










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.1 Bogotá jan./jun. 2007
FISIOLOGÍA DE CULTIVOS
Martha Lucía González1, Claudia Caycedo1, María Fernanda Velásquez1, Víctor Flórez2 y Miriam Ruby Garzón3
1 Licenciada en Biología, Universidad Distrital Francisco José de Caldas, Bogotá. e-mail: marlejcita@yahoo.com
2 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. e-mail: vjflorezr@unal.edu.co
3 Profesora asociada, Proyecto Curricular de Biología, Universidad Distrital Francisco José de Caldas, Bogotá.
Fecha de recepción: 28 de febrero de 2003. Aceptado para publicación: 06 de junio de 2007
Resumen: La coliflor es una de las hortalizas de mayor consumo en Colombia; es muy apetecida comercialmente por sus aportes nutricionales y por ello su mercado se ha ampliado a congelados y procesados con fines de exportación. Con el propósito de determinar el efecto del ácido giberélico (AG3) sobre el crecimiento de órganos cosechados se realizó un experimento en dos épocas de siembra en los que se aplicaron concentraciones de 0, 5, 25 y 125 mg· L-1 de AG3. Se cuantificaron variables como la masa seca total, la masa seca por órganos (raíz, tallo e inflorescencia), el número de flores y la altura de planta. Los resultados demostraron que la dosis de 25 mg· L-1 de AG3 fue la más apropiada para inducir la floración y obtener mayor altura de planta, mientras que la dosis de 5 mg· L-1 de AG3 permitió acumular mayor cantidad de biomasa.
Palabras clave: desarrollo, biomasa, fotoasimilados, fuente-demanda, planta en forma de roseta.
Abstract: The cauliflower is one of the horticultural crops of most consumption in Colombia. It is commercially attractive because of its nutritional qualities, the reason explaining why the market of this crop has been expanded from a processed to an exportation one. In order to verify the effect of gibberellic acid (AG3) on growth of cauliflower organs with a commercial value, an assay containing AG3 with 0, 5, 25 and 125 mg· L-1 concentrations was performed in two periods of planting. The total dry mass, organ (root, stalk, and inflorescence) dry mass, number of flowers, and plant height were quantified. The results showed that AG3 dose of 25 mg· L-1 was the most appropriate one for floral induction and largest plant height, while the dose of 5 mg· L-1 allowed to accumulate the highest quantity of biomass.
Key words: development, biomass, photoassimilates, sink and source, rosette plants.
Introducción
LA COLIFLOR ES UNA PLANTA anual o bienal originaria de Europa y Asia Occidental que presenta formas muy variadas y, por cruzarse fácilmente, se hace difícil la obtención de semillas puras (Lobo et al., 1983). Para algunos botánicos, coliflores y brócolis pertenecen a las mismas especie y variedad (Brassica oleraceae L. var. Botrytis DC), distinguiéndose únicamente en su forma, pues las coliflores presentan forma cauliflora y los brócolis tienen forma cimosa (Maroto, 1992). No obstante, Pérez (1978) diferencia el brócoli como Brassica oleraceae L. var. Asparagoides DC, cuya variedad hace referencia a la forma de espárrago que presenta, mientras que en la coliflor, la variedad describe una forma arracimada.
Según Krarup y Moreira (1998) el sistema radical de las Brassica es reducido, con una raíz pivotante de cerca de 50 cm de largo y raíces laterales relativamente pequeñas, provistas de numerosos pelos radicales. Las hojas son sésiles, enteras, poco a muy onduladas, oblongas (de unos 40 a 50 cm de largo y 20 cm de ancho), elípticas y muy erguidas, extendiéndose de forma más vertical y cerrada en la coliflor que en el brócoli. Durante el desarrollo de la planta se puede diferenciar claramente la fase juvenil en la cual la planta sólo forma hojas y raíces; la duración de este período es variable, según los requerimientos de horas/frío para iniciar la floración y la ocurrencia de esas horas en el ambiente donde se desarrolla la planta.
Por acción de temperaturas vernalizantes (entre 6 y 10° C) ocurren los cambios fisiológicos necesarios para la formación de la pella o pan, que corresponde al órgano de consumo de esta variedad. En los Trópicos la planta casi siempre permanece vegetativa a causa de las altas temperaturas, a menos que se usen cultivares que se hayan desarrollado específicamente para esas condiciones. En las zonas temperadas las formas anuales pasan a la fase reproductiva después de formado cierto número de hojas. Durante la transición a la floración, la mayor parte de las sustancias de reserva elaboradas por las hojas son movilizadas hacia el meristema apical del tallo principal, donde ocurren sucesivas divisiones del ápice para formar los tallos preflorales (futuros pedúnculos) que sostienen los nuevos y múltiples meristemas apicales. En conjunto conforman el pan o pella. Al progresar la fase de inducción de la pella, la planta cesa la formación de hojas; las más jóvenes envuelven progresivamente el pan protegiéndolo de las condiciones externas y de la luz. Posteriormente, el pan se hace visible al desarrollar un diámetro creciente.
En la floración de los tipos de Brassica no vernalizantes las ramificaciones preflorales del pan inician el crecimiento longitudinal pasando a constituirse en los pedúnculos de la inflorescencia; el pan se desarma y comienza a amarillear, y un número significativo de ápices se diferencian en reproductivos, para desarrollar posteriormente las flores de color amarillo. Finalmente, se desarrolla un fruto que corresponde a una silicua amarillenta, de 7 a 8 cm de largo, con cerca de 20 semillas pequeñas y redondas, de color rojizo a pardo oscuro (300 semillas/g) (Krarup y Moreira, 1998). Son semillas sin endospermo y el cotiledón presenta un alto contenido de aceite. La capacidad germinativa media es de aproximadamente cuatro años desde el momento de su recolección (Maroto, 1992).
Según los requerimientos térmicos para la formación de la pella se reconoce la existencia de tres clases de coliflores (Krarup y Moreira, 1998):
Coliflores tropicales. Aquellas capaces de producir pellas de ápices vegetativos, de calidad aceptable, en condiciones de temperatura superior a 20°C. Son selecciones hechas con períodos prolongados de altas temperaturas, como por ejemplo, el cultivar ‘Tropical 55' y sus híbridos.
Coliflores vernalizantes. Aquellas que requieren un período de bajas temperaturas para producir pellas constituidas por flores propiamente dichas (ápices reproductivos). Estas son selecciones típicas de los países del norte de Europa, como por ejemplo el cv. ‘Walcheren Winter' y sus derivados, así como el cv. ‘Gigante de Nápoles'.
Coliflores no vernalizantes. Aquellas capaces de producir pellas constituidas por ápices vegetativos, de alta calidad, con temperaturas de 14 a 20°C propias de zonas o épocas temperadas. En esta clase se encuentra el mayor número de cultivares de polinización abierta y de híbridos obtenidos en la variedad. Los cultivares más tradicionales de este tipo son ‘Erfurt', ‘Snowball' y sus selecciones, ‘Suprimax' y ‘Matra'. En los últimos años se ha ido incrementado el uso de híbridos como ‘Arfak', ‘Guardian', ‘Incline' y ‘Serrano'. En las condiciones ecofisiológicas locales el principal inconveniente es el inicio precoz de la inflorescencia, lo que impide la formación de una pella o cabeza compacta y con buen desarrollo. El clima propicio para el desarrollo normal de la mayoría de las variedades de coliflor es una temperatura media, sin heladas y humedad relativa alta; condiciones que inciden en las épocas de cosecha. Así, una temperatura alta durante la formación de la pella hace que crezca rápidamente, sin obtenerse un buen tamaño o con la aparición de defectos, en consecuencia de una rápida división celular, como el surgimiento de pequeños botones y manchas. También se ha observado una estrecha relación entre el desarrollo de las hojas y el de la pella cuando la temperatura es superior a 35 ºC o inferior a 15,5 ºC; así mismo, se ha observado que en las noches frías se reduce el tamaño de las hojas (Murayama, 1987).
Fernández (1995) encontró que la fase juvenil de la coliflor finaliza, para los cultivares ‘Siria' y ‘Nautilus', con la formación de 19-20 hojas, y que variaciones en la temperatura y la iluminación durante el cultivo pueden afectar su duración cronológica al incidir sobre el número de hojas formado.
En zonas cálidas se han adaptado variedades con tolerancia a altas temperaturas, como es el caso de ‘Snowball', ‘Farmers' y otros tipos de coliflor en el Valle del Cauca (Jaramillo, 1982). La coliflor se siembra en suelos sueltos con pH entre 5,5 y 6,8. Si el pH es más bajo se presentan problemas por ser la especie poco tolerante a la acidez y, por lo tanto, al exceso o deficiencia de molibdeno. En consecuencia, las hojas reducen su lámina foliar, se tornan comprimidas, deformes, encrespadas y con la nervadura principal saliente, síntomas conocidos como ‘cola de látigo'. Las condiciones ambientales desfavorables como vientos continuos, heladas y períodos de sequía, así como una deficiente preparación del suelo, contribuyen a intensificar los síntomas de deficiencia de Mb (Murayama, 1987). Además, los suelos dedicados a este cultivo deben presentar buen drenaje porque el crecimiento de la hortaliza precisa ser rápido y regular (Carvajal, 1983).
Las aplicaciones de giberelinas en las plantas Brassica aceleran la iniciación de la pella y reducen el número de hojas formado tan sólo en condiciones débilmente inductivas (22°C), mientras que no se manifiestan en condiciones fuertemente inductivas (10°C), por lo que las aplicaciones de giberelinas pueden ser empleadas para adelantar la iniciación de la pella en condiciones subóptimas de vernalización (Fernández, 1995).
Han sido publicadas diversas experiencias de aplicación de ácido giberélico (AG3) con el fin de obtener una producción precoz, reducir el número de nudos y alargar la longitud de los mismos (Jaramillo, 1982). Las giberelinas (AG) fueron descubiertas en el Japón, en la década de los 30, en plantas de arroz enfermas a causa del hongo Gibberella fujikuroi. Son compuestos isoprenoides, específicamente diterpenos cíclicos, que se sintetizan a partir de unidades de acetato del acetil CoA en la ruta del ácido mevalónico. Todas las AG se derivan del esqueleto entgiberelano; presentan 19 ó 20 átomos de C agrupados en sistemas de 4 ó 5 anillos y un grupo carboxilo unido al C7, esencial para su actividad biológica. En general, las AG de 19 carbonos son más activas que las de 20 (Taiz y Zeiger, 1998; Azcón-Bieto y Talón, 1993). La mayor parte de las 136 giberelinas) caracterizadas hasta la fecha son precursores o productos inactivos de las AG que poseen actividad biológica intrínseca. Pese al gran número de AG que ocurren naturalmente las que presentan actividad biológica son muy pocas; entre ellas AG1, AG3, AG4, AG5, AG6 y AG7. De las numerosas AG, 128 fueron identificadas en diferentes especies de plantas vasculares y también en siete bacterias y siete hongos (Srivastava, 2002; Sponsel y Hedden, 2004).
Hooley (1994), citado por Fu et al. (2001), describe las AG como hormonas diterpenoides tetracíclicas esenciales para el normal desarrollo de las plantas. Los niveles de AG en los vegetales están regulados por mecanismos homeostáticos que incluyen cambios en la expresión de una familia de enzimas de inactivación de AG, conocidas como AG-2-oxidasas (Singh et al., 2002). Las AG cumplen un importante papel fisiológico en el desarrollo de las semillas, el desarrollo de la floración, el crecimiento del tubo polínico y la elongación de brotes y tallos. Los cambios en la concentración de la hormona y la susceptibilidad del tejido vegetal influyen en estos procesos. Sin embargo, los mecanismos moleculares por los cuales las AG son traducidas a cambios morfológicos y bioquímicos dentro de las plantas son desconocidos (Ikeda et al., 2001).
De acuerdo con Sponsel (1996), las giberelinas son sintetizadas a partir del ácido mevalónico en tejidos jóvenes, aunque el sitio exacto aún no se conoce. Todos los tejidos de crecimiento son potencialmente sitios de biosíntesis. Existen evidencias en las cuales el desarrollo de frutos y semillas ocurre en sitios de biosíntesis por el alto contenido de enzimas que pueden convertir el ácido mevalónico (precursor) a AG de 19 carbonos. Autores como O'Neill y Ross (2002) han demostrado que el ácido 3-indol acético (AIA) promueve la biosíntesis de la giberelina activa (AG1) en brotes de arveja (Pisum sativum) en virtud de la supresión de un gen de esta planta llamado SLN transcribe niveles de AIA y dicho gen está involucrado en la activación de AG1 en los brotes extraídos de los cultivos de tejidos.
Las giberelinas son una clase de hormona vegetal con efectos de diversa índole, uno de ellos es la represión sobre los genes del enanismo, al producir un crecimiento normal de plantas genéticamente enanas e incluso de especies cuyo desarrollo natural del tallo hace que no pasen del estadio de roseta, como la col (Luckwill, 1994).
La eliminación continua de las raíces provoca la disminución de las concentraciones de giberelinas en las partes aéreas de las plantas, de lo cual se puede deducir que las raíces aportan parte de las giberelinas que sustentan el sistema aéreo por medio del xilema, o que la falta de agua y nutrientes afecta su producción en los demás órganos (Salisbury y Ross, 2000).
En los tejidos vegetativos, las giberelinas nativas son estructuralmente menos diversas que en los tejidos reproductivos. La AG12, giberelina precursora de las demás, tanto en plantas como en G. fujikuroi, puede ser hidroxilada en el carbono tres, en el carbono trece o en ambos. En casi todas las especies estudiadas (maíz, arveja, arroz, espinaca), la ruta más importante, o la única en los tejidos de la parte aérea, es la vía de la 13 hidroxilación temprana, en donde la AG20 es la primera giberelina de 19 carbonos formada y ésta se transforma en AG1 a través de 3Β-hidroxilación. La AG1 sería la giberelina activa en la elongación de la parte aérea en las especies mencionadas. Las giberelinas AG3, AG4 y AG7 también son 3 Β-hidroxiladas como la AG1 y, por tanto, potencialmente son tan bioactivas como la AG1 (Taiz y Zeiger, 1998; Azcón-Bieto y Talón, 1993). Más que cualquier otra giberelina, la AG1 ha sido identificada en 86 especies de plantas, indicando que desempeña un papel amplio e importante como la ‘verdadera' giberelina (Sponsel y Hedden, 2004). Existen evidencias de que las giberelinas se translocan en las plantas a través del floema y del xilema (Gómez, 1994). Entre los efectos de las giberelinas, se conoce ampliamente el crecimiento de los tallos, el cual involucra una secuencia de procesos y respuestas, como son la recepción de señales, la activación de uno o más señales de transducción para la transcripción de la respuesta primaria por parte de los genes y una respuesta secundaria que se traduce como tal en la elongación celular. Este efecto se evidencia en el incremento de la longitud en las células y el número de las mismas, lo cual es directamente proporcional al número de aplicaciones de AG3 (Taiz y Zeiger, 1998).
Después de la inducción, las giberelinas promueven diversos aspectos del desarrollo floral, como la identidad de los órganos del meristemo floral, el crecimiento de anteras, y el desarrollo y pigmentación de la corola. En las plantas en roseta, las condiciones de día largo o la aplicación de giberelinas incrementa el tamaño de la región meristemática subapical al aumentar la proporción de células que entran en división celular; esta nueva región meristemática produce la mayoría de células que contribuyen posteriormente a la elongación del tallo (Azcón-Bieto y Talón, 2000).
Según Pantastico (1984), el movimiento de los nutrientes desde las fuentes hasta los órganos almacenadores (flores, frutos) se efectúa en contra de un gradiente de concentración. La coliflor es una planta en roseta y, por tanto, es más fácil visualizar el efecto del AG3 sobre la altura de la planta.
El objetivo de este trabajo fue determinar el efecto de la aplicación del ácido giberélico (AG3) sobre el crecimiento de diferentes órganos de la planta de la coliflor (Brassica oleraceae L. ‘Botrytis DC').
Materiales y métodos
El experimento se realizó en la sede del Centro Multisectorial del Servicio Nacional de Aprendizaje (SENA) ubicado en el municipio de Mosquera (Cundinamarca), el cual se halla ubicado a 2.550 msnm con latitud norte de 4º 42' 28” y una longitud oeste de 74º 13' 58”; la temperatura promedio es de 13,1ºC y presenta una precipitación media anual de 646 mm (Instituto Geográfico Agustín Codazzi - IGAC, 1996). Esta zona, según Espinal y Montenegro (1963), hace parte de la formación bosque seco montano bajo.
Se utilizó semilla certificada de coliflor (Brassica oleraceae L. ‘Botrytis DC') y ácido giberélico (AG3) al 90% de laboratorios Merck (Auxilin°). Los tratamientos ensayados correspondían a las concentraciones de 0, 5, 25 y 125 mg· L-1 de AG3, con tres repeticiones cada uno y dos etapas de siembra. Para tal fin fue utilizada una parcela de 10 m x 15 m a la que se transplantaron las plántulas de ambas etapas; éstas fueron sembradas a 50 cm de distancia en el surco y a 1 m entre surcos. En la primera etapa el transplante se realizó 30 días después de la siembra (dds), con plántulas entre 5 y 7 cm de altura; la segunda etapa fue transplantada 60 dds, con plántulas entre 10 y 12 cm de altura. En total fueron ocho aplicaciones de solución acuosa de AG3, a intervalos de 15 días y empezando 15 días después del transplante.
En campo, fue contado el número de flores y medida la altura de las plantas. En el laboratorio, para la cuantificación de la masa seca, las muestras fueron sometidas a 105oC durante 24 horas.
Se utilizó un Diseño Completo al Azar (DCA) con arreglo factorial de 4x3x2 y los datos se procesaron con el sistema de análisis estadístico SAS°, versión 6,07 de 1992. También fueron sometidos a la prueba Tukey para establecer comparación entre las medias de los tratamientos.
Resultados y discusión
Masa seca por órganos
Pese a que no se verificaron diferencias estadísticamente significativas en la etapa 1 para la variable ‘masa seca de la raíz', el mayor promedio se logró con la concentración de 125 mg· L-1 de AG3 (figura 1). En la etapa 2 los tratamientos testigo y 5 mg· L-1 de AG3 difieren significativamente (figura 1). Sin embargo, los tratamientos con 25 y 125 mg· L-1 de AG3 no difieren estadísticamente del control ni del tratamiento de 5 mg· L-1 de AG3.
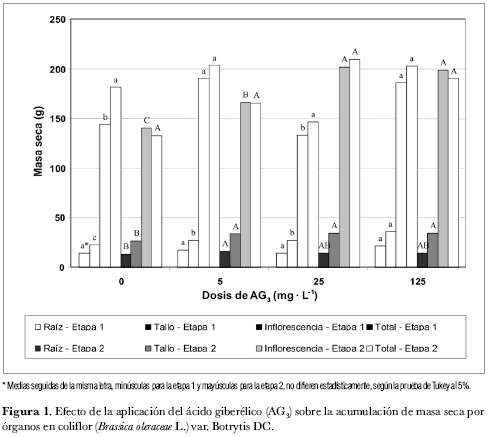
En cuanto a la acumulación de masa seca en el tallo para las dos etapas de siembra, se observa una tendencia creciente en la medida en que se incrementa la concentración de AG3 (figura 1). Así mismo, es comprobable que, a excepción del tratamiento con 125 mg· L-1 de AG3, los valores de la etapa 2 son mayores a los de la etapa 1.
Para ambas etapas, hubo diferencia estadísticamente significativa entre los tratamientos (figura 1), considerándose las dosis de 125 mg· L-1 y de 5 mg· L-1 de AG3 como las de respuesta óptima en las etapas 1 y 2, respectivamente.
Soriano (1995) afirma que cada planta se puede considerar fisiológicamente como un conjunto de órganos productores y consumidores de fotoasimilados, interrelacionados por haces conductores que facilitan el movimiento de estas sustancias desde los primeros hacia los segundos. Para Salisbury y Ross (2000), los productos de la asimilación metabólica de las hojas, incluso los fotosintatos, son necesarios para el desarrollo de todos los órganos de la planta. Los órganos capaces de fotosintetizar pueden suplir estas necesidades en estadios avanzados de desarrollo cuando son fotosintéticamente activos.
Respecto del comportamiento de la acumulación de fotoasimilados en la inflorescencia, en la etapa 2 se verifica una tendencia creciente en la acumulación de masa seca, en la medida que aumenta la concentración del AG3 (figura 1). Para la etapa 1 es verificable la diferen cia significativa de los tratamientos de AG3 con respecto al testigo, excepto para la concentración de 25 mg· L-1. Considerando el efecto de la fitohormona sobre la acumulación de carbohidratos en la inflorescencia de la coliflor, los resultados con más economía de AG3 se obtienen con la dosis de 5 mg· L-1 para las plantas de la etapa 1 y con 25 mg· L-1 para las plantas de la etapa 2.
Fernández (1995) empleó plántulas procedentes de cepellón de mayor tamaño y consiguió producciones similares, una duración de la recolección similar y una reducción del ciclo de cultivo, por lo que su uso se puede aconsejar en algunos ciclos de cultivo, si bien el costo de producción es algo superior. El uso de plántulas de mayor edad no resultó aconsejable ya que se produjeron pellas de menor masa y se alargó el periodo de recolección, aunque se disminuyó considerablemente la duración del ciclo de cultivo.
La respuesta de las plantas a mayores concentraciones de AG3 durante la segunda etapa, podría atribuirse a las condiciones ambientales en esa época (enero a abril de 1997), cuando se presentó el Fenómeno del Niño. Ante esta situación la planta translocaría sus asimilados en función de producir flores y profundizar el sistema radical, como posibilidad de sobrevivencia a condiciones adversas. Esto explicaría que el tratamiento con 5 mg· L-1 AG3 para la variable ‘masa seca de la raíz' en la etapa 2 presentó diferencias significativas con relación al tratamiento testigo (figura 1).
La absorción foliar de AG3 es influenciada, no sólo por las condiciones climáticas durante y después de la pulverización, sino también por las condiciones en que se ha desarrollado la hoja, ya que pueden haber afectado el grosor y estructura de la cutícula (Luckwill, 1994).
La radiación solar incrementa la temperatura y como consecuencia se desarrollan manchas en la superficie los órganos a causa de quemaduras solares; también ocurre deshidratación acelerada de las hortalizas de hoja, así como de las hortalizas de inflorescencias como la coliflor y el brócoli (Cañet et al., 2003).
Aunque todavía no hay una síntesis clara sobre el papel de las fitohormonas en la respuesta de la planta ante el estrés, debido a que son muchas las funciones implicadas y muy complejas las interacciones, es posible que los factores de estrés perturben el equilibrio hormonal y, por tanto, produzcan respuestas especificas en el desarrollo de las plantas (p.e. la floración bajo déficit hídrico) y en los ajustes osmóticos (Itai, 1999, citado por Reigosa et al., 2004).
A pesar de que no se verificaron diferencias estadísticamente significativas entre los tratamientos para ambas etapas, en cuanto a la acumulación de masa seca total (figura 1) se puede observar un aumento progresivo durante la etapa 2 para las concentraciones de 5 y 25 mg· L-1 de AG3.
La relación fuente/demanda que se presenta en los órganos de las plantas durante su desarrollo está afectada por una serie de procesos inherentes a cada especie y por la influencia climática. Según Mason y Maskell, citados por Barceló et al. (1995), un órgano de demanda es un lugar de consumo de sustancias orgánicas, bien sea para la diferenciación de tejidos, para la acumulación de sustancias de reserva o para el mantenimiento de un órgano o tejido. Este es el caso del desarrollo de frutos, meristemos, hojas jóvenes, cotiledones, endospermos de semillas en formación, tejidos de tallo, hoja o raíz, cuando almacenan estas sustancias, principalmente carbohidratos. Para Taiz y Zeiger (1998) el proceso de elongación celular promovido por las giberelinas sería independiente de la orientación de las microfibrillas de celulosa en la pared celular y no actuarían incrementando la tasa de absorción de agua. Se ha planteado la hipótesis de que las giberelinas actuarían de forma asociada con las auxinas modificando las propiedades de la pared celular, donde las auxinas inducen la extrusión de protones y las giberelinas estimulan la actividad de la enzima endotransglicosilasa xyloglucano (XET), la cual permite la penetración de la expansina en la pared celular, en donde estas hormonas se activarían por el pH ácido.
Altura de planta
De acuerdo con la figura 2, durante la etapa 1 se presentaron los valores más altos para la variable ‘altura de planta'. Aunque la concentración de 25 mg· L-1 de AG3 mostró los mayores promedios en ambas etapas, no hay diferencias significativas estadísticamente entre los tratamientos.
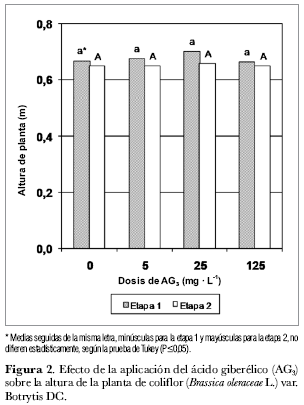
La coliflor es una planta en forma de roseta y, por tanto, es más fácil visualizar el efecto del AG3 sobre la altura de la planta. Entre tanto, la altura alcanzada por las plantas tratadas con AG3 no es común en plantas de esta especie. El crecimiento del tallo, en las plantas en condiciones de día largo y con forma de roseta, depende del fotoperíodo. En condiciones de día corto, estas especies permanecen en estadio vegetativo y desarrollan una roseta de hojas basales. La elongación del tallo está mediada por el control fotoperiódico del metabolismo de giberelinas (Azcón-Bieto y Talón, 1993 y 2000). Además, las giberelinas inducen elongación del tallo y la floración en variedades genéticamente enanas (Phinney, 1956, citado por Azcón-Bieto y Talón, 1993). Las coles y otras especies en forma de roseta que tienen entrenudos cortos en ocasiones crecen hasta 2 m de altura y después florecen, tras la aplicación de AG3, mientras que las plantas no tratadas permanecen pequeñas y en estadio vegetativo (Salisbury y Ross, 2000).
Las AGs actúan sobre el RNA desinhibiendo genes. Esta acción está caracterizada con respecto a dos genes, que en ausencia de AGs se hallan reprimidos. El metabolismo de las plantas está regulado, entre otros compuestos, por la presencia de hormonas endógenas, que actúan en determinados momentos, ante condiciones adversas, como el ataque de plagas y enfermedades, y variaciones climáticas desfavorables (Gómez, 1994).
Número de flores
En la figura 3 se constata un efecto significativamente mayor del tratamiento con 25 mg· L-1 de AG3 sobre el número de flores en la etapa 1. Durante la segunda etapa no hay diferencias significativas entre los tratamientos. Las giberelinas modifican sustancialmente los procesos reproductivos de los vegetales, participando en el control de la inducción de la floración, en el crecimiento y producción de flores, y en el cuajado y desarrollo de los frutos (Azcón-Bieto y Talón, 2000). La aplicación de giberelinas a los tallos produce un incremento pronunciado de la división celular en el meristemo subapical (Sach et al., 1960 citado por Weaver, 1996). Pueden provocar la floración en muchas especies que requieren temperaturas frías, como son la zanahoria, escarola, col y nabo (Weaver, 1996). Puede sustituir la exigencia de días largos y suplir la necesidad de un periodo inductivo frío de algunas especies si están a punto de florecer o para hacerlo más pronto (Salisbury y Ross, 2000).
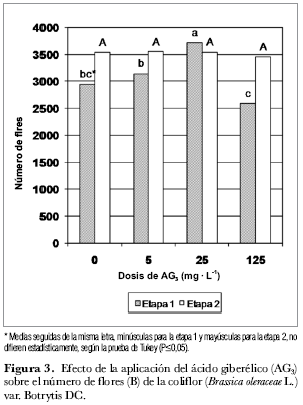
Mediante el presente estudio fue constatado el efecto de la aplicación del ácido giberélico (AG3) sobre la acumulación de masa seca en los diferentes órganos de la planta de la coliflor (Brassica oleraceae var. ‘Botrytis DC'). Se consideran como resultados relevantes los tratamientos de 125 mg· L-1 en la etapa 1 y de 5 mg· L-1 de AG3 en la etapa 2, para la variable ‘acumulación de masa seca en el tallo'; así mismo, la dosis de 25 mg· L-1 de AG3 para las variables ‘número de flores' durante la etapa 1 y ‘acumulación de fotoasimilados en la inflorescencia' durante la etapa 2. De otra parte, para ambas etapas de siembra, el efecto de la fitohormona sobre las variables masa seca total y altura de planta no fue significativo entre tratamientos. Por último, se enfatiza en que las plantas no deben ser sometidas a estrés climático pues se interferiría con el efecto de la fitohormona.
Agradecimientos
Los autores agradecen en forma especial a la Facultad de Agronomía de la Universidad Nacional de Colombia y a los laboratorios del Proyecto Curricular de Licenciatura en Biología de la Universidad Distrital Francisco José de Caldas por la disposición incondicional de sus instalaciones. Al doctor Hernán Peña Villamil director del Centro Multisectorial SENA de Mosquera por su permanente colaboración y asesoría y al profesor Guillermo Corredor por su importante y constante asesoría en el análisis estadístico.
Literatura citada
Azcón-Bieto, J. y M. Talón. 1993. Fisiología y bioquímica de plantas. McGraw-Hill Interamericana, Madrid 581 p. [ Links ]
Azcón-Bieto, J. y M. Talón. 2000. Fundamentos de fisiología vegetal. McGraw-Hill Interamericana, Barcelona. 522 p. [ Links ]
Barceló, J., G. Nicolás, B. Sabater y R. Sánchez. 1995. Fisiología vegetal, 2a ed. Ediciones Pirámide S.A., Madrid. 662 p. [ Links ]
Cañet, F.M., M. Vega, M. Gordillo y E. Peña. 2003. Importancia del aseguramiento de la calidad e inocuidad en las producciones orgánicas de frutas y vegetales. V Encuentro de Agricultura Orgánica. ACTAF: Enfocando una agricultura orgánica sostenible frente al desafío de la mega-urbanización en América Latina y el Caribe. En: http://www.rlc.fao.org/prior/segalim/ aup/pdf/organica.pdf. pp. 115-137; consulta: marzo 2007. [ Links ]
Carvajal, C. 1983. Hortalizas en el trópico. Universidad Nacional Agraria de la Selva. Centro de Investigación y Promoción Amazónica, Bogotá. 254 p. [ Links ]
Espinal, T.L. y E.M. Montenegro. 1963. Formaciones vegetales de Colombia. Instituto Geográfico Agustín Codazzi. Departamento Agrológico, Bogotá. 146 p. [ Links ]
Fernández J.A. 1995. Influencia de las condiciones medioambientales en el crecimiento y desarrollo de la coliflor. Departamento de Biología Vegetal, Universidad de Murcia. En: http://www. cibernetia.com/tesis_es/ciencias_de_la_vida/botanica/fisiologia_ vegetal/11; consulta: marzo 2007. [ Links ]
Fu, X., D. Sudhakar, J. Peng, D.E. Richard, P. Christou y N. Haarberd. 2001. Expression of Arabidopsis AGI in transgenic rice represses multiple gibberellin responses. Plant Cell 13, 1791- 1802. [ Links ]
Gómez, C. 1994. Hormonas vegetales. Escuela Técnica Superior de Ingenieros Agrónomos, Universidad Politécnica de Madrid, 386 p. [ Links ]
Ikeda, A., M. Ueguchi-Tanaka, Y. Sonoda, H. Kitano, M. Koshioka, Y. Futsuhara, M. Matsuoka y J. Yamaguchi. 2001. Slender rice, a constitutive gibberellin response mutant, is caused by a null mutation of the SLR1 gene, an ortholog of the heightregulating gene AGI/RAG/RHT/D8. Plant Cell Nagoya 113, 999-1010. [ Links ]
Instituto Geográfico Agustín Codazzi (IGAC).1996. Diccionario geográfico de Colombia. Tomo 3. Tercera edición, Bogotá. 1858 p. [ Links ]
Jaramillo, J. 1982. Programa Nacional de Hortalizas. Informe Anual de Progreso, Barcelona. 286 p. [ Links ]
Krarup, C. e I. Moreira. 1998. Hortalizas de estación fría. Biología y diversidad cultural. Universidad Católica de Chile, Facultad de Agronomía e Ingeniería Forestal, Santiago. En: http://www. puc.cl/sw_educ/hort0498; consulta: marzo 2007. [ Links ]
Lobo, M., E.L. Girard y V.E. Mejía. 1983. Hortalizas. Coliflor. Manual de asistencia técnica. Instituto Colombiano Agropecuario (ICA), Bogotá. pp. 189-195. [ Links ]
Luckwill, L 1994. Reguladores de crecimiento en la producción vegetal. Editorial Monserrat, Barcelona. 408 p. [ Links ]
Maroto, J. 1992. Horticultura herbácea especial. 3ª edición. Ediciones Mundi-Prensa, Madrid. 568 p. [ Links ]
Murayama, S. 1987. Horticultura. Instituto Campineiro de Ensino Agrícola. ICEA, Sao Paulo. 115 p. [ Links ]
O'Neill, D.P. y J.J. Ross. 2002. Auxin regulation of the gibberellin pathway in pea. Plant Physiol. 130, 1974-1982. [ Links ]
Pantastico, E.R.B. 1984. Fisiología de la posrecolección, manejo y utilización de frutas y hortalizas tropicales y subtropicales. Editorial Continental, México. 250 p. [ Links ]
Pérez, E. 1978. Plantas útiles de Colombia. Litografía ARCO, Bogotá. 831 p. [ Links ]
Reigosa, M.J., N. Pedrol y A. Sánchez. 2004. La ecofisiología vegetal. Una ciencia de síntesis. Paraninfo S. A., Madrid. 1193 p. [ Links ]
Salisbury, F.B. y C.W. Ross. 2000. Fisiología de las plantas. Desarrollo de las plantas y fisiología ambiental. Paraninfo S. A., Madrid. pp. 529-988. [ Links ]
Singh, D.P., A.M. Jermakow y S.M. Swain. 2002. Gibberellins are required for seed development and pollen tube growth in Arabidopsis. Plant Cell 14, 3133-3147. [ Links ]
Soriano, J. 1995. Determinación de la 'Unidad fuente demanda' en el llenado del fruto de guanábana (Annona muricata L.). Facultad de Agronomía, Universidad Nacional de Colombia. Bogotá. 103 p. [ Links ]
Sponsel, V.M. 1996. Capítulo B2. The biosynthesis and metabolism of gibberellins in higher plants. pp. 66-97. En: Davies, P.J. (ed.). Plant hormones. Physiology, biochemistry and molecular biology. Kluwer Academic Publishers, Dordrecht. 833 p. [ Links ]
Sponsel, V.M. y P. Hedden. 2004. Capítulo B2: Gibberellin biosynthesis and inactivation. pp. 63-94. En: Davies, P.J. (ed.). Plant hormones. Biosynthesis, signal transduction, action! Kluwer Academic Publishers, Dordrecht. 750 p. [ Links ]
Srivastava, L.M. 2002. Plant growth and development. Hormones and environment. Academic Press, Amsterdam. 772 p. [ Links ]
Taiz, L. y E. Zeiger. 1998. Plant physiology, 2nd edition. Sinauer Associates Inc. Publishers, Sunderland. 792 p. [ Links ]
Weaver, R.J. 1996. Reguladores de crecimiento de las plantas en la agricultura. Editorial Trillas, México. 622 p. [ Links ]

