el crecimiento poblacional de las SP y otros grupos bacterianos. Los valores en el pH de los suelos muestreados no presentan diferencias significativas: 6,2 para suelo de cultivo de cebolla y 6,6 para suelo de cultivo de papa; pero es importante resaltar que en ambos casos tiende a la neutralidad, siendo éste un pH óptimo para el crecimiento bacteriano y pueden presentarse en este caso valores más altos de ufc de los microorganismos SP.
La disponibilidad del P es controlada por la mineralización e inmovilización de la fracción orgánica y la solubilización y la precipitación de fosfatos en forma inorgánica (Silva, 1999). Por consiguiente, una mayor presencia de bacterias SP en estos suelos puede ser favorable para el desarrollo de las plantas, pues garantizaría una mayor disponibilidad de fosfatos asimilables.
Sin embargo, paradójicamente la disponibilidad de P en el cultivo de cebolla es muy baja, según los datos obtenidos mediante el análisis fisicoquímico del suelo (tabla 1). Al parecer, aunque no existe reducción en la población de SP, la disponibilidad de estos microorganismos no es tan buena como se esperaría; probablemente existen otros factores que pueden estar afectando la actividad de estas bacterias y, por lo tanto, la disponibilidad del P en esos suelos, tal como el grado de eficiencia en la capacidad solubilizadora de la cepas.
Microorganismos del ciclo del C (celulolíticos, proteolíticos, amilolíticos) y bacterias libres fijadoras de N
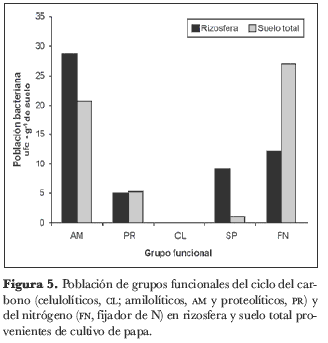
En la rizosfera del cultivo de cebolla junca, los grupos funcionales del ciclo del C presentaron un mayor número de ufc que las bacterias SP y las FN. La población de amilolíticos (AM) varió entre 20,5 y 83,5 · 103 ufc· g-1 de suelo, los proteolíticos (PR) entre 4,5 y 15,5· 103 ufc· g-1 de suelo y los celulolíticos (CL) entre 11,5 y 46· 103 ufc· g-1 de suelo (figura 4). En las muestras de suelo total, las bacterias de los grupos funcionales del ciclo del C siguieron presentando valores de ufc más altos que las FN y las SP. En las muestras de la rizosfera de papa, el grupo de bacterias con mayor número de ufc fueron las AM, seguidas de las poblaciones PR y de las bacterias FN (figura 5). Para las muestras de suelo, la abundancia bacteriana fue más alta para el grupo de AM, seguidas de las PR y las FN.
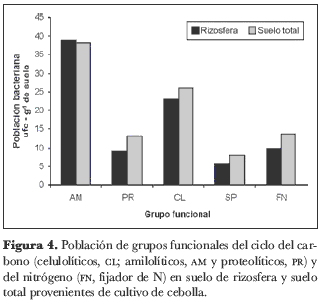
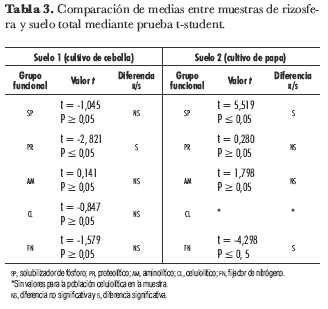
Con el fin de determinar la existencia de diferencias significativas en el número de ufc entre las muestras de rizosfera y suelo total para cada grupo funcional, incluyendo las bacterias SP, se realizó una prueba t-student para comparación de medias. Tal como se observa en la tabla 3, esta prueba determinó que sólo para las bacterias PR en muestra de cultivo de cebolla, las bacterias SP en cultivo de papa y las FN en cultivo de papa existen diferencias estadísticamente significativas entre muestras de rizosfera y suelo total.
Mediante análisis de varianza se determinaron diferencias estadísticamente significativas en el número de ufc por gramo de suelo entre cultivo de papa y cebolla. Para establecer en cuál de estos cultivos se presentaba un mayor número de ufc, se realizó una prueba de rango múltiple, que determinó que el suelo de cultivo de cebolla presenta un mayor número de ufc por gramo de suelo, con un promedio de 18.675· 103 ufc· g-1 de suelo.
El conteo de grupos funcionales del ciclo del C (AM, PR y CL), junto con bacterias FN de vida libre, permitió observar un mayor número de ufc de los grupos del C,alcanzando valores de hasta 83,5· 103 ufc· g-1 de suelo para el grupo de AM, que se constituye en el grupo con mayor numero de ufc en los dos suelos, tanto en las muestras de rizosfera como en las de suelo total (figuras 4 y 5).
Los valores de ufc para SP en relación con los demás grupos funcionales permiten observar que la población de SP es más reducida que los demás grupos de bacterias en las dos muestras de suelo y mantiene tamaños poblacionales casi similares a los del grupo de FN, tal vez porque este grupo presenta una correlación positiva en los dos suelos, tanto en rizosfera como en suelo total (tabla 4).
Con base en estos resultados se puede establecer cierto grado de dominancia de los grupos funcionales correspondientes al ciclo del C sobre los demás grupos bacterianos cuantificados. Esta predominancia puede estar quizás relacionada con la oferta de nutrientes proporcionadas por el porcentaje de materia orgánica disponible en el suelo.
La mayor abundancia poblacional de los grupos funcionales del ciclo del C en estos suelos puede influir en que la descomposición de los materiales orgánicos del suelo sea efectiva y que las moléculas complejas sean rápidamente degradadas, de forma tal que sus componentes estén disponibles para otros grupos bacterianos.
Al realizar una comparación de las poblaciones entre las muestras de rizosfera y de suelo total, se encontró que no todos los grupos funcionales presentaron diferencias significativas. Con un valor t = 2,074, 22 grados de libertad y un nivel de confianza de 95%, se encontró una diferencia significativa entre el número de ufc en rizosfera y suelo total para PR (t = -2,821, P = 0,05) en el suelo de cultivo de cebolla, así como para SP (t = 5,519, P = 0,05) y FN (t = 0,298, P = 0,05) en suelo de cultivo de papa (tabla 3). Las raíces de las plantas tienen una influencia directa en la composición y en la densidad de la microbiota del suelo; este efecto se conoce como efecto rizosférico y puede verse como la relación del número de organismos en el suelo de la rizosfera (R) sobre el número de microorganismos en el suelo alejado de las raíces (S). Generalmente la relación R/S está entre 5 y 20 y hasta 100, es decir, las poblaciones microbianas son hasta 100 veces mayores en la rizosfera (Gray y Parkinson; Woldendorp, 1978 citado en Atlas y Bartha, 2002). Sin embargo, es posible que estas proporciones puedan cambiar, pues en este trabajo las diferencias entre R y S para SP y los demás grupos funcionales resultaron en su mayoría ser poco significativas (tabla 3). El hecho de que los suelos cultivados estén sometidos a las labores de labranza podría determinar una variabilidad en sus propiedades físicas, químicas y en la disponibilidad de nutrientes, que ya no estarían limitados a la zona de la rizosfera de las plantas. Estas características de variabilidad podrían cambiar a su vez la proporcionalidad establecida en los microhábitat del suelo (Carniero et al., 2004).
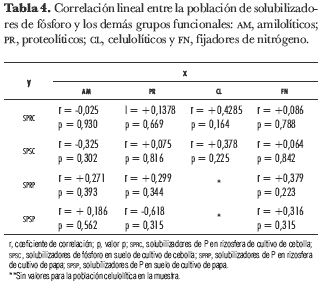
Los coeficientes de correlación encontrados entre SP y los demás grupos funcionales no resultaron estadísticamente significativos.
Para las muestras de cultivo de cebolla se encontraron los siguientes tipos de correlaciones: correlación negativa entre SP y AM en muestras de rizosfera y suelo total; correlación positiva entre SP y PR en muestras de rizosfera y suelo total; correlación positiva entre SP y CL en muestras de rizosfera y suelo total (tabla 4).
En muestras de cultivo de papa se encontró: correlación positiva entre SP y AM en muestras de rizosfera y suelo total; correlación positiva entre SP y PR en muestras de rizosfera y negativa en muestras de suelo total; con el grupo de CL no se estableció ninguna relación en este suelo por ausencia de datos para este grupo bacteriano (tabla 4).
En los dos suelos, tanto en las muestras de rizosfera como de suelo total, se halló una relación positiva entre SP y FN. Dentro de los valores del coeficiente de relación más altos estuvieron: los hallados entre SP y CL en la muestra de cultivo de cebolla, tanto en rizosfera como en suelo total (r = +0,4285, p = 1,16; r = +0,378, p = 0,22); entre SP y PR en las muestras de suelo total del cultivo de papa (r = -0,618, p = 0,31) y SP con FN en rizosfera (r = -0,8618, p = 0,78).
El crecimiento vegetal está directamente relacionado con las comunidades de grupos funcionales, porque diversas etapas de los ciclos de los nutrientes son mediadas exclusivamente por microorganismos, y algunos de éstos pueden participar en uno o más ciclos biogeoquímicos (Matsumoto et al., 2004). Según esto, el recuento de bacterias y las relaciones encontradas entre SP y los demás grupos funcionales en estos suelos podrían indicar el uso de bacterias SP como alternativa para suplir la deficiencia de P en los suelos agrícolas. Las relaciones negativas encontradas, aunque no son estadísticamente significativas, muestran una posible relación de competencia entre estos grupos bacterianos (tabla 4).
Algunas de las relaciones encontradas en el presente trabajo también fueron registradas en otros trabajos. Es el caso de la correlación positiva existente en SP y FN en la rizosfera de especies vegetales forestales y encontrada por Matsumoto et al. (2004), que ha sido relacionada directamente con el crecimiento de las plantas.
Las correlaciones encontradas entre SP y los grupos funcionales del ciclo del C son en su mayoría positivas, lo que puede estar relacionado con la función de degradación de la materia orgánica por parte de las bacterias PR, AM y CL, permitiendo así la disponibilidad de moléculas más sencillas para ser utilizadas por otros grupos bacterianos, como SP y FN. Sin embargo, cuando algún grupo de bacterias alcanza tamaños poblacionales muy por encima de los demás, pueden entrar en juego factores biológicos de carácter interespecífico, como competencia por sustratos, espacios y microhábitat, que pueden afectar el crecimiento de los demás grupos funcionales.
Conclusiones
Se encontró un mayor número de ufc en el suelo correspondiente al cultivo de cebolla, posiblemente asociado a un aumento en el pH y la disponibilidad de nutrientes del suelo por aplicación de fertilizantes. Las bacterias SP no se encontraron en menor proporción que los grupos funcionales del ciclo del C y mantuvieron tamaños poblacionales similares a las bacterias FN, tanto en rizósfera como en suelo total en los suelos de ambos cultivos. Según los resultados obtenidos, en estos suelos agrícolas la diferencia en la proporción de microorganismos entre rizosfera y el suelo circundante podría variar por la disponibilidad de nutrientes determinada por la adición de fertilizantes.
Se determinó que la actividad enzimática en estos dos suelos podría estar afectada por factores como el porcentaje de materia orgánica, la biomasa bacteriana y la disponibilidad de P.
En ninguno de los suelos muestreados se halló una relación significativa entre la actividad de la fosfatasa ácida y la población de bacterias SP.
Literatura citada
Andrade, G. 2004a. Role of functional groups of microorganisms on the rhizosphere microcosm dynamics. pp. 51-69. En: Varma, A., L. Abbott, D. Werner y R. Hampp. Plant surface microbiology. Springer-Verlag, Berlin. [ Links ]
Andrade, G. 2004b. The functional groups of microorganism used as bio-indicator on soil disturbance caused by biotech products such as Bacillus thuringiensis and Bt transgenic plants. pp. 121- 132. En: Varma, A., L. Abbott, D. Werner y R. Hampp (eds.). Plant surface microbiology. Springer-Verlag, Berlin. [ Links ]
Atlas, R. y R. Bartha. 2002. Ecología microbiana y microbiología ambiental. 4a edición. Pearson Educación S.A., Madrid. 500 p. [ Links ]
Bandinck, A.K. y R.P. Dick. 1999. Field management effects on soil enzyme activities. Soil Biol. Biochem. 31(11), 1471-1479. [ Links ]
Barroti, G. y E. Nahas. 2000. Populaçao microbiana total e solubilizadora de fosfato em solo submetido a diferentes sistemas de cultivo. Pesquisa Agropecuaria Brasileira 35(10), 1830-1838. [ Links ]
Carniero, R., L. Mendes, P. Lovato, A. Carvalho y L. Vivaldi. 2004. Indicadores biológicos asociados ao ciclo de fósforo em solos de cerrado sob plantio direto e plantio convencional. Pesquisa Agropecuaria Brasilera 39(7), 661-669. [ Links ]
Contreras, F., C. Rivero y J. Paolini. 1996. Efecto del uso de residuos orgánicos y dos tipos de labranza sobre la actividad de la fosfatasa ácida en un Alfisol. Revista Facultad de Agronomía (Maracay) 22,139-149. [ Links ]
Créptin, J. y R. Johnson. 2000. Soil sampling for environmental assessment. Soil sampling and methods of analysis. Canadian Society of Soil Science. [ Links ]
Feijo, A. y H. Quintero. 2001. Relación del uso de la tierra en agro ecosistemas del neotrópico con la diversidad del suelo. X Congreso de la Sociedad Colombiana de la Ciencia del Suelo, Medellín (Colombia). [ Links ]
Hysek, J. y B. Sarapatka. 1998. Ralationship between phosphatase active bacteria and phosphatase activities in forest soils. Biol. Fertil. Soil 26,112-115. [ Links ]
Linares, N. 1999. Efecto de las bacterias solubilizadoras de fósforo en la producción del cultivo de arroz. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 70 p. [ Links ]
Madigan, M., J. Martinko y J. Parker. 2004. Brock biología de los microorganismos. 4a edición. Prentice Hall Iberia, Madrid. 600 p. [ Links ]
Matsumoto, L., A. Martines, M. Avanzi, U. Albino, C. Brasil, D. Saridakis, L. Rampazo, W. Zangaro y G. Andrade. 2004. Interactions among functional groups in the cycles of carbon, nitrogen and phophorous in the rizosfhere of three succesional species of tropical woody trees. J. Appl. Soil Ecol. 10 (1016),1-9. [ Links ]
Oberson, M., J.M. Besson, N. Maire y H. Sticher. 1996. Microbiological processes in soil organic phosphorus transformations in conventional and biological cropping systems. Biol. Fertil. Soil 21, 138-148. [ Links ]
Osorio, N.W. y J.C. Pérez. 2001. Microbial solubilization of phosphates in soil. A review. X Congreso de la Sociedad Colombiana de la Ciencia del Suelo, Medellín (Colombia). [ Links ]
Pontecorvo, G., J.A. Roper, L.M. Hemons, K.D. MacDonalds y A.W.J. Buffon. 1953. The genetics of Aspergillus nidulans. Adv. Gen. 5, 141-238. [ Links ]
Porras, C. 1994. Metabolismo de los nutrientes en las plantas. pp. 35- 42. En: Porras, C. y R. Silva-Lora. Fertilidad de suelos. Diagnóstico y control. 2a edición. Editorial Guadalupe Ltda., Bogotá. [ Links ]
Portilla, C., E. Molina, G. Cruz-Flores, I. Ortiz y G. Manske. 1998. Mycorrhizal arbuscular colonization, phosphatase activity and root length as response to the phosphorus stress in wheat and triticale cultivated in an andisol. Terra 16(1), 55-61. [ Links ]
Silva-Lora, R. 1994. Factores que afectan la disponibilidad de nutrientes para las plantas. p 20-28. En: Porras, C. y R. Silva- Lora. Fertilidad de suelos. Diagnóstico y control. 2a edición. Editorial Guadalupe Ltda., Bogotá. [ Links ]
Silva, J. 1999. The soil habitat. Principles and applications of soil microbiology. Vol 2. Editorial Prince Hall, New Jersey. 400 p. [ Links ]
Sylvester-Bradley, R., N. Akasawa, S. La Torranca, F. Magalhães, L.A Oliveira y R.M. Pereira. 1982. Levantamento quantitativo de microrganismos solubilizadores de fosfatos na rizosfera de gramíneas e leguminosas forrageiras na Amazonia. Acta Amazonica 12, 15-22 [ Links ]
Tabatabai, M.A. y M.J. Bremner. 1969. Use of p-nitrophenylphosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1, 301-307. [ Links ]
Tabatabai, M.A. y R. Eivazi. 1977. Phosphatases in soils. Soil Biol. Biochem. 9,167-172. [ Links ]
Vera, D., H. Prez y H. Valencia. 2002. Distribución de hongos solubilizadores de fósforo de dos unidades fisiográficas de Guaviare, Colombia. Rev. Acta Biológica Colombiana 7(1), 23-31. [ Links ]
Vera, D., H. Pérez y H. Valencia. 2002. Aislamiento de hongos solubilizadores de fosfatos de la rizosfera de arazá (Eugenia stipitata, Myrtaceae). Rev. Acta Biológica Colombiana 7(1), 33-40. [ Links ]
Wood, P. 1980. Specify in the interactions of direct dyes with polysaccharides. Carbohydrate Res. 85, 271-287 [ Links ]
Wilson, P. y S.G. Knight. 1952. Experiments in bacterial physiology. Burguess, USA. [ Links ]
Wright, A. y K. Reddy. 2000. Phosphorous loading on extracellular enzyme activity in everglades wetland soil. Appl. Environ. Microbiol. 67(4),1830-1838. [ Links ]

